- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Acizii nucleici презентация
Содержание
- 1. Acizii nucleici
- 2. Obiectivele: Tipurile de acizi nucleici, funcţiile şi
- 3. Acizi nucleici Acizi nucleici –sunt polinucleotide, alcătuite
- 4. ADN Localizarea: 97-99% - concentrat în
- 5. ARN Localizarea: 11% - în nucleu 15%
- 6. ARN mesager (mARN) constituie 25% din
- 7. ARN de transport (tARN) constituie 15%
- 8. Structura chimică a AN La hidroliză AN
- 9. Bazele azotate a. BA se clasifică în
- 10. Bazele purinice
- 11. Bazele pirimidinice
- 13. Molecular Biology Bicyclic Purines: Thymine (T) is
- 14. Structura BA minore
- 15. Molecular Biology C1 Nucleic Acid Structure-2 Nucleosides The structures of pentose sugar
- 16. Nucleozidul constă dintr-o BA ( purinică sau
- 17. Nucleozidele Proprietăţile: Mai solubile în H2O
- 18. NUCLEOTIDE - compuşi alcătuiţi din nucleozide
- 19. Nucleotide - Rolul Element structural al AN
- 20. Molecular Biology A nucleotide is
- 21. Structura chimică
- 24. Structura primară a AN Reprezintă secvenţa mononucleotidelor
- 27. Structura secundară a ADN Watson şi Crick
- 28. Structura secundară a ADN 3. lanţurile sunt
- 30. Legităţile lui Chargaff Conţinutul adeninei este egal
- 31. Există diferite forme de DNA care sunt
- 33. Structura terţiară Reprezintă superspiralizarea dublului helix la
- 34. 30 nm Solenoid ~40 / 50 DNA
- 35. Structura secundară şi terţiară a ARNm ARNm
- 37. Structura secundară a t-RNA are infăţişarea unei
- 38. Structura terţiară a tRNA Are forma L
- 39. Structura terţiară a tRNA Are forma L
- 40. ARNr Structura secundară – e prezentată prin
- 41. Proprietăţile fizico-chimice ale acizilor nucleici - masa
- 42. Denaturarea şi renaturarea Denaturarea –sub acţiunea temperaturii,
- 43. Hibridizarea AN Pe capacitatea de renaturare a
- 44. Această metodă a permis descoperirea particularităţilor
- 46. Obiectivele: Dogma centrală a geneticii moleculare. Concepţia:
- 47. Dogma centrală a geneticei moleculare Postulatul de
- 48. Dogma centrală a geneticii moleculare
- 49. Structura genelor- dimensiuni, GS; GR Gene- porţiunile
- 50. Replicarea Replicarea – transmiterea informaţiei genetice
- 52. Componentele necesare replicării: Matriţă -
- 53. d. ADN polimeraza ( ADNp I,
- 54. Mecanismul replicării 3 etape: iniţierea, elongarea, terminarea
- 55. b. Sinteza primerului - sub acţiunea
- 56. Elongarea ADN polimeraza III unindu-se la capătul
- 57. Terminarea Terminarea replicării are loc atunci, cînd
- 60. Replicarea la eucariote Particularităţi: ADN polimerazele: αβγδ
- 61. Telomer Telomeraza Replicarea capetelor 5’ ale catenelor
- 62. Telomeraza – fiind o revertază (ADN
- 63. Mecanismul elongării capetelor cromozomului la eucariote
- 65. Mecanismul elongării capetelor cromozomului la eucariote
- 66. Structura şi funcţia RNA telomerazice. Structura
- 67. Inhibitorii telomerazei oligonucleotidele modificate, complementare regiunii
- 68. La om telomeraza e activă numai
- 69. Numărul telomerilor determină durata vieţii fiecărei
- 70. Reparaţia ADN Erorile în timpul replicării sunt
- 71. Reparaţia ADN Incizia dimerului sub acţiunea endonucleazelor
- 72. Reparaţia prin excizia dimerului
- 73. Reparaţie prin fotoreactivare
- 74. Reparaţie prin recombinare
- 75. Transcripţia biosinteza ARN pe matriţă de ADN
- 76. ARN polimeraza este o holoenzimă
- 77. Etapele transcripţiei Sinteza decurge în 3 etape:
- 78. Fig. 5.4
- 79. Etapele transcripţiei Sinteza decurge în 3 etape:
- 80. Sigma subunitatea găseşte punctul de iniţiere
- 81. Elongarea şi Terminarea Elongarea - alunecarea ARN
- 83. Transcripţia la eucariote RNA p alcătuită din
- 84. Procesingul Toţi precursorii de ARN în nucleu
- 85. Fig. 5.11
- 86. 2.Splisingul - excizia intronilor şi sudarea
- 87. Fig. 5.13
- 88. e.g., Fig. 5.13
- 90. Transcripţia inversă sinteza ADN pe catena
- 91. Codul genetic Translaţia Reglarea sintezei proteinei
- 92. Obiectivele: Codul genetic. Proprietăţile. Ribozomii - sediul
- 93. Codul genetic Informaţia genetică referitor la biosinteza
- 94. Proprietăţile codului genetic Este triplet -64
- 95. Este universal – toate veţuitoarele utilizează
- 96. Ribozomii Reprezintă sediul de traducere a ARNm
- 97. Centrele catalitice ale ribosomilor Situsul A -
- 98. Mai mulţi ribozomi pot citi simultan
- 99. Translaţia Translaţia sau biosinteza proteinelor propriu zisă.
- 100. Etapele se realizeaza in 5 etape:
- 101. Activarea AA are loc în citozol Sunt
- 102. Se desfăşoară în două etape:
- 103. Activarea AA Parcurge în 2 etape:
- 104. Activarea AA Esenţa procesului de activare este
- 105. Activarea AA Activarea AA consumă 2 legături
- 107. Translaţia propriu zisă Citirea ARNm se face
- 108. Iniţierea Necesar: ARNm (AUG) Ribosomul cu subunităţile
- 109. Formarea complexului de iniţiere: Subunitatea mică leagă
- 111. Elongarea Necesar: ARNm cu următorul codon ARNt
- 112. 1. Adaptarea (legarea)AA are loc după principiul
- 114. 2. Transpeptidarea este formarea legăturii peptidice între
- 116. 3. Translocarea deplasarea ARNm cu un triplet
- 117. Elongarea Decurge în 3 etape :1 fixarea
- 118. Terminarea are loc cînd sunt întîlniţi codonii
- 119. La formarea unei legături peptidice se
- 120. Prelucrările posttraducere Modificarea capătului N- şi C-terminal;
- 121. Inhibitori ai sintezei proteinei la nivelul
- 122. Inhibitorii sintezei proteinei la nivelul translatiei:
- 123. Reglarea sintezei proteinelor Sinteza proteinelor nu e
- 124. Teoria lac-operonului Schema reglării biosintezei proteinei la
- 125. Ce se întâmplă dacă bacteria dispune
- 126. Bacteria are ca sursă glucoza
- 127. Reglarea sintezei proteinei prin inducţie În prezenţa
- 128. Bacteria are ca sursă lactoza
- 129. În lipsa glucozei se măreşte c%
- 130. Reglarea lac-operonului într-un mediu ce conţine glucoză
- 131. Ilustrarea mecanismului de reglare a sintezei proteinei
- 132. Ilustrarea mecanismului de reglare a sintezei proteinei prin represie
- 134. REZUMĂM: GR controlează exprimarea anumitor GS prin
- 135. Reglarea sintezei la eucariote Atât la nivelul
- 136. Ingineria genetică ştiinta, preocupată de crearea
- 137. Căpătarea ADN-ului recombinat- gena necesară se întroduce
- 138. 3.Clonarea ADN-ului recombinat- obtinerea cantităţilor dorite de
- 139. Ranadamentul sintezei bacteriene este impresionabil: Ex- 100
- 140. Mutaţiile. Modificările genomului organismului, care se
- 141. Mutatiile punctiforme: pot decurge prin: l.
- 143. Secvenţa de AA în porţiunea variabilă este
- 144. Sinteza Anticorpilor Fiecare dintre milioanele de
- 145. Gradul de diversificare în obţinerea lanţurilor

