- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Кроветворение (гемопоэз) презентация
Содержание
- 1. Кроветворение (гемопоэз)
- 2. Кроветворение (гемопоэз) – процесс образования крови.
- 3. Согласно унитарной теории кроветворения,
- 4. Эмбриональный гемопоэз делится на три периода в
- 5. Мегалобластический период начинается со 2-3 недели внутриутробной
- 6. Большая часть бластов делится и превращается в
- 7. Одновременно с эритропоэзом в желточном мешке экстраваскулярно
- 8. В печени кроветворение начинается на 5-6 неделях
- 9. Начиная с 5-го месяца красный костный мозг
- 10. Постэмбриональный гемопоэз – процесс образования форменных элементов
- 11. Современные представления о кроветворении основаны на признании
- 13. КЛАСС I. - СТВОЛОВАЯ ГЕМОПОЭТИЧЕСКАЯ КЛЕТКА (СКК)
- 14. способность к самоподдержанию: СКК способны поддерживать постоянство
- 15. устойчивость к действию повреждающих факторов, вероятно вследствие
- 16. морфологически СКК не идентифицируются: то есть их
- 17. основное место локализации СКК – красный костный
- 18. II. КЛАСС - МУЛЬТИПОТЕНТНЫЕ КОММИТИРОВАННЫЕ, частично детерминированные
- 19. Мультипотентные Коммитированные клетки, как и клетки следующего
- 20. КЛАСС III. УНИПОТЕНТНЫЕ (КОММИТИРОВАННЫЕ) РОДОНАЧАЛЬНЫЕ КЛЕТКИ
- 21. IY. КЛЕТКИ-ПРЕДШЕСТВЕННИКИ (БЛАСТЫ, PRECURSORS) представляют отдельные
- 22. Y.Созревающие клетки Подвергаются структурной и функциональной дифференцировке,
- 23. ДИФФЕРОН - совокупность всех клеток, составляющих ту
- 24. ЭРИТРОЦИТОПОЭЗ ДИФФЕРОН ЭРИТРОЦИТАРНОГО РЯДА СКК → КОЕ-ГЭММ
- 25. Начало эритроидного ряда – взрывообразующая единица эритропоэза
- 26. Из проэритробласта последовательно образуются: базофильный эритробласт (накопление
- 27. При дифференцировке предшественников эритроцитов в зрелые эритроциты
- 28. Гранулоцитопоэз СКК → КОЕ-ГЭММ → КОЕ-ГнМ →
- 29. Гранулоциты при развитии проходят следующие стадии: миелобласт
- 30. По мере созревания гранулоцитов в зрелые клетки
- 31. Тромбоцитопоэз – процесс образования и созревания тромбоцитов
- 32. Мегакариоцит – очень крупная клетка (до 150
- 33. В ходе дифференцировки происходит образование и накопление

