Доставка жирных кислот к месту их окисления – к митохондриям –
происходит сложным путем:
при участии альбумина осуществляется транспорт жирной кислоты в клетку;
при участии специальных белков (fatty acid binding proteins, FABP) – транспорт в пределах цитозоля до митохондрий;
при участии карнитина – транспорт жирной кислоты из цитозоля в митохондрии.
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ презентация
Содержание
- 1. ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
- 2. Активация жирных кислот. Свободная жирная кислота независимо
- 3. Под действием фермента карнитинацилтрансферазы I (КАТ 1)
- 4. Интенсивность поступления ВЖК в матрикс митохондрий
- 5. Попав в матрикс митохондрий, ацильный остаток в
- 6. Первая стадия дегидрирования. Ацил-КоА в
- 7. Стадия гидратации. Ненасыщенный ацил-КоА (еноил-КоА) при
- 8. Вторая стадия дегидрирования. Образовавшийся β-оксиацил-КоА
- 9. Тиолазная реакция. Тиолазная реакция представляет
- 11. Таким образом, попав в матрикс митохондрий, ацильный
- 12. Выход энергии при β-окислении ВЖК За один
- 13. Общая формула для подсчета выхода АТФ при
- 14. Окисление ненасыщенных ВЖК идет по пути β-окисления
- 16. Жирные кислоты с нечетным числом углеродных атомов
- 18. Нарушения процесса β-окисления. Встречаются патологии, связанные
Слайд 1ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
Окисление ВЖК протекает в митохондриальном матриксе только в аэробных
условиях, так как тесно связано с функционированием цитратного цикла и цепи переноса электронов.

Слайд 2Активация жирных кислот. Свободная жирная кислота независимо от длины углеводородной цепи
является метаболически инертной и не может подвергаться никаким биохимическим превращениям, в том числе окислению, пока не будет активирована.
Активация жирной кислоты протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима A (HS-KoA) и ионов Mg2+.
Реакция катализируется ферментом ацил-КоА-синтетазой:
Активация жирной кислоты протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима A (HS-KoA) и ионов Mg2+.
Реакция катализируется ферментом ацил-КоА-синтетазой:
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.

Слайд 3Под действием фермента карнитинацилтрансферазы I (КАТ 1) ацильный остаток с ацил-КоА
переносится на карнитин с образованием ацилкарнитина. Специфическая транслоказа проводит это вещество через внутреннюю мембрану митохондрий.
На внутренней поверхности внутренней мембраны митохондрий карнитинацилтрансфераза II (КАТ П) расщепляет ацилкарнитин с помощью митохондриального HS-КоА.
Ацил-КоА, освобождающийся в матрикс, участвует в реакциях β-окисления, а свободный карнитин той же транслоказой возвращается на наружную мембрану.
 ацильный остаток с ацил-КоА переносится на карнитин с")
Слайд 4
Интенсивность поступления ВЖК в матрикс митохондрий зависит от соотно-
шения количества малонил-КоА/ацил-КоА.
Чем выше в клетке концентрация малонил-КоА, тем ниже скорость переноса жирных кислот в матрикс митохондрий, так как малонил-КоА — аллостерический ингибитор КАТ1, а ацил-КоА — его активатор

Слайд 5Попав в матрикс митохондрий, ацильный остаток в циклическом процессе с помощью
совокупности ферментов окисляется по β-углеродному атому.
Каждый цикл включает четыре последовательные реакции, в результате которых жирная кислота укорачивается на два углеродных атома, которые отщепляются в виде ацетил-КоА.
Ацетил-КоА может вступать в цитратный цикл и окисляться до СО2 и Н2О, а укороченный ацильный остаток будет вовлекаться в следующий цикл β-окисления.
Каждый цикл включает четыре последовательные реакции, в результате которых жирная кислота укорачивается на два углеродных атома, которые отщепляются в виде ацетил-КоА.
Ацетил-КоА может вступать в цитратный цикл и окисляться до СО2 и Н2О, а укороченный ацильный остаток будет вовлекаться в следующий цикл β-окисления.