Слайд 2Obiectivele:
Tipurile de acizi nucleici, funcţiile şi repartizarea lor în celulă.
Constituienţii acizilor
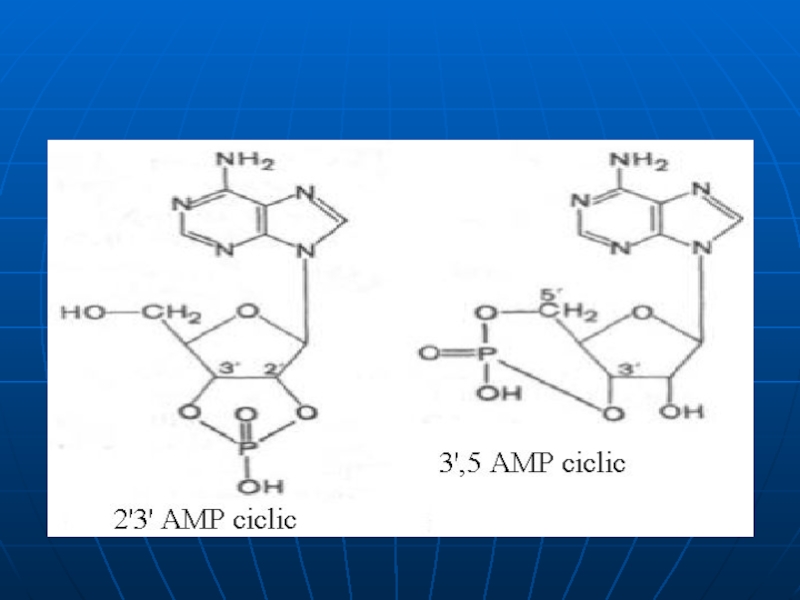
Nucleozidele şi nucleotidele. 3′, 5′- cAMP
Structura primară, secundară şi terţiară a acizilor dezoxiribonucleici Cromatina. Nucleosomul.
Structura acizilor ribonucleici (tRNA, mRNA, rRNA).
Denaturarea şi hibridizarea acizilor nucleici.

