ТАҚЫРЫБЫ:
Тотығу түрлері.Липидтердің пероксидті тотығуы (ЛПТ), антиоксиданттар.
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Тотығу түрлері.Липидтердің пероксидті тотығуы (ЛПТ), антиоксиданттар презентация
Содержание
- 2. Тотығу түрлері. Оксидазды тотығу 2. Липидтердің
- 3. Оксидазды тотығу=БТ Оттек молекуласы 4е- электронмен
- 4. ЛИПИДТЕРДІҢ ПЕРОКСИДТІ ТОТЫҒУЫ (ЛПТ) ЖӘНЕ ОНЫҢ РӨЛІ.
- 5. ПРООКСИДАНТТАР -ЛИПИДТЕРДІҢ ПЕРОКСИДТІ ТОТЫҒУЫН (ЛПТ)
- 6. ЛПТ дегеніміз – майлардың, әсіресе мембранадағы фосфолипидтер
- 8. Бос радикал дегеніміз не?
- 9. КӨП ҚАНЫҚПАҒАН МАЙ ҚЫШҚЫЛДАРЫ (КҚМҚ) (
- 10. Животные – наиболее ценные среди них –
- 11. Оттек молекуласы әр түрлі реакциялардан бір-бірден электронды
- 14. +2Н+ Н2О2 + О2
- 15. ОТТЕГlНlҢ АКТИВТl ФОРМАЛАРЫ (ОАФ) O2*-
- 16. НОСl- гипохлорит-анион O3 -
- 17. Супероксидті радикал түзіледі: оттек
- 18. Гидроксиль радикалы түзіледі: сутек асқын
- 20. Transition metal-catalyzed formation of free radicals X-H
- 21. Бос радикалдар қайдан пайда болады?
- 22. ОАФ-ың түзілуіне әкелетін себептер: антиоксиданттық жүйенің жеткіліксіздігі,
- 24. л2-сатысы: Түзілген липид радикалы (L1∙) ары қарай
- 25. Түзілген май қышқылының радикалы қайтадан 2-ші реакцияға
- 26. О•2 О2
- 27. Липидтердің пероксидті тотығуының алғашқы өнімі: ДИЕНДl
- 28. L1Н L•1 L1OО• L•2 L 2H L1OОН
- 30. ─Қалыпты жағдайда организмде Бос радикалды процесс жүреді
- 31. ЛПТ не үшін керек?
- 32. Арахидон қышқылынан простагландиндер (ПГ) ж/е олардың туындыларын
- 33. иммунды жүйенің қызметіне қатысады Бактерияларды
- 34. ФЕРМЕНТТІ СУПЕРОКСИДДИСМУТАЗА (СОД);
- 35. Супероксиддисмутаза катализдейтін реакция: О2- + О2- +
- 36. Н202 Fe2+ OH.
- 38. белоктардың SH тобына әсер етіп , оларың
- 39. БИОМЕМБРАНАДАҒЫ КҚМҚ-ын ыдыратып жасуша н/е органеллаларды
- 40. ОКИСЛИТЕЛЬНЫЙ СТРЕСС И ДЕСТРУКЦИЯ КЛЕТКИ
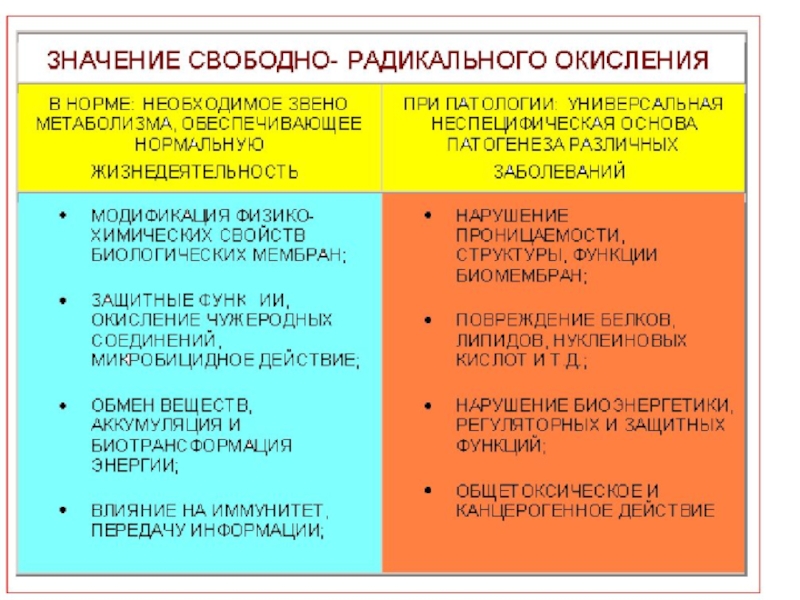
- 42. Бос радикалдар жасушаларға әсер етіп, олардың ДНҚ-сын, белоктарын, майларын зақымдайтын жоғары активті бөлшектер
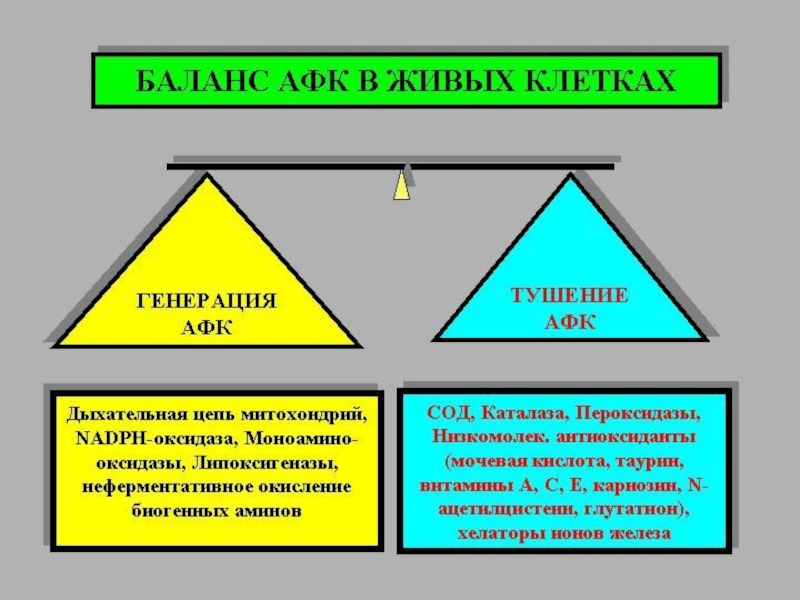
- 43. Оксиданттық стресс және антиоксиданттық қорғаныс
- 46. Антиоксиданттық система .
- 48. Aнтиоксиданттар Антиоксиданттар бос радикалға электронын беріп оларды нейтралдайды.
- 50. Избыток антиоксидантов вызывает прооксидантный
- 51. РЕГУЛЯЦИЯ
- 52. Витамин Е антиоксид ретінде Зақымдалған фосфолипидтер Бос
- 53. C и E как Кo-Aнтиоксиданты Поскольку
- 54. C и E как Кo-Антиоксиданты (2)
- 56. + 2 Н2О
- 57. ВОЗМОЖНО ЛИ ПРЕДОТВРАТИТЬ ИЛИ УДАЛИТЬ СШИВКИ?
- 58. Оксигеназды тотығу эндоплазмалық ретикулум мембрансы (микросома) ж/е
- 59. Монооксигеназдық тотығуда оттектің бір атомы субстратпен байланысады
- 60. Реакция микросомаларда, әсіресе залалсыздандыру бауырда жүреді. Мембранамен
- 61. Оксигеназды тотығудың маңызы Бұл процестің қатысуымен
- 62. Микросомальное окисление - совокупность реакций первой фазы
- 65. Пероксидаздық тотығу Бұл процесс пероксисомаларда жүреді.
- 66. Пероксидаздық тотығу
- 67. Негзгl ж/е косымша әдебиеттер
, антиоксиданттар.")
Слайд 2Тотығу түрлері.
Оксидазды тотығу
2. Липидтердің пероксидті тотығуы (ЛПТ),
Оксигеназды тотығу
Пероксидазды тотығу
90%Оттек
оксидазды тотығуға жұмсалады
10% басқа тотығу түрлеріне пайдаланылады.
10% басқа тотығу түрлеріне пайдаланылады.
, Оксигеназды тотығуПероксидазды тотығу90%Оттек оксидазды тотығуға жұмсалады 10%")
Слайд 3
Оксидазды тотығу=БТ
Оттек молекуласы 4е- электронмен тотықсызданады.
SН2+ ½ О2 S+ Н2О +Е
субстрат
МАҢЫЗЫ:
1. ЭНЕРГИЯ (Е) БӨЛІНЕДІ (Е=40- 45% АТФ +ЖЫЛУ)
2. ЭНДОГЕНДІ СУ ТҮЗІЛЕДІ.