Слайд 6Первая стадия дегидрирования.
Ацил-КоА в митохондриях прежде всего подвергается ферментативному дегидрированию,
при этом ацил-КоА теряет 2 атома водорода в α- и β-положениях, превращаясь в КоА-эфир ненасыщенной кислоты.
Таким образом, первой реакцией в каждом цикле распада ацил-КоА является его окисление ФАД-содержащей ацил-КоА-дегидрогеназой, приводящее к образованию еноил-КоА с двойной связью между С-2 и С-3:
Таким образом, первой реакцией в каждом цикле распада ацил-КоА является его окисление ФАД-содержащей ацил-КоА-дегидрогеназой, приводящее к образованию еноил-КоА с двойной связью между С-2 и С-3:

Слайд 7Стадия гидратации.
Ненасыщенный ацил-КоА (еноил-КоА) при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды.
В
результате образуется β-оксиацил-КоА (или 3-гидроксиацил-КоА):
 при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды.В результате образуется β-оксиацил-КоА (или 3-гидроксиацил-КоА):")
Слайд 8Вторая стадия дегидрирования.
Образовавшийся β-оксиацил-КоА (3-гидроксиацил-КоА) затем дегидрируется.
Эту реакцию катализируют
НАД+-зависимые дегидрогеназы:
 затем дегидрируется. Эту реакцию катализируют НАД+-зависимые дегидрогеназы:")
Слайд 9Тиолазная реакция.
Тиолазная реакция представляет собой расщепление 3-оксоацил-КоА
с помощью тиоловой
группы второй молекулы КоА.
В результате образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетил-КоА. Данная реакция катализируется ацетил-КоА-ацилтрансферазой (β-кетотиолазой):
В результате образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетил-КоА. Данная реакция катализируется ацетил-КоА-ацилтрансферазой (β-кетотиолазой):

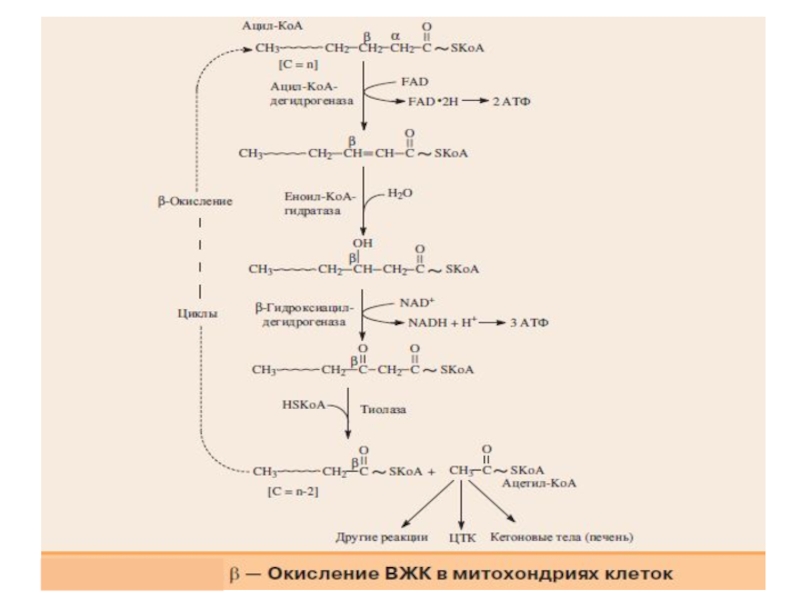
Слайд 11Таким образом, попав в матрикс митохондрий, ацильный остаток в циклическом процессе
с помощью совокупности ферментов окисляется по β-углеродному атому.
Каждый цикл включает четыре последовательные реакции, в результате которых жирная кислота укорачивается на два углеродных атома, которые отщепляются в виде ацетил-КоА. Ацетил-КоА может вступать в цитратный цикл и окисляться до СО2 и Н2О, а укороченный ацильный остаток будет вовлекаться в следующий цикл β-окисления.
В результате β-окисления ВЖК полностью расщепляются до ацетил-КоА, и
суммарное уравнение окисления, например пальмитиновой кислоты (С16) име-
ет следующий вид:
СН3(СН2)14-СО~SКоА + 7FAD + 7NAD+ +7H2O + 7HSKoA→ 8CH3-
CO~SКоА + 7FADH2 + 7(NADH + H+)
Каждый цикл включает четыре последовательные реакции, в результате которых жирная кислота укорачивается на два углеродных атома, которые отщепляются в виде ацетил-КоА. Ацетил-КоА может вступать в цитратный цикл и окисляться до СО2 и Н2О, а укороченный ацильный остаток будет вовлекаться в следующий цикл β-окисления.
В результате β-окисления ВЖК полностью расщепляются до ацетил-КоА, и
суммарное уравнение окисления, например пальмитиновой кислоты (С16) име-
ет следующий вид:
СН3(СН2)14-СО~SКоА + 7FAD + 7NAD+ +7H2O + 7HSKoA→ 8CH3-
CO~SКоА + 7FADH2 + 7(NADH + H+)