Слайд 2Кроветворение (гемопоэз) – процесс образования крови.
Выделяют эмбриональный и постэмбриональный гемопоэз.
Эмбриональный гемопоэз – это процесс образования крови как ткани.
Постэмбриональный гемопоэз – процесс образования форменных элементов крови в ходе физиологической и репаративной регенерации.
 – процесс образования крови. Выделяют эмбриональный и постэмбриональный гемопоэз. Эмбриональный гемопоэз – это")
Слайд 3
Согласно унитарной теории кроветворения, все клетки крови развиваются из одной родоначальной
стволовой кроветворной клетки (СКК).
.")
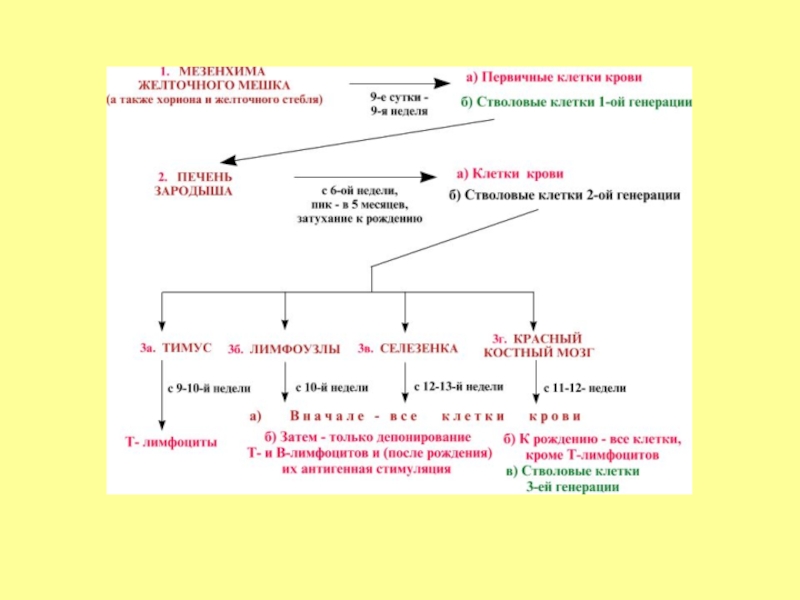
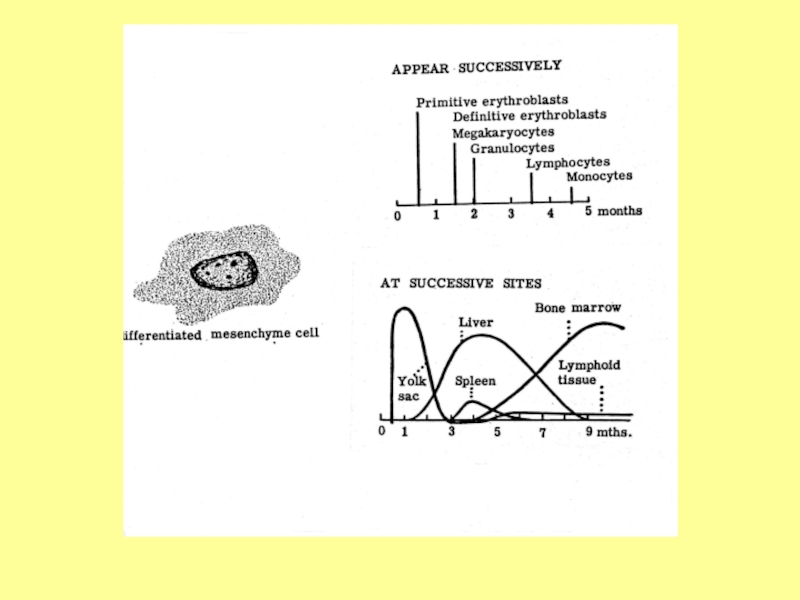
Слайд 4Эмбриональный гемопоэз делится на три периода в зависимости от времени и
места протекания. Это периоды в определенной степени перекрываются:
мегалобластический (внезародышевый) период - 1-2-й месяцы эмбриогенеза;
гепато-тимо-лиенальный период - 2-5-й месяцы эмбриогенеза;
медулло-тимо-лимфатический период – 5-10-й месяцы эмбриогенеза.
мегалобластический (внезародышевый) период - 1-2-й месяцы эмбриогенеза;
гепато-тимо-лиенальный период - 2-5-й месяцы эмбриогенеза;
медулло-тимо-лимфатический период – 5-10-й месяцы эмбриогенеза.

Слайд 5Мегалобластический период начинается со 2-3 недели внутриутробной жизни в мезенхиме
желточного мешка.
В результате интенсивного деления клеток в мезенхиме образуются кровяные островки, клетки которых дифференцируются в двух направлениях:
ангиобласты, лежащие по периферии, превращаются в эндотелий и образуют стенки первичных кровеносных сосудов;
стволовые кроветворные клетки, которые лежат в центре островков, превращаются в первичные клетки крови – бласты.
В результате интенсивного деления клеток в мезенхиме образуются кровяные островки, клетки которых дифференцируются в двух направлениях:
ангиобласты, лежащие по периферии, превращаются в эндотелий и образуют стенки первичных кровеносных сосудов;
стволовые кроветворные клетки, которые лежат в центре островков, превращаются в первичные клетки крови – бласты.