Слайд 3Acizi nucleici
Acizi nucleici –sunt polinucleotide, alcătuite din mononucleotide, unite prin legături
ADN - acidul dezoxiribonucleic;
ARN - acidul ribonucleic.

Слайд 4ADN
Localizarea:
97-99% - concentrat în nucleu
1-3% - situat în mitocondrii.
Rolul: păstrează şi transmite informaţia genetică de la ADN parental la ADN fiică sau ARN.

Слайд 5ARN
Localizarea:
11% - în nucleu
15% -în mitocondrii
50% - în ribosomi
24% - în
Deosebim:i:
ARN mesager
ARN ribozomal
ARN de transport
ARN cromosomial
ARN nuclear

Слайд 6
ARN mesager (mARN) constituie 25% din totalul ARN-lui.
Localizat -în nucleu
Prezintă copia sectorului de ADN şi conţine informaţia despre structura catenei polipeptidice a proteinei.
Rolul:Transmite informaţia de la ADN spre ribozomi, sediul de sinteză a proteinei.
ARN ribozomal (rARN) constituie 60% din totalul ARN-ului.
Localizat- în ribozomii citoplasmei.
Rolul - formează scheletul ribozomilor. Joacă un rol auxiliar în procesul de asamblare a proteinelor.
 constituie 25% din totalul ARN-lui. Localizat -în nucleu şi citozol. Prezintă copia")
Слайд 7
ARN de transport (tARN) constituie 15% din totalul ARN-lui.
Localizat: în
Rolul: participă la activarea şi transportul AA spre ribozomi şi asamblarea lor în polipeptide.
ARN cromosomial – activarea genelor ADN
ARN nuclear – formarea scheletelor particulei proteice care transportă ARN din nucleu în citoplasmă
 constituie 15% din totalul ARN-lui. Localizat: în citoplasmă, ribosomi, mitocondrii.Rolul: participă")
Слайд 8Structura chimică a AN
La hidroliză AN degradează în mononucleotide, care la
ADN----A; G; C; T+dR+H2PO3
ARN----A; G; C; U+ R+H2PO3

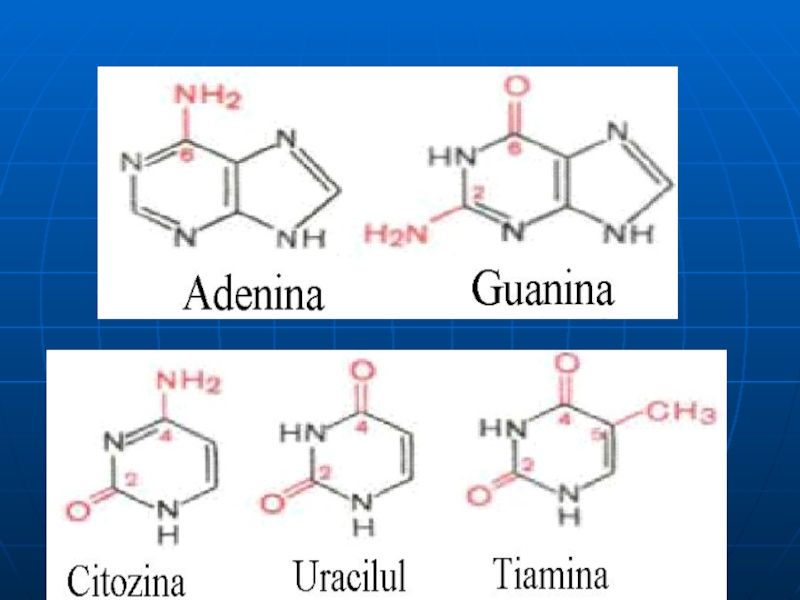
Слайд 9Bazele azotate
a. BA se clasifică în :
majore: purinice: A, G
minore:purinice (2metil A; 1 metilG) şi
pirimidinice (5 metil C;5 hidroximetil C)
b. Sunt slab solubile în H2O
c. Prezintă fenomenul de tautomerie (forme lactim-lactam)
d. Sunt responsabile de informaţia genetică
e. BA purinice- au structură plană; cele pirimidinice- aproape plană, puţin plată
j. Max capacităţii de absorbţie în ultraviolet este între 260-280 nm



Слайд 13Molecular Biology
Bicyclic Purines:
Thymine (T) is a 5-methyluracil (U)
C1 Nucleic Acid Structure-1
Bases
Monocyclic
pyrimidine:
 is a 5-methyluracil (U)C1 Nucleic Acid Structure-1BasesMonocyclic pyrimidine:")


Слайд 16Nucleozidul
constă dintr-o BA ( purinică sau
Pirimidinică) +
o pentoză (riboza sau
atomul C-1 al pentozei este unit cu N-9 al purinei sau N-1 al pirimidinei - leg. N glicozidică.
În funcţie de pentoză: dezoxi şi ribonucleozide
BA purinice +R(dR) --- ozin
(adenozin, guanozin
sau dezoxiadenozin, dezoxiguanozin)
BA pirimidinice +R (dR) --- idin
(citidin, timidin, uridin sau dezoxicitidin)
Unite între ele prin
legătura N glucozidică
 +o pentoză (riboza sau dezoxiriboza)atomul C-1 al pentozei")
Слайд 17Nucleozidele
Proprietăţile:
Mai solubile în H2O decât BA
Mai stabile în soluţii alcaline
Uşor