Слайд 12Выход энергии при β-окислении ВЖК
За один цикл β-окисления образуется 1 моль
ацетил-КоА, окисление кото-
рого в цитратном цикле обеспечивает синтез 12 моль АТФ. Кроме того, в этом
процессе восстанавливаются 1 моль FADH2 и 1 моль NADH, окисление которых в дыхательной цепи дает 2 и 3 моль АТФ соответственно.
При окислении пальмитиновой кислоты (С16) проходит 7 циклов β-окисления и образуется 8 моль ацетил-КоА, 7 моль FADH2 и 7 моль NADH+Н+. Следовательно, выход АТФ составляет 35 АТФ при β-окислении и 96 АТФ в результате цитратного цикла, что в сумме дает 131 моль АТФ. Поскольку активация ВЖК требует затраты 1 АТФ, то из 131 следует вычесть 1 моль АТФ.
рого в цитратном цикле обеспечивает синтез 12 моль АТФ. Кроме того, в этом
процессе восстанавливаются 1 моль FADH2 и 1 моль NADH, окисление которых в дыхательной цепи дает 2 и 3 моль АТФ соответственно.
При окислении пальмитиновой кислоты (С16) проходит 7 циклов β-окисления и образуется 8 моль ацетил-КоА, 7 моль FADH2 и 7 моль NADH+Н+. Следовательно, выход АТФ составляет 35 АТФ при β-окислении и 96 АТФ в результате цитратного цикла, что в сумме дает 131 моль АТФ. Поскольку активация ВЖК требует затраты 1 АТФ, то из 131 следует вычесть 1 моль АТФ.

Слайд 13Общая формула для подсчета выхода АТФ при полном окислении насыщенных ВЖК
:
[(n/2 — 1)x 5 + n/2 × 12] — 1,
где n — число С атомов в молекуле ВЖК;
n/2 — 1 — число циклов β-окисления;
5 — выход АТФ в одном цикле β-окисления;
n/2 — число ацетильных остатков;
12 — выход АТФ при полном окислении ацетил-КоА в цитратном цикле до СО2 и Н2О.
[(n/2 — 1)x 5 + n/2 × 12] — 1,
где n — число С атомов в молекуле ВЖК;
n/2 — 1 — число циклов β-окисления;
5 — выход АТФ в одном цикле β-окисления;
n/2 — число ацетильных остатков;
12 — выход АТФ при полном окислении ацетил-КоА в цитратном цикле до СО2 и Н2О.
x 5")
Слайд 14Окисление ненасыщенных ВЖК идет по пути β-окисления до получения жирной кислоты
с –НС=СН-связью в положении С3–С4. Дополнительные ферменты: еноил-КоА-изомераза и трансфераза перемещают двойную связь в положение 2–3 и цис-изомер превращают в транс-изомер. Далее β-окисление продолжается с участием ферментов, описанных ранее.
При расчете выхода АТФ за счет окисления ненасыщенных ВЖК можно пользоваться формулой для расчета выхода энергии при окислении насыщенных ВЖК, вычитая 2АТФ на каждую двойную связь.
При расчете выхода АТФ за счет окисления ненасыщенных ВЖК можно пользоваться формулой для расчета выхода энергии при окислении насыщенных ВЖК, вычитая 2АТФ на каждую двойную связь.