Слайд 6Большая часть бластов делится и превращается в первичные эритробласты больших размеров
– мегалобласты. Мегалобласты активно делятся и начинают синтезировать и накапливать эмбриональные гемоглобины.
Из оксифильных мегалобластов образуются эритроциты больших размеров – мегалоциты. Часть мегалоцитов содержат ядро, часть – является безъядерными. Процесс образования мегалоцитов называется мегалобластическим эритропоэзом. Кроме мегалоцитов в желточном мешке образуется некоторое количество безъядерных эритроцитов обычного размера - нормобластический эритропоэз. Образование эритроцитов в желточном мешке идёт внутри кровеносных сосудов – интраваскулярно.
Из оксифильных мегалобластов образуются эритроциты больших размеров – мегалоциты. Часть мегалоцитов содержат ядро, часть – является безъядерными. Процесс образования мегалоцитов называется мегалобластическим эритропоэзом. Кроме мегалоцитов в желточном мешке образуется некоторое количество безъядерных эритроцитов обычного размера - нормобластический эритропоэз. Образование эритроцитов в желточном мешке идёт внутри кровеносных сосудов – интраваскулярно.

Слайд 7Одновременно с эритропоэзом в желточном мешке экстраваскулярно – вне просвета сосудов
- идёт гранулоцитопоэз – образуются нейтрофильные и эозинофильные гранулоциты.
После образования кровеносных сосудов в теле зародыша и соединения их с сосудами желточного мешка эти клетки попадают в другие органы, участвующие в эмбриональном гемопоэзе.
В дальнейшем желточный мешок постепенно редуцируется, и к 12-й неделе эмбриогенеза кроветворение в нём полностью прекращается
После образования кровеносных сосудов в теле зародыша и соединения их с сосудами желточного мешка эти клетки попадают в другие органы, участвующие в эмбриональном гемопоэзе.
В дальнейшем желточный мешок постепенно редуцируется, и к 12-й неделе эмбриогенеза кроветворение в нём полностью прекращается

Слайд 8В печени кроветворение начинается на 5-6 неделях развития. Здесь образуются эритроциты,
гранулоциты и тромбоциты.
К концу 5-го месяца интенсивность гемопоэза в печени уменьшается, но в небольшой степени продолжается ещё несколько недель после рождения.
Гемопоэз в селезёнке наиболее выражен с 4-го по 8-й месяцы внутриутробного развития.
К концу 5-го месяца интенсивность гемопоэза в печени уменьшается, но в небольшой степени продолжается ещё несколько недель после рождения.
Гемопоэз в селезёнке наиболее выражен с 4-го по 8-й месяцы внутриутробного развития.

Слайд 9Начиная с 5-го месяца красный костный мозг постепенно становится универсальным органом
кроветворения, и происходит разделение на миелопоэз (образование всех видов форменных элементов крови за исключением лимфоцитов) и лимфопоэз

Слайд 10Постэмбриональный гемопоэз – процесс образования форменных элементов крови в ходе физиологической
и репаративной регенерации после рождения. Обновление различных клеточных популяций крови необходимо, поскольку абсолютное большинство форменных элементов крови имеет короткий жизненный цикл (скорость распада эритроцитов, например, составляет 10 млн в секунду).
Гемопоэз обеспечивает поддержание постоянного количества форменных элементов в периферической крови.
Постэмбриональный гемопоэз протекает в миелоидной (красный костный мозг) и лимфоидных (тимус, селезенка, лимфоузлы, миндалины, аппендикс, лимфатические фолликулы) тканях.
Гемопоэз обеспечивает поддержание постоянного количества форменных элементов в периферической крови.
Постэмбриональный гемопоэз протекает в миелоидной (красный костный мозг) и лимфоидных (тимус, селезенка, лимфоузлы, миндалины, аппендикс, лимфатические фолликулы) тканях.