Слайд 18
NUCLEOTIDE - compuşi alcătuiţi din nucleozide şi rest de acid fosforic
Nucleozid

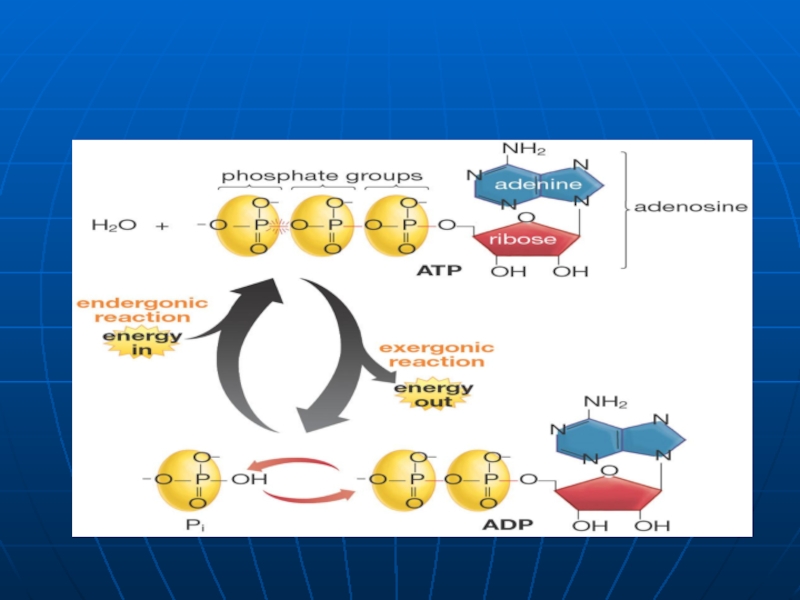
Слайд 19Nucleotide - Rolul
Element structural al AN
Intermediari energetici (ATP- purtătorul energiei chimice
Intră în componenţa Co
Servesc ca activatori ai unor molecule (UDP-Gl; CDP-colina)
Servesc ca mesageri secunzi intracelulari ai hormonilor (AMPc; GMPc)
Intră în componenţa")
Слайд 20Molecular Biology
A nucleotide is a nucleoside with one or
Deoxyribonucleotides
(containing deoxyribose)
Ribonucleotides
(containing ribose)
Phosphate ester bonds
C1 Nucleic Acid Structure-3
Nucleotides


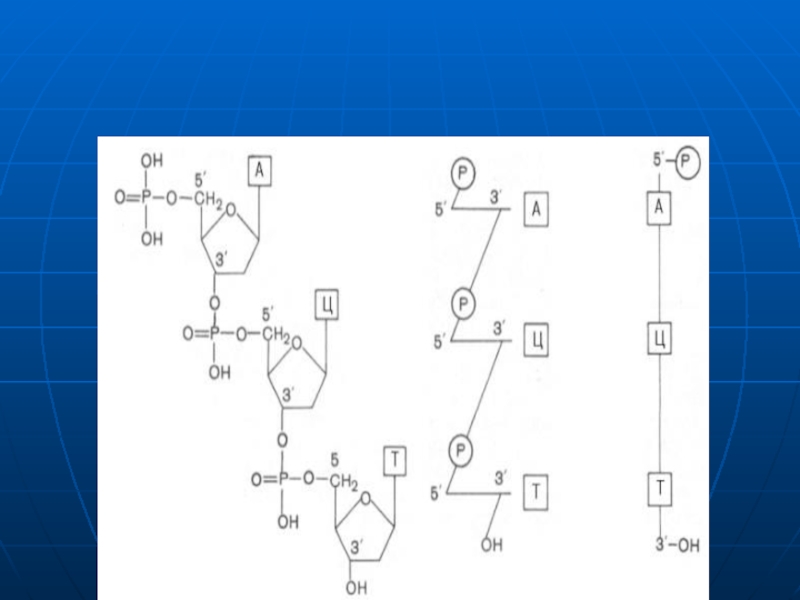
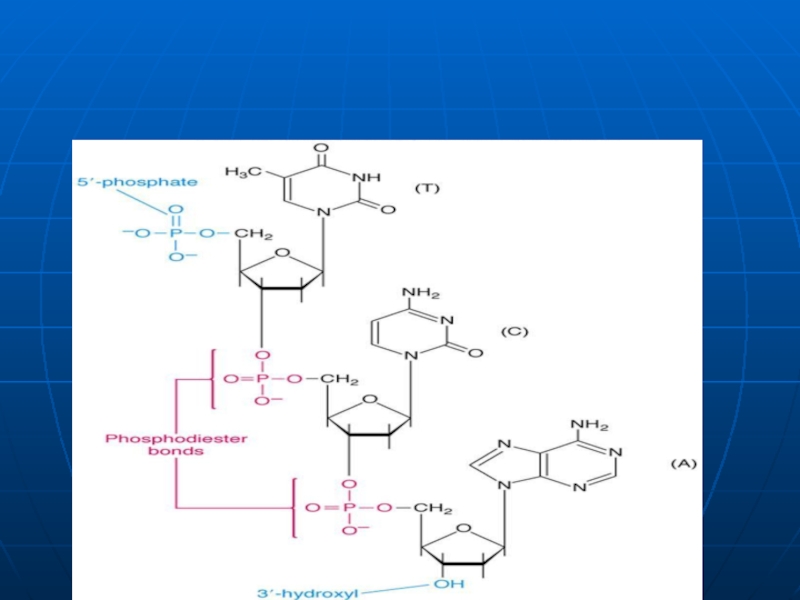
Слайд 24Structura primară a AN
Reprezintă secvenţa mononucleotidelor în lanţul polinucleotidic liniar, legate
Catenele au două capete:
5‘ – nucleozid tri fosfatul;
3‘ – gr. OH liberă