 ЖӘНЕ ОНЫҢ РӨЛІ.")
Слайд 5
ПРООКСИДАНТТАР -ЛИПИДТЕРДІҢ ПЕРОКСИДТІ ТОТЫҒУЫН (ЛПТ) ТУҒЫЗАТЫН ЗАТТАР.
АНТИОКСИДАНТТАР -ЛПТ
ТЕЖЕЙТІН ЗАТТАР.
 ТУҒЫЗАТЫН ЗАТТАР. АНТИОКСИДАНТТАР -ЛПТ ТЕЖЕЙТІН ЗАТТАР.")
Слайд 6 ЛПТ дегеніміз – майлардың, әсіресе мембранадағы фосфолипидтер құрамына кіретін көп қанықпаған
май қышқылдарының (КҚМҚ) бос радикалдық тотығуы.
 бос")
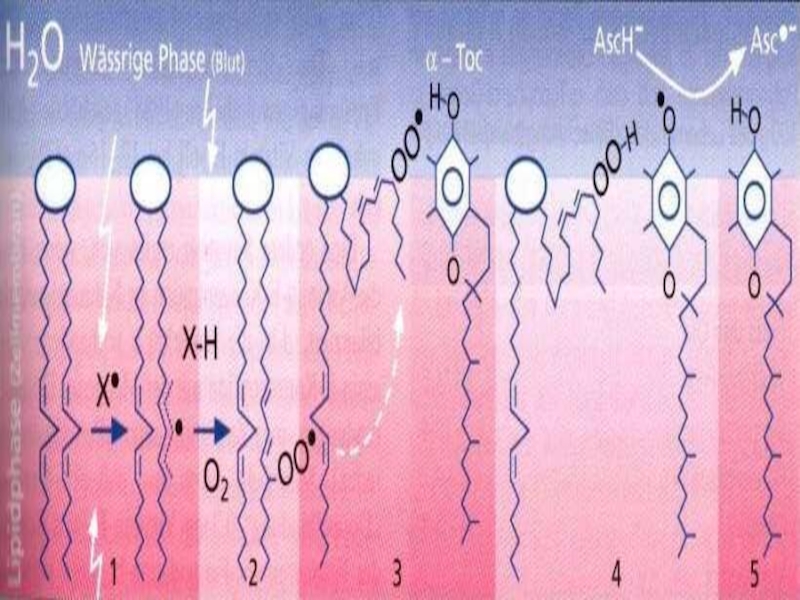
Слайд 8Бос радикал дегеніміз не? Сыртқы валенттік орбитасында жұптаспаған
электроны бар атом немесе атом топтары.
Свободные радикалы стремятся вернуть себе недостающий электрон, отняв его от окружающих молекул
Свободные радикалы стремятся вернуть себе недостающий электрон, отняв его от окружающих молекул