Слайд 11Современные представления о кроветворении основаны на признании унитарной теории кроветворения. Согласно
этой теории, развитие всех клеток крови начинается со стволовой клетки крови (СКК), дифференцировка которой в различные форменные элементы определяется микроокружением и действием специфических веществ – гемопоэтинов.
Во взрослом организме человека СКК в норме локализованы в костном мозге (0,05% от всех клеток костного мозга), однако в низких концентрациях они присутствуют также в периферической крови (0,0001% от всех лимфоцитов). Богатым источником СКК является пуповинная кровь и плацента.
СК дают начало прогениторным клеткам и клеткам-предшественникам, которые делятся и дифференцируются в зрелые клетки определенного типа ткани. Такие клетки называют еще коммитированными.
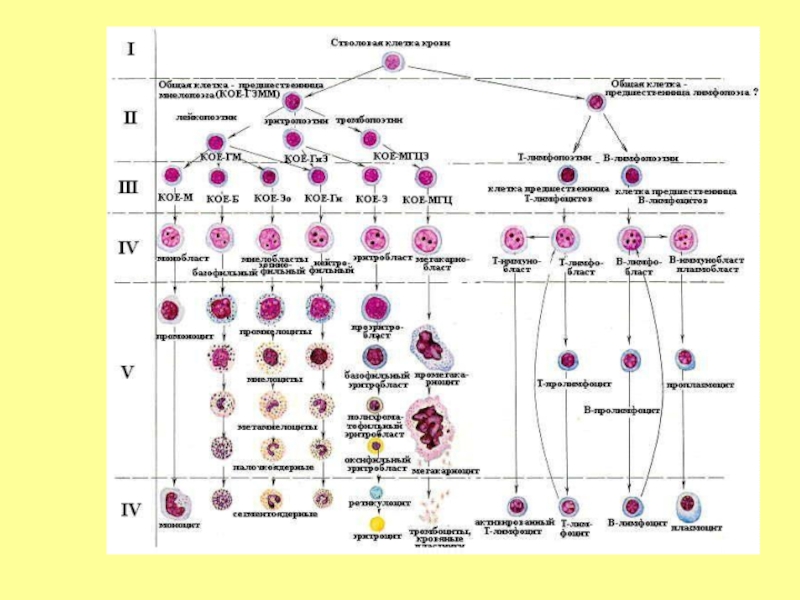
Клетки предшественники образуют дифференцированные клетки через ряд поколений промежуточных клеток, становящихся все более зрелыми. Таким образом, гемопоэтические клетки подразделяются на 6 классов, в зависимости от уровня дифференцировки.
Во взрослом организме человека СКК в норме локализованы в костном мозге (0,05% от всех клеток костного мозга), однако в низких концентрациях они присутствуют также в периферической крови (0,0001% от всех лимфоцитов). Богатым источником СКК является пуповинная кровь и плацента.
СК дают начало прогениторным клеткам и клеткам-предшественникам, которые делятся и дифференцируются в зрелые клетки определенного типа ткани. Такие клетки называют еще коммитированными.
Клетки предшественники образуют дифференцированные клетки через ряд поколений промежуточных клеток, становящихся все более зрелыми. Таким образом, гемопоэтические клетки подразделяются на 6 классов, в зависимости от уровня дифференцировки.

Слайд 13КЛАСС I. - СТВОЛОВАЯ ГЕМОПОЭТИЧЕСКАЯ КЛЕТКА (СКК)
СВОЙСТВА СКК:
плюрипотентность: СКК способна к
дифференцировке в различных направлениях и даёт начало любому виду форменных элементов крови (эритроцитам, лейкоцитам, кровяным пластинкам), поэтому СКК называют родоначальными клетками.
 СВОЙСТВА СКК: плюрипотентность: СКК способна к дифференцировке в")
Слайд 14способность к самоподдержанию: СКК способны поддерживать постоянство численности своей популяции за
счёт того, что после деления стволовой клетки одна из дочерних клеток остается стволовой, сохраняя все свойства родительской клетки; вторая дочерняя клетка дифференцируется в полустволовую (коммитированную) стволовую клетку. Такой митоз называется асимметричным
способность к делению (пролиферации). СКК – долгоживущая клетка; срок её жизни - жизнь индивидуального организма.
способность к делению (пролиферации). СКК – долгоживущая клетка; срок её жизни - жизнь индивидуального организма.

Слайд 15устойчивость к действию повреждающих факторов, вероятно вследствие того, что СКК делятся
редко; большую часть своей жизни они пребывают в состоянии покоя; при необходимости могут вновь вступать в клеточный цикл (например, при значительных кровопотерях и при воздействии факторов роста); кроме того СКК защищены своим местоположением.

Слайд 16морфологически СКК не идентифицируются: то есть их нельзя различить обычными методами
под световым или электронным микроскопом, СКК выглядит как любой малый лимфоцит, но они имеют свой фенотип (антигенный профиль): для них характерно присутствие на поверхности маркеров CD34+,CD59+, Thy1/CD90+, CD38lo/-, C-kit/cd117+, и отсутствие ряда маркеров, свойственных зрелым клеткам крови (Lin-негативность);
благодаря определенному фенотипу СКК можно выявить методами иммуноцитохимии (с помощью меченых моноклональных антител).
благодаря определенному фенотипу СКК можно выявить методами иммуноцитохимии (с помощью меченых моноклональных антител).