Слайд 27Structura secundară a ADN
Watson şi Crick (1953) au postulat modelul structural
Caracteristicile dublei spirale:
2 lanţuri polidezoxiribonucleotidice se răsucesc helicoidal în jurul unui ax comun, formând o dublă helice cu orientare spre dreapta;
Cilindrul ce încadrează dublul helix are d=2nm
 au postulat modelul structural al moleculei de DNA")
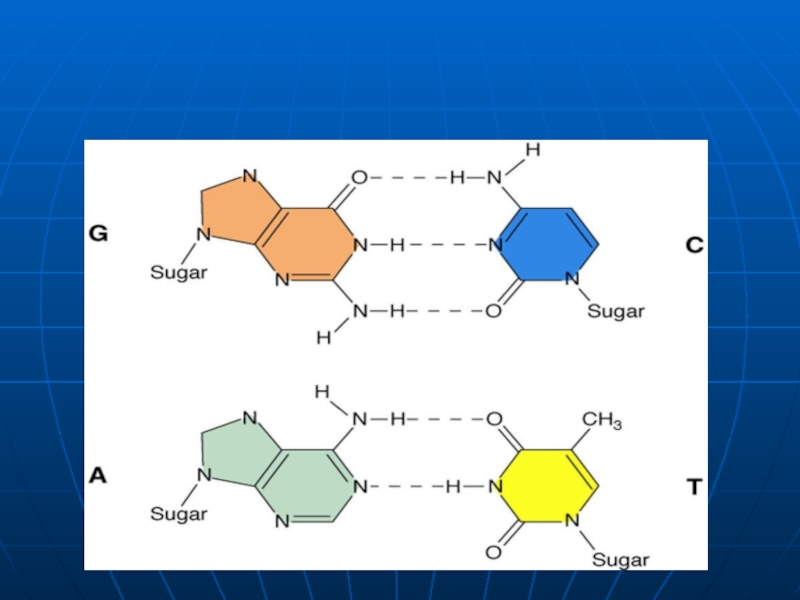
Слайд 28Structura secundară a ADN
3. lanţurile sunt antiparalele (unul are direcţia 5’→3’,
4. complimentaritatea (A îi corespunde T; iar G-C).
5. Stabilitatea dublului helix este asigurată atât de interacţiunile hidrofobe dintre BA, cât şi de legăturile de hidrogen între BA (A=T formează 2 legături de hidrogen, iar G ≡C trei legături).
6. BA hidrofobe sunt situate în interiorul spiralei duble şi aranjate sub formă de stive, pe cînd complexul pentozofosfat este situat la exteriorul spiralei duble, bine interacţionează cu apa, de aceia molecula gigantă de DNA se dizolva în apă.
7. Spirală este regulată (fiecare spiră cuprinde 10 nucleotide). Distanţa dintre BA învecinate este de 0,34 nm, perioada de identitate (pasul) – 3,4 nm.
8. La pH=7 grupele fosfat sunt ionizate, poarta sarcini negative, deaceia DNA prezintă acid puternic.
9. Dublul helix este de tip plectonemical, dar nu paranemical
4. complimentaritatea (A")
Слайд 30Legităţile lui Chargaff
Conţinutul adeninei este egal cu al timinei, iar al
În orice preparat de DNA independent de specie suma bazelor purinice este egală cu cea a bazelor pirimidinice (A+G=T+C)
Preparatele de DNA separate din diferite ţesuturi a uneia şi aceeiaş specie de organisme sunt absolut identice privind componenţa nucleotidică.
Componenţa nucleotidică a DNA la aceeaşi specie nu se modifică odată cu vârstă, nu depinde de regimul alimentar şi modificările mediului.
dacă A+T este mai mare decît G+T avem DNA de tip AT
dacă G+T este mai mare decît A+T avem DNA de tip GT
t de topire este mai mica cînd predomină perechile A-T
t de topire este mai mare cînd predomină perechile G-C
la eucariote DNA mitocondrial este circular