Слайд 9КӨП ҚАНЫҚПАҒАН МАЙ ҚЫШҚЫЛДАРЫ (КҚМҚ) ( ВИТАМИН F, ЭССЕНЦИАЛДЫ МАЙ ҚЫШҚЫЛДАРЫ,
АЛМАСТЫРЫЛМАЙТЫН МАЙ ҚЫШҚЫЛДАРЫ)
С17Н31СООН-ЛИНОЛЬ 18 ¦ 2
С17Н29СООН-ЛИНОЛЕН 18 ¦ 3
С19Н31СООН-АРАХИДОН 20 ¦ 4
С17Н31СООН-ЛИНОЛЬ 18 ¦ 2
С17Н29СООН-ЛИНОЛЕН 18 ¦ 3
С19Н31СООН-АРАХИДОН 20 ¦ 4
 ( ВИТАМИН F, ЭССЕНЦИАЛДЫ МАЙ ҚЫШҚЫЛДАРЫ, АЛМАСТЫРЫЛМАЙТЫН МАЙ ҚЫШҚЫЛДАРЫ) С17Н31СООН-ЛИНОЛЬ")
Слайд 10Животные – наиболее ценные среди них – жиры морской рыбы (содержат
незаменимые ПНЖК)
Растительные – содержат незаменимые ПНЖК (полиненасыщенные жирные кислоты)
Растительные – содержат незаменимые ПНЖК (полиненасыщенные жирные кислоты)
Растительные – содержат")
Слайд 11Оттек молекуласы әр түрлі реакциялардан бір-бірден электронды қосып алып тотықсызданып оттектің
активті формаларына (ОАФ) айналуы мүмкін.
 айналуы")
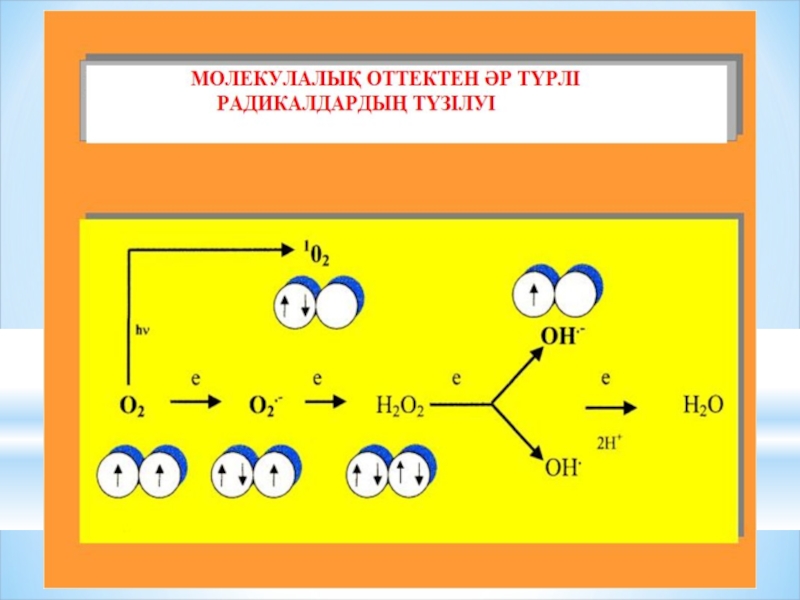

Слайд 14
+2Н+
Н2О2 + О2
О2 + ОН∙ - + ОН∙
(реакция
Хабера-Вайса)
+ О2∙ -
ОН - + ОН∙ - + Fe3+
(реакция Фентон)
+ NO ∙
ONOO-
+ Fe2+
DCFH2
+ О2∙ -ОН - +")
Слайд 15ОТТЕГlНlҢ АКТИВТl ФОРМАЛАРЫ (ОАФ)
O2*- Супероксид-анион
OH-
Гидроксил радикалы
NО- АЗОТ ОКСИДlНlН РАДИКАЛЫ ж/е т.б.
LOO • липопероксирадикал (LOO•),
LO• алкилоксирадикал
NО- АЗОТ ОКСИДlНlН РАДИКАЛЫ ж/е т.б.
LOO • липопероксирадикал (LOO•),
LO• алкилоксирадикал
 O2*- Супероксид-анионOH- Гидроксил радикалыNО-")
Слайд 16 НОСl- гипохлорит-анион
O3 - озон
LOOH (ГПЛ)-липидтер гидропероксиді
'O2
Синглетті оттек
H2O2 Сутек асқын тотығы
H2O2 Сутек асқын тотығы
-липидтер гидропероксиді'O2 Синглетті оттекH2O2 Сутек асқын тотығы")
Слайд 17 Супероксидті радикал түзіледі:
оттек ауыспалы валентті металдармен әрекеттескенде м\лы,
Fe++ + О2 → Fe+++ + О2•
НАДФН-оксидазаның қатысуымен ферментативті жолмен (фагоциттердің ферментативті жүйесі):
НАДФН + 2О2 → НАДФ+ + 2(О2•)

Слайд 18Гидроксиль радикалы түзіледі:
сутек асқын тотығынан (реакция Фентона):
Fe++ +
Н2О2 → Fe+++ + ОН• + ОН––
Н2О2 супероксиданион-радикалымен әрекеттескенде (реакция Хабера-Вайса): Н2О2 + О2• → ОН• + ОН–– + О2
Н2О2 супероксиданион-радикалымен әрекеттескенде (реакция Хабера-Вайса): Н2О2 + О2• → ОН• + ОН–– + О2
: Fe++ + Н2О2 → Fe+++ + ОН•")
Слайд 20Transition metal-catalyzed formation
of free radicals
X-H + Cu+2 → X• + H+
+ Cu+
X-H + Fe+3 → X• + H+ + Fe+2
X-H + Mn+1 → X• + H+ + Mn+
(X-H – e- → X• + H+ )
or, alternatively,
LOOH + Cu+ → LO• + Cu+2 +OH-
Binding transition metals (e.g. chelation) may inhibit oxidation
X-H + Fe+3 → X• + H+ + Fe+2
X-H + Mn+1 → X• + H+ + Mn+
(X-H – e- → X• + H+ )
or, alternatively,
LOOH + Cu+ → LO• + Cu+2 +OH-
Binding transition metals (e.g. chelation) may inhibit oxidation


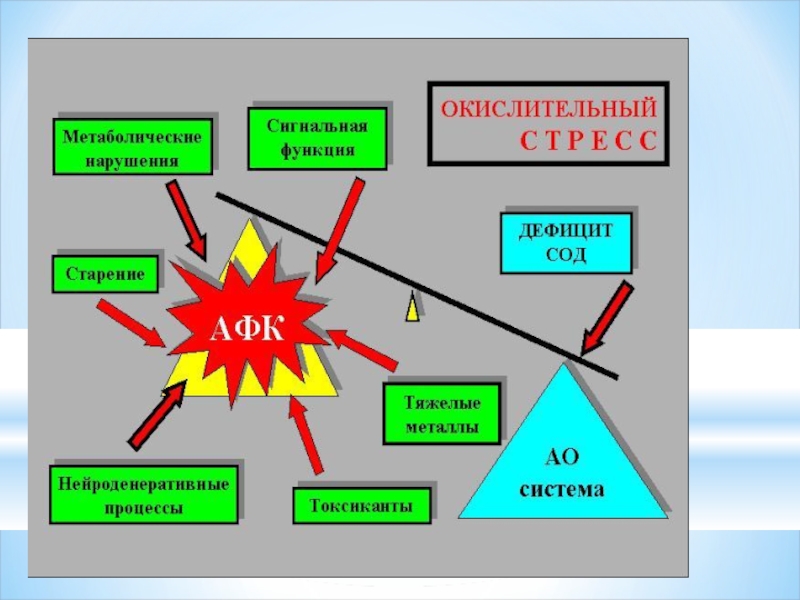
Слайд 22ОАФ-ың түзілуіне әкелетін себептер: антиоксиданттық жүйенің жеткіліксіздігі, радиация сәулесі, шылым шегу,
алкоголизм, қоршаған ортаның ластануы, қартаю, гипоксия, стресс, жоғары қысымды оттегімен емдеу және өспе ауруларын емдеуге арналған химиялық препараттар, т.б.