Слайд 17основное место локализации СКК – красный костный мозг, хотя численность СКК
невелика (1 СКК на 2000 клеток красного костного мозга; или 1 СКК на 1 000 000 лейкоцитов периферической крови).

Слайд 18II. КЛАСС - МУЛЬТИПОТЕНТНЫЕ КОММИТИРОВАННЫЕ, частично детерминированные (ПОЛУСТВОЛОВЫЕ) КЛЕТКИ
Мультипотентные коммитированные клетки
дают начало форменным элементам крови нескольких, но не всех, видов.
Этот класс представлен 2 типами клеток:
родоначальной клеткой миелопоза – КОЕ-ГЭММ: эта клетка даёт начало гранулоцитам, эритроцитам, моноцитам и мегакариоцитам.
родоначальной клеткой лимфопоэза: эта клетка даёт начало В- и Т-лимфоцитам, натуральным киллерам и некоторым дендритным клеткам.
Клетки этого класса способны к ограниченному самоподдержанию.
Митотическая активность клеток этого класса по-прежнему низкая.
Морфологически не идентифицируются (малые лимфоциты на вид)
Этот класс представлен 2 типами клеток:
родоначальной клеткой миелопоза – КОЕ-ГЭММ: эта клетка даёт начало гранулоцитам, эритроцитам, моноцитам и мегакариоцитам.
родоначальной клеткой лимфопоэза: эта клетка даёт начало В- и Т-лимфоцитам, натуральным киллерам и некоторым дендритным клеткам.
Клетки этого класса способны к ограниченному самоподдержанию.
Митотическая активность клеток этого класса по-прежнему низкая.
Морфологически не идентифицируются (малые лимфоциты на вид)
 КЛЕТКИМультипотентные коммитированные клетки дают начало форменным элементам")
Слайд 19Мультипотентные Коммитированные клетки, как и клетки следующего класса – также называют
КОЛОНИЕОБРАЗУЮЩИМИ
ЕДИНИЦАМИ (КОЕ), поскольку в экспериментах на летально облученных мышах они способны образовывать колонии кроветворных клетках в их органах (селезенке).
Каждая колония возникает как результат деления одной клетки, поэтому анализируя клеточный состав колонии, можно сделать вывод о потентности клетки, давшей начало этой колонии
КОЕ-ГЭММ – значит, что эта клетка даёт селезеночную колонию, состоящую из гранулоцитов (Г), эритроцитов (Э), моноцитов (М) и мегакариоцитов (М).
Каждая колония возникает как результат деления одной клетки, поэтому анализируя клеточный состав колонии, можно сделать вывод о потентности клетки, давшей начало этой колонии
КОЕ-ГЭММ – значит, что эта клетка даёт селезеночную колонию, состоящую из гранулоцитов (Г), эритроцитов (Э), моноцитов (М) и мегакариоцитов (М).
, поскольку в")
Слайд 20КЛАСС III. УНИПОТЕНТНЫЕ (КОММИТИРОВАННЫЕ) РОДОНАЧАЛЬНЫЕ КЛЕТКИ (ПРОГЕНИТОРНЫЕ, PROGENITORS)
унипотентны - детерминированы
в направлении развития только одного вида форменных элементов (за исключением бипотентной КОЕ-ГМ) [детерминация – выбор направления развития];
низкий потенциал самоподдержания
митотическая активность выше, чем у клеток 2-го класса;
морфологически не идентифицируются (малый лимфоцит).
образуют «чистые» колонии (из одного вида форменных элементов).
низкий потенциал самоподдержания
митотическая активность выше, чем у клеток 2-го класса;
морфологически не идентифицируются (малый лимфоцит).
образуют «чистые» колонии (из одного вида форменных элементов).
 РОДОНАЧАЛЬНЫЕ КЛЕТКИ (ПРОГЕНИТОРНЫЕ, PROGENITORS)унипотентны - детерминированы в направлении развития только одного")
Слайд 21IY. КЛЕТКИ-ПРЕДШЕСТВЕННИКИ (БЛАСТЫ, PRECURSORS)
представляют отдельные линии развития форменных элементов;
пролиферативная активность ограничена,
но выше, чем у 3-го класса;
не обладают способностью к самоподдержанию;
морфологически распознаваемые (хотя все клетки этого класса сходны друг с другом, их можно идентифицировать при использовании стандартных методов окраски, не прибегая к выявлению иммуноцитохимических маркеров); имеют вид крупных клеток с крупным светлым овальным ядром, в котором хорошо определяются ядрышки, и базофильную цитоплазму.
не обладают способностью к самоподдержанию;
морфологически распознаваемые (хотя все клетки этого класса сходны друг с другом, их можно идентифицировать при использовании стандартных методов окраски, не прибегая к выявлению иммуноцитохимических маркеров); имеют вид крупных клеток с крупным светлым овальным ядром, в котором хорошо определяются ядрышки, и базофильную цитоплазму.
 представляют отдельные линии развития форменных элементов;пролиферативная активность ограничена, но выше, чем")
Слайд 22Y.Созревающие клетки
Подвергаются структурной и функциональной дифференцировке, в ходе которой утрачивают способность
к делению (за исключением лимфоцитов и моноцитов).
Идентифицируются морфологически
Идентифицируются морфологически