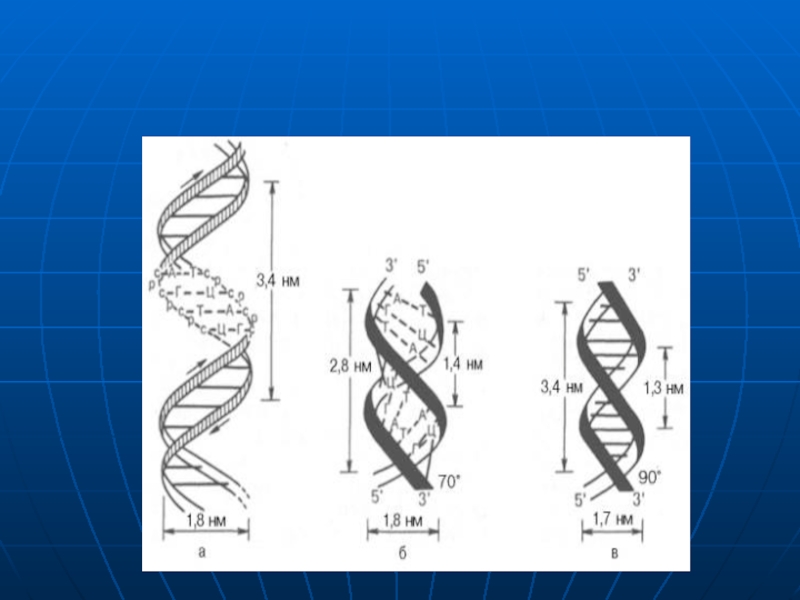
Слайд 31Există diferite forme de DNA
care sunt determinate de gradul de
dehidratare
dependente de anturajul extern ai
moleculei de DNA.
Dublul helix posedă dinamism.
forma A:
conţine 11 resturi la o spiră,
este răsucită spre dreapta.
forma clasica B:
conţine 10 mononucleotide la o spiră.
este răsucită spre dreapta.
- 10 nucleotide ocupă 34 A (3,4 nm).
- o nucleotidă cuprinde 3,4A (0,34 nm).•
conformatia Z spre deosebire de A
şi B este răsucită spre stînga.

Слайд 33Structura terţiară
Reprezintă superspiralizarea dublului helix la care participă proteinele histonice şi
Unitatea structurală a cromatinei este nucleosomul
Nucleosomul – este un octamer histonic (2H2A; 2H2b; 2H3; 2H4) înfăşurat de aproximativ de 2 ori de dublul helix cu o lungime de 146 perechi de nucleotide.
Între 2 nucleosmi se conţin porţiuni de ADN alcătuit din 20-60 perechi de nucleotide asociate cu H1
Lanţul polinucleosomic formează un superhelix (solenoid), fiecare spiră are 10 nucleosomi, d=30nm şi pasul de 10nm

Слайд 3430 nm Solenoid ~40 / 50
DNA
Firul de cromatină?
~ 1,000
Cromosoma metafazică/
~ 10,000
Compactizarea
cromatinei
Nucleosoma
= оctamer de histone
H2a, H2b, H3, H4
146 / 200 bp DNA
Compactizare
~10 ori

Слайд 35Structura secundară şi terţiară a ARNm
ARNm – fiecărei gene îi corespunde
Elementul de codificare al ARNm este tripletul nucleotidic – numit codon. Fiecare codon corespunde unui anumit AA
Structura secundară a ARNm – o catenă curbată
Structura terţiară – se aseamănă cu un fir înfăşurat pe bobină, rolul căreia îl îndeplineşte o proteină de transport numită informer


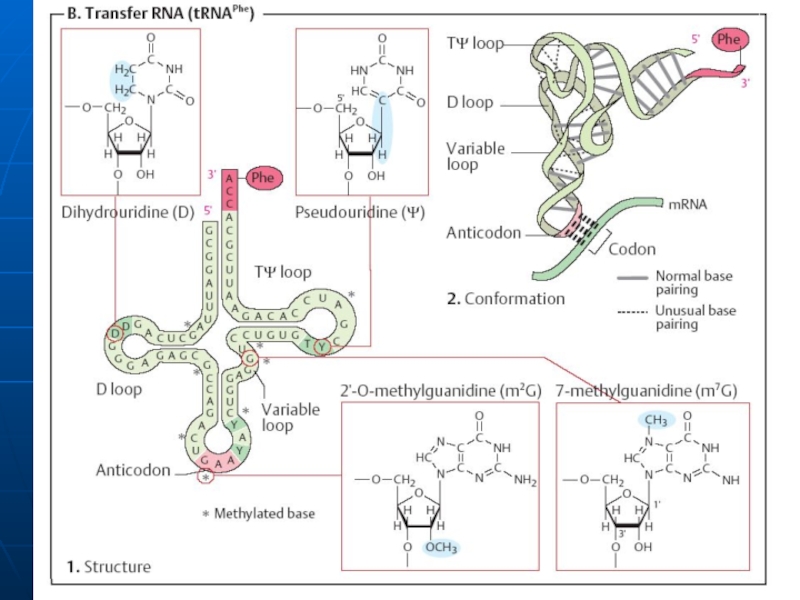
Слайд 37Structura secundară a t-RNA
are infăţişarea unei "frunze de trifoi" - se
Sectoarele, care nu sunt încadrate în formarea legăturilor de H formează lanţuri sau bucle
Sectorul de acceptare (4 nucleotide, 3 din care au aceeaşi succesiune- CCA cu hidroxilul 3’OH liber la care se fixează grupa COOH al AA.
Bucla anticodonică - formată din 7 nucleotide. Conţine un triplet nucleotidic specific pentru fiecare t-RNA numit anticodon. Anticodonul t-RNA după principiul complementaritătii se împerechează cu codonul respectiv din RNAm. Interacţiunea codon-anticodon determină ordinea aranjării aminoacizilor în catena polipeptidică.
Bucla pseudouridilică - constă din 7 nucleotide (restul acidului pseudouridilic este obligatoriu), participă la interacţiunea cu ribozomii.
Bucla dihidrouridinică - constă din 8-12 resturi nucleotidice ( resturi de dihidrouridină ), interactiunează cu E- aminoacil-RNAt-sintetaza, care contribuie la recunoaşterea de către aminoacid a ARN-t specific.