Слайд 23
ЛПТ реакциялары
ОАФ әсерінен сатылы тізбекті реакция арқылы іске асады.
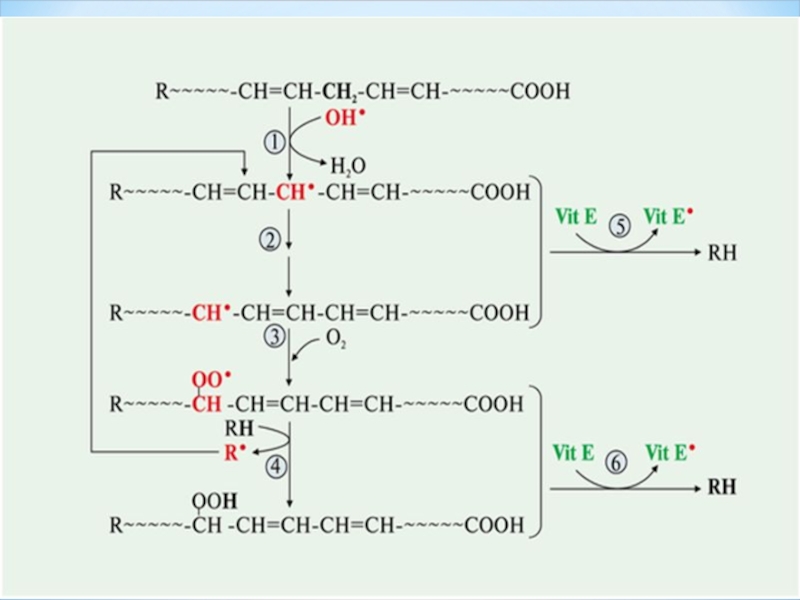
1-сатысы: тізбекті реакцияның басталуы – инициация.
Бос радикалдар (О2∙) мембранадағы фосфолипидтер құрамына кіретін КҚМҚ-дағы (L1Н =КҚМҚ) қос байланысқа жақын орналасқан СН2 –ден (метилен ) Н бөліп алып КҚМҚ қалдығын бос радикалға (L1∙) айналдырады.
ОАФ әсерінен сатылы тізбекті реакция арқылы іске асады.
1-сатысы: тізбекті реакцияның басталуы – инициация.
Бос радикалдар (О2∙) мембранадағы фосфолипидтер құрамына кіретін КҚМҚ-дағы (L1Н =КҚМҚ) қос байланысқа жақын орналасқан СН2 –ден (метилен ) Н бөліп алып КҚМҚ қалдығын бос радикалға (L1∙) айналдырады.

Слайд 24л2-сатысы: Түзілген липид радикалы (L1∙) ары қарай оттек молекуласымен (О2) тез
реакцияласып май қышқылының пероксид радикалын (L1ОО∙) түзеді.
3–сатысы:Түзілген пероксид радикалы (L1ОО∙) жанындағы екінші КҚМҚ– на (L2Н) шабуыл жасап ондағы СН2 –ден Н бөліп алып өзі гидропероксидке, ал КҚМҚ-ын Липид радикалына (L•2) айналдырады.
3–сатысы:Түзілген пероксид радикалы (L1ОО∙) жанындағы екінші КҚМҚ– на (L2Н) шабуыл жасап ондағы СН2 –ден Н бөліп алып өзі гидропероксидке, ал КҚМҚ-ын Липид радикалына (L•2) айналдырады.
 ары қарай оттек молекуласымен (О2) тез реакцияласып май қышқылының пероксид")
Слайд 25Түзілген май қышқылының радикалы қайтадан 2-ші реакцияға одан кейін 3-ші реакцияға
түсіп тағыда радикалдар түзіледі. Бұл сатыда реакция тізбекті жалғаса береді.
3-ші реакцияда түзілген гидропероксид тұрақсыз қосылыс ол қаныққан және қанықпаған альдегидтерге ыдырайды. ОАФ тек процесті бастау үшін қажет , процесс басталса болды ОАФ -на тәуелсіз жалғаса береді.
3-ші реакцияда түзілген гидропероксид тұрақсыз қосылыс ол қаныққан және қанықпаған альдегидтерге ыдырайды. ОАФ тек процесті бастау үшін қажет , процесс басталса болды ОАФ -на тәуелсіз жалғаса береді.


Слайд 27Липидтердің пероксидті тотығуының алғашқы өнімі:
ДИЕНДl КОНЪЮГАТТАР
Аралық өнімдері: ГИДРОПЕРОКСИДТЕР
Соңғы өнімдерінің
бірі:
МАЛОН ДИАЛЬДЕГИДl (МДА)
МАЛОН ДИАЛЬДЕГИДl (МДА)
")

Слайд 30─Қалыпты жағдайда организмде Бос радикалды процесс жүреді ме?
─Үнемі, аз мөлшерде.
Себебі,
ағзадағы антиоксиданттық жүйе оны реттеп отырады.