Слайд 16Жирные кислоты с нечетным числом углеродных атомов окисляются таким же образом,
как и жирные кислоты с четным числом углеродных атомов,однако, что на последнем этапе расщепления (β-окисления) образуется одна молекула пропионил-КоА и одна молекула ацетил-КоА, а не 2 молекулы ацетил-КоА.
Активированный трехуглеродный фрагмент – пропионил-КоА – включается в цикл трикарбоновых кислот после превращения в сукцинил-КоА.
Активированный трехуглеродный фрагмент – пропионил-КоА – включается в цикл трикарбоновых кислот после превращения в сукцинил-КоА.


Слайд 18Нарушения процесса β-окисления.
Встречаются патологии, связанные со снижением транспорта ВЖК в
матрикс митохондрий. Они могут быть вызваны:
– дефицитом карнитина в результате снижения его синтеза, потерями этого вещества при гемодиализе или за счет экскреции с кетоновыми телами;
– низкой активностью КАТ 1 фермента, связанной с дефектом в структуре гена этого фермента или его ингибированием некоторыми лекарственными препаратами, например сульфонилмочевиной, которая используется для лечения сахарного диабета.
Среди ферментов β-окисления ацил-КоА-дегидрогеназа представлена не-
сколькими видами, специфичными к длине углеводородного радикала жир
ной кислоты.
Достаточно часто встречается наследственная болезнь, вызванная дефектами в структуре гена ацил-КоА-дегидрогеназы, окисляющей жирные кислоты со средним числом углеродных атомов –С4 — С12. Установлено, что смерть каждого десятого новорожденного происходит в результате недостаточности этого фермента. В жирах молока содержится много среднецепочечных жирных кислот, которые не могут окисляться у таких детей. Единственным источником энергии для таких больных становятся углеводы, а при сравнительно продолжительных интервалах между кормлениями развивается тяжелая гипогликемия, сопровождающаяся потерей сознания, а иногда и гибелью ребенка.
– дефицитом карнитина в результате снижения его синтеза, потерями этого вещества при гемодиализе или за счет экскреции с кетоновыми телами;
– низкой активностью КАТ 1 фермента, связанной с дефектом в структуре гена этого фермента или его ингибированием некоторыми лекарственными препаратами, например сульфонилмочевиной, которая используется для лечения сахарного диабета.
Среди ферментов β-окисления ацил-КоА-дегидрогеназа представлена не-
сколькими видами, специфичными к длине углеводородного радикала жир
ной кислоты.
Достаточно часто встречается наследственная болезнь, вызванная дефектами в структуре гена ацил-КоА-дегидрогеназы, окисляющей жирные кислоты со средним числом углеродных атомов –С4 — С12. Установлено, что смерть каждого десятого новорожденного происходит в результате недостаточности этого фермента. В жирах молока содержится много среднецепочечных жирных кислот, которые не могут окисляться у таких детей. Единственным источником энергии для таких больных становятся углеводы, а при сравнительно продолжительных интервалах между кормлениями развивается тяжелая гипогликемия, сопровождающаяся потерей сознания, а иногда и гибелью ребенка.