Слайд 38Structura terţiară a tRNA
Are forma L
Include 2 segmente de dublu helix
În afara spiralei bazele formează legături de hidrogen. Interacţiuni apar între bazele necomplementare (A-A; A-C).Multe baze sunt aranjate în stive (hidrofobe)

Слайд 39Structura terţiară a tRNA
Are forma L
Include 2 segmente de dublu helix
În afara spiralei bazele formează legături de hidrogen. Interacţiuni apar între bazele necomplementare (A-A; A-C).Multe baze sunt aranjate în stive (hidrofobe)

Слайд 40ARNr
Structura secundară – e prezentată prin sectoare spiralate unite între ele
Structura terţiară – prezintă scheletul ribosomului. Are forma unui bastonaş sau ghem pe suprafaţa căruia sunt înfăşurate proteinele ribosomului.

Слайд 41Proprietăţile fizico-chimice ale acizilor nucleici
- masa moleculară mare.
proprietatile coloidale si osmotice,
Proprietăţile lor hidrofile depind de fosfaţi.
viscozitatea şi densitatea înaltă a soluţiilor,
capacitatea de denaturare.
la pH fiziologic toti AN sunt polianioni (-)

Слайд 42Denaturarea şi renaturarea
Denaturarea –sub acţiunea temperaturii, mediului PH, substanţelor chimice are
La denaturare DNA îşi pierde proprietăţile biologice. Ex. încălzirea DNA-duce la desfacerea spiralei duble în două catene ( are loc transformarea „spirală - ghem“).
Degradarea unei jumătăţi de structură de ADN are loc la temperatura de topire. ADN bogat în C şi G au o t mai înaltă decît cele bogate în A şi T.
La răcirea treptată catenele din nou se reunesc după principiul complementaritătii, formînd spirala dublă nativă. Acest fenomen se numeşte renaturare (atunci cînd t e mai mică decît cea de topire).
La racirea bruscă renaturarea nu are loc.
Denaturarea şi renaturarea acizilor nucieici este însoţită de schimbarea activitătii lor optice.

Слайд 43Hibridizarea AN
Pe capacitatea de renaturare a AN este bazată metoda de
La baza ei stă împerecherea complementară a sectoarelor unicatenare ale AN cu formarea unui heteroduplex
Hibridizarea se efectuează în felul următor:
AN se denaturează separat;
se incubează împreună ambele tipuri de DNA (ori DNA şi RNA).
În condiţiile unui grad relativ crescut de complementaritate a acestora se formează moleculele hibride (DNA-DNA sau DNA-RNA). Aceste molecule constau din sectoare spiralate şi nespiralate. Cu cît gradul de înrudire este mai înalt, cu atît hibridizarea este mai complectă.

Слайд 44
Această metodă a permis descoperirea particularităţilor structurii primare a DNA. S-a


Слайд 46Obiectivele:
Dogma centrală a geneticii moleculare. Concepţia: o genă - un polipeptid.
Replicarea
Telomeraza. Rolul şi structura..
Reparaţia ADN.
Transcripţia sau biosinteza ARN: matricea, substratele, enzimele, mecanismul
Trsanscripţia inversă.
Biosinteza ARN pe matrice de ARN
Modificările posttranscripţionale (processing)
Inhibitorii sintezei acizilor nucleici.
Ingeneria genetică şi semnificaţia ei practică. Sinteza anticorpilor

Слайд 47Dogma centrală a geneticei moleculare
Postulatul de bază a geneticei moleculare a
este transmiterea informaţiei genetice de la ADN la proteină. Sînt încluse trei procese:
replicarea;
transcripţia;
translaţia.
Primele două procese au loc în nucleu, iar al treilea – în citozol.
Procesul de transcripţie este reversibil. Enzima care catalizează transcripţia inversă se numeşte revertaza (reverstranscriptaza) şi a fost descoperită la oncoviruşi. Sinteza ARN-ului pe baza ARN se numeşte replicarea ARN, ea are loc la viruşi, care nu au ADN. Procesul de translaţie este ireversibil şi se numeşte biosinteza proteinei.