Слайд 31
ЛПТ не үшін керек?
ЛПТ биологиялық маңызы:
Биомембрананың өткізіштігін арттырады;
Мембрананың липидтік
құрамының жаңалануын қамтамасыз етеді;

Слайд 32Арахидон қышқылынан
простагландиндер (ПГ) ж/е олардың туындыларын түзеді
(простациклиндер, тромбоксандар,
лейкотриендер);
Ксенобиотиктер мен метаболизмнің улы
өнімдерін усыздандырады
кейбір гормондардың синтезіне қатысады. Мысалы, тиреоиды гормондар;
кейбір гормондардың синтезіне қатысады. Мысалы, тиреоиды гормондар;
 ж/е олардың туындыларын түзеді(простациклиндер, тромбоксандар,лейкотриендер);Ксенобиотиктер мен метаболизмнің улы өнімдерін усыздандырады кейбір гормондардың")
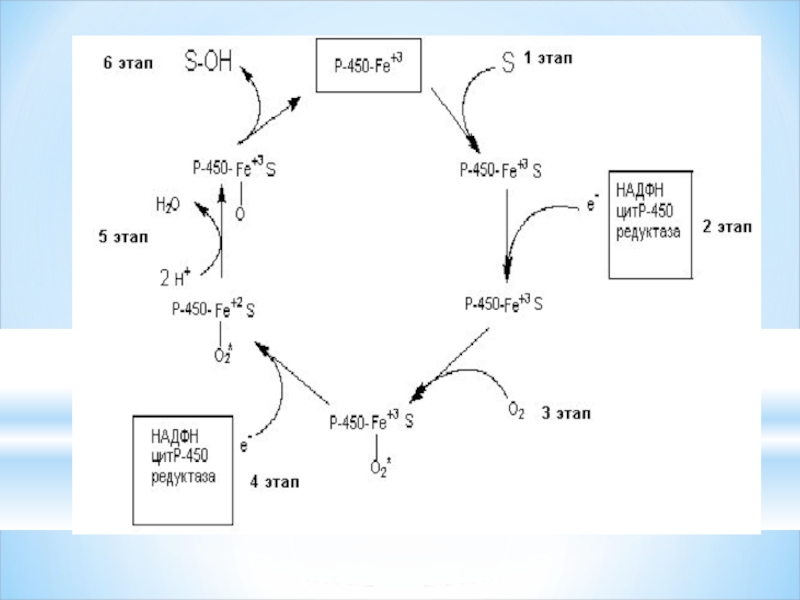
Слайд 33иммунды жүйенің қызметіне қатысады
Бактерияларды жояды. М\ы, жедел қабыну кезінде
лейкоциттер (әсіресе нейтрофилдер) НАДФН-ЦхР450-оксидаза жүйесі және миелопероксидаза- Н2О2-галоген жүйесі арқылы ОАФ түзіп бактерияларды, өспе жасушаларын жояды.
 НАДФН-ЦхР450-оксидаза жүйесі")
Слайд 34 ФЕРМЕНТТІ
СУПЕРОКСИДДИСМУТАЗА (СОД);
КАТАЛАЗА;
ГЛУТАТИОНРЕДУКТАЗА
ГЛУТАТИОНПЕРОКСИДАЗА
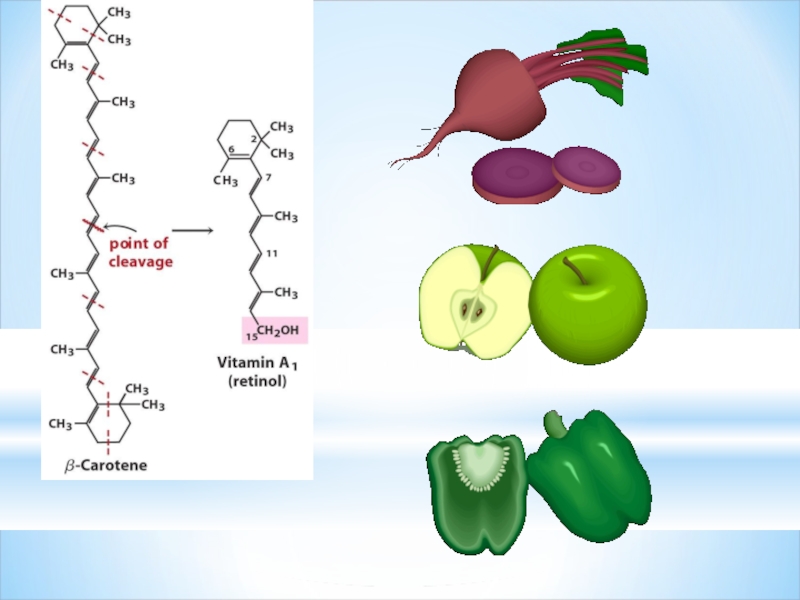
ФЕРМЕНТТІ ЕМЕС
АНТИОКСИДАНТТАР
Майда
еритін витаминдер: А ж\е Е;
С,Р витаминдер
Каротиндер (вит А-ның провитамині)
Карнозин – миоциттерде гидроксил
радикалдарды бейтараптайды
Ферритин – екі валентті темірді
байланыстырады
Церулоплазмин – екі валентті мысты байланыстырады, феррооксидаздық
активтілікті көрсетіп екі валентті темірді тотықтырады
Металлотионеиндер – мысты ж\е
т.б. металлдарды байланыстырып
антитоксикалық қызмет атқарады
Эстрогендер – тізбекті реакцияны тоқтатады: ROO– + эстрадиол-ОН →
ROOH + эстрадиол-О–– → тотығу өнімі
С,Р витаминдер
Каротиндер (вит А-ның провитамині)
Карнозин – миоциттерде гидроксил
радикалдарды бейтараптайды
Ферритин – екі валентті темірді
байланыстырады
Церулоплазмин – екі валентті мысты байланыстырады, феррооксидаздық
активтілікті көрсетіп екі валентті темірді тотықтырады
Металлотионеиндер – мысты ж\е
т.б. металлдарды байланыстырып
антитоксикалық қызмет атқарады
Эстрогендер – тізбекті реакцияны тоқтатады: ROO– + эстрадиол-ОН →
ROOH + эстрадиол-О–– → тотығу өнімі
;КАТАЛАЗА;ГЛУТАТИОНРЕДУКТАЗАГЛУТАТИОНПЕРОКСИДАЗА ФЕРМЕНТТІ ЕМЕСАНТИОКСИДАНТТАРМайда еритін витаминдер: А ж\е Е;С,Р витаминдерКаротиндер (вит А-ның")
Слайд 35Супероксиддисмутаза катализдейтін реакция:
О2- + О2- + 2Н+→ Н2О2+О2
Каталаза катализдейтін реакция:
2 Н2О2→2Н2О
+О2
Глутатионпероксидаза катализдейтін реакция:
Н2О2+ 2GSH→2Н2О + GS-SG
Тотыққан глутатион НАДФН2 мен тотығып тұруы керек
Глутатионредуктаза катализдейтін реакция:
GS-SG +НАДФН2→2GSH+НАДФ
Глутатионпероксидаза катализдейтін реакция:
Н2О2+ 2GSH→2Н2О + GS-SG
Тотыққан глутатион НАДФН2 мен тотығып тұруы керек
Глутатионредуктаза катализдейтін реакция:
GS-SG +НАДФН2→2GSH+НАДФ