Слайд 23ДИФФЕРОН - совокупность всех клеток, составляющих ту или иную линию дифференцировки
от стволовых (наименее дифференцированных) клеток до терминально (наиболее зрелых) дифференцированных.
Многие ткани содержат несколько различных дифферонов, которые взаимодействуют друг с другом.
Многие ткани содержат несколько различных дифферонов, которые взаимодействуют друг с другом.
")
Слайд 24ЭРИТРОЦИТОПОЭЗ
ДИФФЕРОН ЭРИТРОЦИТАРНОГО РЯДА
СКК → КОЕ-ГЭММ → БОЕ-Э → КОЕ-Э →
→
ПРОЭРИТРОБЛАСТ → БАЗОФИЛЬНЫЙ ЭРИТРОБЛАСТ →
→ ПОЛИХРОМАТОФИЛЬНЫЙ ЭРИТРОБЛАСТ→
→ ОКСИФИЛЬНЫЙ ЭРИТРОБЛАСТ → РЕТИКУЛОЦИТ →
→ ЭРИТРОЦИТ
→ ПОЛИХРОМАТОФИЛЬНЫЙ ЭРИТРОБЛАСТ→
→ ОКСИФИЛЬНЫЙ ЭРИТРОБЛАСТ → РЕТИКУЛОЦИТ →
→ ЭРИТРОЦИТ

Слайд 25Начало эритроидного ряда – взрывообразующая единица эритропоэза – BFU-E.
При активации
и делении BFU-E образуется множество унипотентных КОЕ-Э.
BFU-E реагирует на интерлейкин 3, но в отличие от КОЕ-Э не чувствительна к эритропоэтину, образующемуся в почке.
BFU-E реагирует на интерлейкин 3, но в отличие от КОЕ-Э не чувствительна к эритропоэтину, образующемуся в почке.

Слайд 26Из проэритробласта последовательно образуются:
базофильный эритробласт (накопление рибосом и начало синтеза Hb);
полихроматофильный
эритробласт (накопление Hb);
оксифильный эритробласт (высокое содержание Hb и остатки белоксинтезирущего аппарата, потеря способности к делению и выброс ядра);
ретикулоцит
оксифильный эритробласт (высокое содержание Hb и остатки белоксинтезирущего аппарата, потеря способности к делению и выброс ядра);
ретикулоцит
;полихроматофильный эритробласт (накопление Hb);оксифильный эритробласт")
Слайд 27При дифференцировке предшественников эритроцитов в зрелые эритроциты происходят следующие процессы:
уменьшение размеров
клетки;
выработка и накопление гемоглобина в цитоплазме;
постепенное снижение числа органелл;
изменение окраски цитоплазмы от базофильной (в связи с большим числом полирибосом) до оксифильной (обусловленной накоплением гемоглобина);
снижение, а в дальнейшем утрата способности к делению;
уменьшение размера, конденсация хроматина и выталкивание ядра из клетки.
выработка и накопление гемоглобина в цитоплазме;
постепенное снижение числа органелл;
изменение окраски цитоплазмы от базофильной (в связи с большим числом полирибосом) до оксифильной (обусловленной накоплением гемоглобина);
снижение, а в дальнейшем утрата способности к делению;
уменьшение размера, конденсация хроматина и выталкивание ядра из клетки.