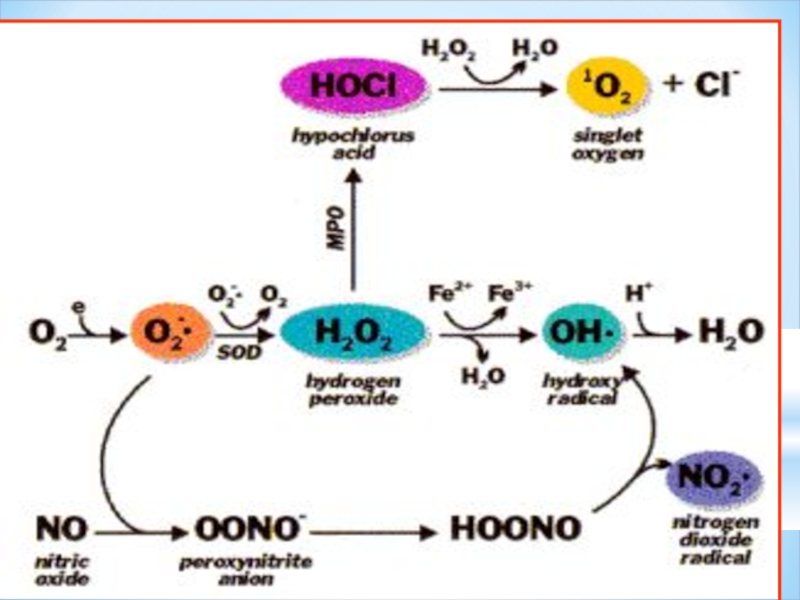
Слайд 38белоктардың SH тобына әсер етіп , оларың денатурациясын ж/е ферменттердің белсенділігін
төмендетеді;
жасуша мембранасының қызметтерін бұзады ;
изменяется ионная проницаемость (ионндық каналдар мен ионндық насостар бұзылады, мысалы,Са2+ каналы, Nа,К-АТФазы)
Ядроға (нуклеин қышқылдарына) әсер етіп жасушаның гендік аппаратын зақымдайды(мутация);
митохондрияға әсер етіп АТФ синтезін бұзады;
усиленная генерация свободных радикалов
кислорода сопровождает болезни Паркинсона,
Альцгеймера и сам процесс старения , ведет к
появлению катаракты
жасуша мембранасының қызметтерін бұзады ;
изменяется ионная проницаемость (ионндық каналдар мен ионндық насостар бұзылады, мысалы,Са2+ каналы, Nа,К-АТФазы)
Ядроға (нуклеин қышқылдарына) әсер етіп жасушаның гендік аппаратын зақымдайды(мутация);
митохондрияға әсер етіп АТФ синтезін бұзады;
усиленная генерация свободных радикалов
кислорода сопровождает болезни Паркинсона,
Альцгеймера и сам процесс старения , ведет к
появлению катаракты
БОС РАДИКАЛДАРДЫҢ (ЛПТ) ЗИЯНЫ

Слайд 39
БИОМЕМБРАНАДАҒЫ КҚМҚ-ын ыдыратып жасуша н/е органеллаларды зақымдайды,
ЯДРО,МИТОХОНДРИЯ ,
БЕЛОК ,
НУКЛЕИН ҚЫШҚЫЛДАРЫ, ЛИПИДТЕР т.б.
ЗАҚЫМДАЙТЫН ЖЕРЛЕРІ
ПАЙДА БОЛАТЫН АУРУЛАР
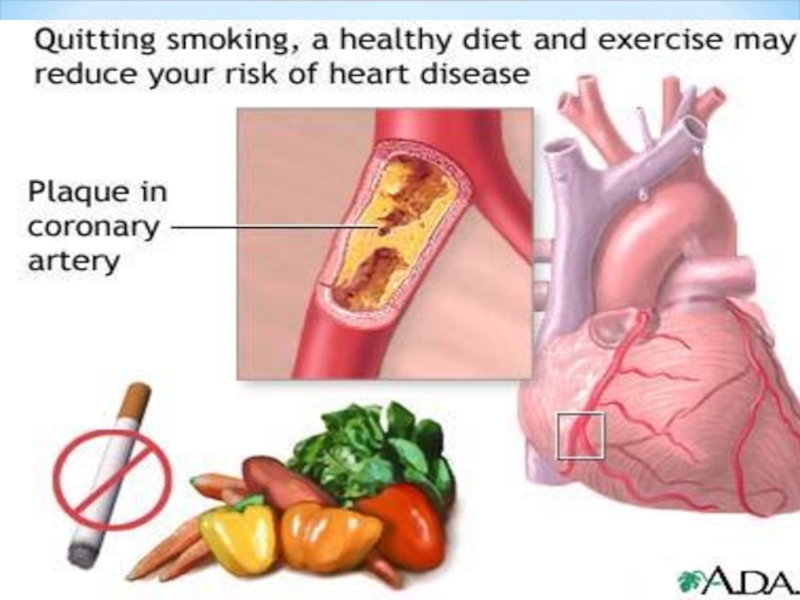
Бос радикалдар
АТЕРОСКЛЕРОЗ ҚАРТАЮДЫ;
әртүрлі РАК аурулары;
ҚАНТТЫ ДИАБЕТ;
ЖҮРЕК ИНФАРКТЫ;
КАТАРАКТ;
ПАРКИНСОН;
БҰЛШЫҚ ЕТ ДИСТРОФИЯСЫ Ж\Е Т.Б.