Слайд 28Гранулоцитопоэз
СКК → КОЕ-ГЭММ → КОЕ-ГнМ → КОЕ-Гн → МИЕЛОБЛАСТ → ПРОМИЕЛОЦИТ
→ МИЕЛОЦИТ →
→ МЕТАМИЕЛОЦИТ→ ПАЛОЧКОЯДЕРНЫЙ НЕЙТРОФИЛ → СЕГМЕНТОЯДЕРНЫЙ НЕЙТРОФИЛ
→ МЕТАМИЕЛОЦИТ→ ПАЛОЧКОЯДЕРНЫЙ НЕЙТРОФИЛ → СЕГМЕНТОЯДЕРНЫЙ НЕЙТРОФИЛ

Слайд 29Гранулоциты при развитии проходят следующие стадии:
миелобласт (не имеет гранул)
промиелоцит (первичные, азурофильные
гранулы)
миелоцит (появление специфических гранул, округлое ядро)
метамиелоцит (бобовидное ядро)
палочкоядерный
сегментоядерный
миелоцит (появление специфических гранул, округлое ядро)
метамиелоцит (бобовидное ядро)
палочкоядерный
сегментоядерный
промиелоцит (первичные, азурофильные гранулы)миелоцит (появление специфических гранул,")
Слайд 30По мере созревания гранулоцитов в зрелые клетки происходит:
уменьшение размеров клетки;
изменение формы
их ядер от округлой до сегментированной;
накопление и изменение состава гранул в цитоплазме (постепенное увеличение доли специфических гранул);
утрата способности к делению;
нарастание подвижности клеток и приобретение разнообразных рецепторов плазмолеммы, обеспечивающих выполнение главных функций клеток (фагоцитоз, хемотаксис и др.).
накопление и изменение состава гранул в цитоплазме (постепенное увеличение доли специфических гранул);
утрата способности к делению;
нарастание подвижности клеток и приобретение разнообразных рецепторов плазмолеммы, обеспечивающих выполнение главных функций клеток (фагоцитоз, хемотаксис и др.).

Слайд 31Тромбоцитопоэз – процесс образования и созревания тромбоцитов происходит в миелоидной ткани.
Тромбоциты (кровяные пластинки) образуются в результате частичной фрагментации цитоплазмы мегакароицитов.
Последовательность дифференцировки можно представить следующим рядом клеток:
СКК → КОЕ-ГЭММ → КОЕ-МГЦ → МЕГАКАРИОБЛАСТ → ПРОМЕГАКАРИОЦИТ → МЕГАКАРИОЦИТ → ТРОМБОЦИТЫ (кровяные пластинки).
Последовательность дифференцировки можно представить следующим рядом клеток:
СКК → КОЕ-ГЭММ → КОЕ-МГЦ → МЕГАКАРИОБЛАСТ → ПРОМЕГАКАРИОЦИТ → МЕГАКАРИОЦИТ → ТРОМБОЦИТЫ (кровяные пластинки).
 образуются")
Слайд 32Мегакариоцит – очень крупная клетка (до 150 мкм в диаметре); имеет
крупное, дольчатое полиплоидное ядро (до 64n), слабобазофильную цитоплазму.
; имеет крупное, дольчатое полиплоидное ядро")
Слайд 33В ходе дифференцировки происходит образование и накопление гранул, характерных для тромбоцитов
и содержащих специфические для них белки;
формирование системы мембран (демаркационных каналов), разрезающих цитоплазму мегакариоцита на участки размером 2-4мкм, соответствующие размерам будущих тромбоцитов;
образование филоподий (протромбоцитов) – узких длинных отростков мегакариоцитов, которые через поры эндотелия синусов красного костного мозга проникают в их просвет и распадаются на отдельные кровяные пластинки.
формирование системы мембран (демаркационных каналов), разрезающих цитоплазму мегакариоцита на участки размером 2-4мкм, соответствующие размерам будущих тромбоцитов;
образование филоподий (протромбоцитов) – узких длинных отростков мегакариоцитов, которые через поры эндотелия синусов красного костного мозга проникают в их просвет и распадаются на отдельные кровяные пластинки.