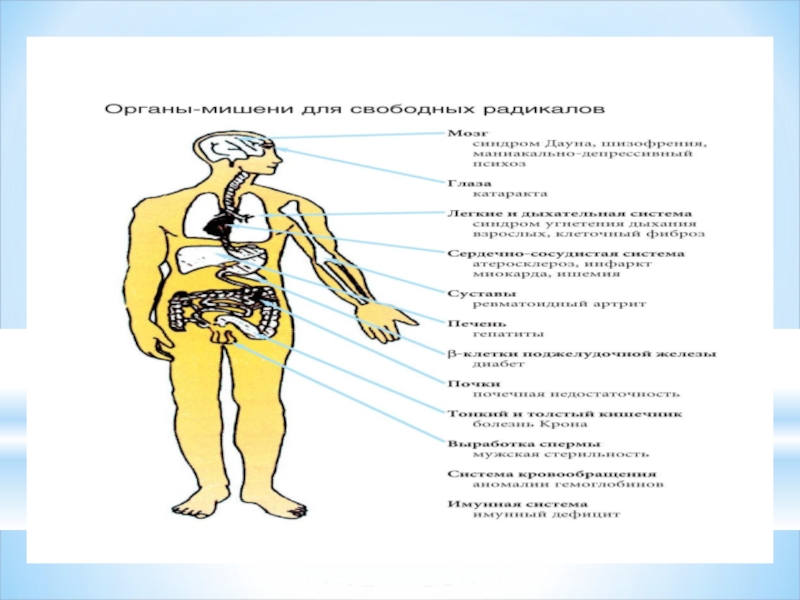
Слайд 42Бос радикалдар
жасушаларға әсер етіп, олардың ДНҚ-сын, белоктарын, майларын зақымдайтын жоғары активті
бөлшектер




Слайд 50
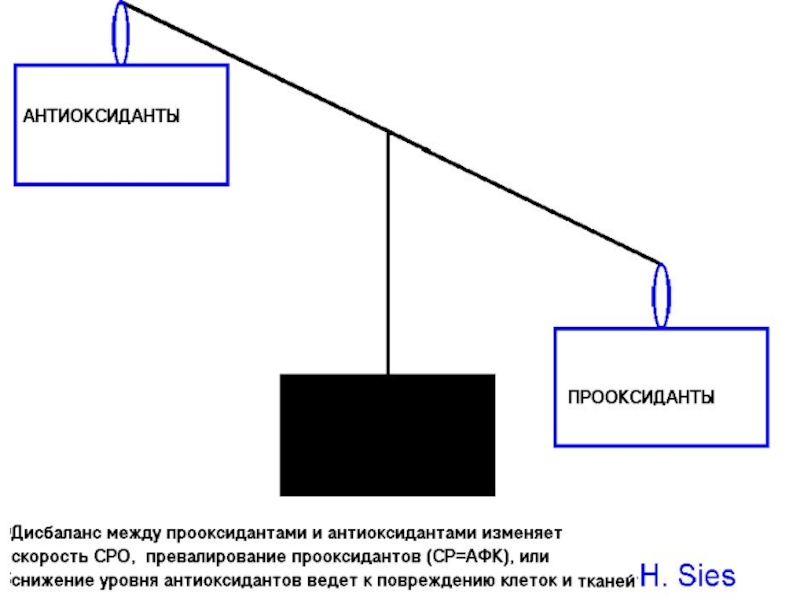
Избыток антиоксидантов вызывает прооксидантный эффект
GluSH
GS-SG
α-Tocoferol
α-Tocoferyl*
Ascorbat
Asc*
АО
АО*
е*
АФК
NADH2
NAD


Слайд 52Витамин Е антиоксид ретінде
Зақымдалған фосфолипидтер
Бос радикал
Қанықпаған майқышқылы
Қос мембрана
Нейтралданған бос радикал
Витамин Е

Слайд 53C и E как Кo-Aнтиоксиданты
Поскольку кислород реагирует с цепью ЖК, изменяется
дипольный момент и это выдаливает пероксильный радикал в плавающее состояние.
Витамин E
Lipid

Слайд 54C и E как Кo-Антиоксиданты (2)
Липид
Витамин Е
Витамин Е
удаляет пероксильный радикал и превращается сам в СР; аскорбат может восстанавливать витамин Е;
энзимы затем удаляют поврежденные ЖК и вставляют новые ЖК, репарируя липиды МЕМБРАН
энзимы затем удаляют поврежденные ЖК и вставляют новые ЖК, репарируя липиды МЕМБРАН
Гидроперекись ЖК
Р
Е
П
А
Р
А
Ц
И
Я
ЛипидВитамин Е Витамин Е удаляет пероксильный радикал и превращается")

Слайд 57ВОЗМОЖНО ЛИ ПРЕДОТВРАТИТЬ ИЛИ УДАЛИТЬ СШИВКИ?
Вещества, используемые для предотвращения и удаления
сшивок в коже.
Карнозин
Карнозин предотвращает образование в коллагене сшивок, ведущих к потере эластичности кожи и морщинам.
Карнозин обладает фантастической способностью к омоложению стареющих клеток и увеличению предела их деления.
Карнозин способен улучшать заживление ран.
Карнозин помогает макрофагам лучше узнавать AGE молекулы, способствуя тем самым их удалению.
Метформин
Аминогуанидин
Гуанидин
Молодые
фибробласты
Старые
фибробласты
Старые фибробласты
после обработки карцинином (β-аланил-гистамином)

Слайд 58Оксигеназды тотығу
эндоплазмалық ретикулум мембрансы (микросома) ж/е митохондрия мембранасының ішкі жағында жүреді.
Оксигеназды
тотығу 2 түрлі жолмен жүреді: монооксигеназды диоксигеназды.
 ж/е митохондрия мембранасының ішкі жағында жүреді.Оксигеназды тотығу 2 түрлі жолмен")
Слайд 59Монооксигеназдық тотығуда оттектің бір атомы субстратпен байланысады ал 2-сі су молекуласын
түзуге жұмсалады:
RH+O2+NADPH2→ROH+H2O+ NADP+
Диоксигеназдық тотығуда оттектің
2атомы да субстратқа байланысады:
S + O2 → SО2
қанықпаған май қышқылдар
RH+O2+NADPH2→ROH+H2O+ NADP+
Диоксигеназдық тотығуда оттектің
2атомы да субстратқа байланысады:
S + O2 → SО2
қанықпаған май қышқылдар

Слайд 60Реакция микросомаларда, әсіресе залалсыздандыру бауырда жүреді. Мембранамен байланысқан НАДФН2, ФП ж\е
цхР450 мультиферментті жүйе қатысады.
Он не дает клетке энергии
Ферменты: оксигеназы
• диоксигеназы, • монооксигеназы (гидроксилазы).
Он не дает клетке энергии
Ферменты: оксигеназы
• диоксигеназы, • монооксигеназы (гидроксилазы).

Слайд 61Оксигеназды тотығудың маңызы
Бұл процестің қатысуымен
Д витаминнің ауыспалы активті түрлері (1,25
(ОН)2Д3), стероидты гормондардың өт қышқылдарының синтезі жүреді, кейбір эндогенді және экзогенді улы заттар (дәрілер) залалсызданады.
2Д3), стероидты гормондардың")
Слайд 62Микросомальное окисление - совокупность реакций первой фазы биотрансформации ксенобиотиков и эндогенных
соединений, катализирующихся ферментными системами мембран эндоплазматического ретикулума гепатоцитов при участии цитохрома - совокупность реакций первой фазы биотрансформации ксенобиотиков и эндогенных соединений, катализирующихся ферментными системами мембран эндоплазматического ретикулума гепатоцитов при участии цитохрома Р-450. При дифференциальном центрифугировании эндоплазматический ретикулум оказывается в микросомальной фракции, поэтому эти реакции получили название микросомальных, а соответствующие ферменты - микросомальных оксигеназ.
Суть реакций заключается в гидроксилировании вещества типа R-H с использованием одного атома молекулы кислорода О2, второй атом соединяется с протонами водорода H+ с образованием воды. Донором протонов водорода является восстановленный с образованием воды. Донором протонов водорода является восстановленный NADPH + H+. Таким образом, меняется структура исходного вещества, а значит и его свойства, причём они могут как угнетаться, так и наоборот, усиливаться. Гидроксилирование позволяет перейти процессу обезвреживания ко второй фазе — реакциям конъюгации, в ходе которых к созданной функциональной группе будут присоединяться другие молекулы эндогенного происхождения.
Суть реакций заключается в гидроксилировании вещества типа R-H с использованием одного атома молекулы кислорода О2, второй атом соединяется с протонами водорода H+ с образованием воды. Донором протонов водорода является восстановленный с образованием воды. Донором протонов водорода является восстановленный NADPH + H+. Таким образом, меняется структура исходного вещества, а значит и его свойства, причём они могут как угнетаться, так и наоборот, усиливаться. Гидроксилирование позволяет перейти процессу обезвреживания ко второй фазе — реакциям конъюгации, в ходе которых к созданной функциональной группе будут присоединяться другие молекулы эндогенного происхождения.

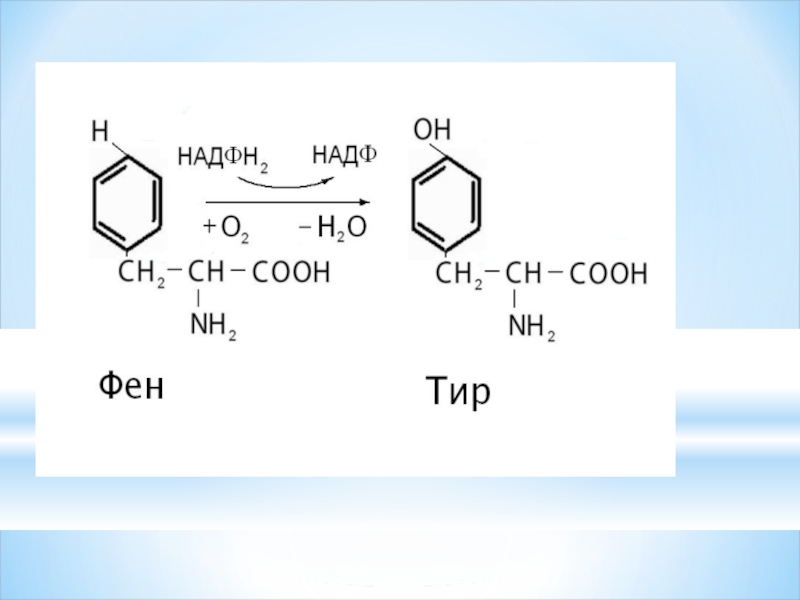
Слайд 65Пероксидаздық тотығу
Бұл процесс пероксисомаларда жүреді. Бұл процесте Н2О2 түзіледі.
Пероксидаздық тотығуға АҚ-ның,
биогенді аминдердің, пуриндердің оксидазалары қатысады. Олар негізінен флавинферменттер болып келеді.
SН2 +ФП ФПН2 + S
ФПН2+О 2 ФП+Н2О2
SН2 +ФП ФПН2 + S
ФПН2+О 2 ФП+Н2О2

Слайд 66Пероксидаздық тотығу
ксантиноксидаза
Ксантин + Н2О + O2 --------------- несеп қышқылы + Н2О2
каталаза
2 Н2О2 ---------------- 2Н2О + O2
Ксантин + Н2О + O2 --------------- несеп қышқылы + Н2О2
каталаза
2 Н2О2 ---------------- 2Н2О + O2
Оттектің 2% тотықсызданған ФП(ФАД)-ны тотықтыруға кетеді, қосымша сутек пероксиді түзіледі, оны каталаза ыдыратады.
ФПН2 + О2 ---- ФП + Н2О2

Слайд 67
Негзгl ж/е косымша әдебиеттер
1.Березов Т.Т., Коровкин Б.Ф. «Биологическая химия», 2002 -
С. 188-200, 363-406.
2. Плешкова С.М.,Омирзакова К.К «ЗАТТАР АЛМАСУЫ Ж/е ОНЫН РЕТТЕЛУ1», 2006ж-С.45-55, 71
3. Аблаев Н.Р. «Биохимии в рисунках и схемах» 2006 г - С. 41-56.
4. Сеитов З.С. «Биохимия», 2000 - С. 523-566.
5. Зайчик А.Ш., Чурилов Л.П. «Основы патохимии»2000 - С. 132-209.
6. Бышевский А.Ш., Терсенов О.А. «Биохимия для врача» 1994 - С.313-322,231,94-105.
7. Harper's Biochemistry - R.K. Murray, D.K. Granner, P.A. Mayes, V.W.
Rodwell - APPLETON&LANGE, Stamford, Connecticut, 2004
8. Биохимия человека - P. Марри, Д. Греннер, П. Мейес, В. Родуэлл- М.,Мир, 1993
9. Шарманов Т.Ш., Плешкова СМ. - Метаболические основы питания с курсом общей биохимии - Алматы, 1998