Слайд 49Structura genelor- dimensiuni, GS; GR
Gene- porţiunile ADN ce conţin informaţia genetică
Fiecărei gene îi corespunde un lanţ polipeptidic- de aici şi conceptul: o genă –un lanţ polipeptidic
GS- genele ce codează polipeptide şi ARN. Porţiunile GS ce conţin informaţie (transductibile) –exoni; iar secvenţele ce nu sunt traduse în ARNm – introni
GR – segmente de ADN, repetabile, relativ mici ce au un rol reglator.
Rolul lor:
Pot fi semnale ce ne arată începutul şi sfârşitul GS
Participă în iniţierea şi terminarea transcripţiei GS
Dimensiunile genelor – f. variabile. Ex. Proteina ce conţine 350 AA--- 350X3=1050 nucleotide. Ştiind că BA sunt localizate la 0,34 nm_----
0,34 nm X 1050=357 nm =0,36μm

Слайд 50Replicarea
Replicarea – transmiterea informaţiei genetice de la ADN parental la ADN
Caracteristicile:
Se petrece în nucleu
Proces semiconservativ
se desfăşoară în trei etape : iniţiere, elongare, terminare
prezenţa praimerului este obligatorie
replicarea este cuplată cu desfăşurarea DNA parental (necesită energie)
replicarea decurge în ambele direcţii cu aceeaşi viteză.
Pe catena întîrziată se sintetizează fragmentele Okazaki.
Este bazată pe împachetarea complementară a BA
Catena-fiică este antiparalelă cu catena parentală dar nu identică după secvenţa nucleotidică
Forţa motrice a procesului este hidroliza pirofosfatului
angajeaza simultan intregul cromozom.

Слайд 52
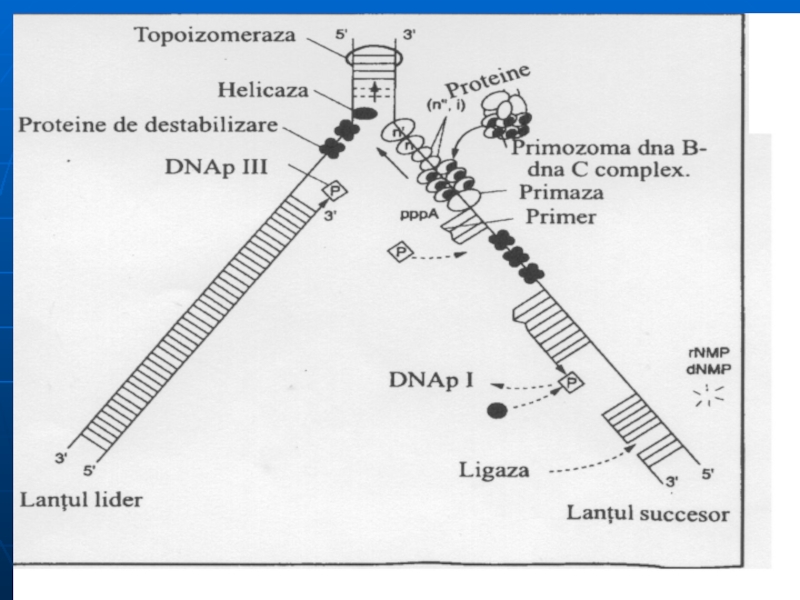
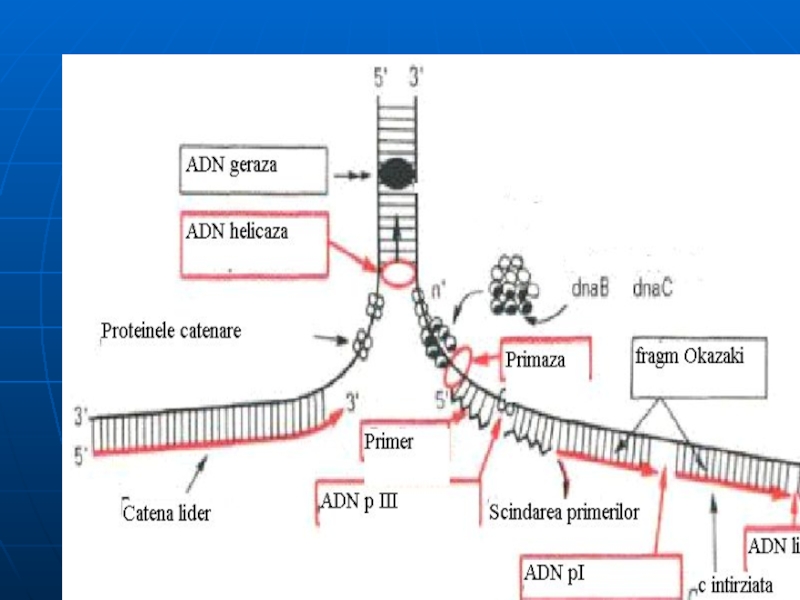
Componentele necesare replicării:
Matriţă - ADN bicatenar
Substrat: dATP, dTTP, dGTP, dCTP; ATP,
prezenţa ionilor de Mg, Mn; Zn
Sistemul multienzimatic complex:
Helicaza-desfacerea dublului helix, treptat, pe porţiuni mici. Cele 2 catene rămân separate ca urmare a intervenţiei proteinelor de stabilizare (SSB).
Topoizomerazele I şi II – înlătură supertorsiunile ADN, rezolvă problemele topologice apărute în cursul desfacerii lui. (I – introduce supertorsiuni negative; II – scindează o leg. fosfodiesterică pe una din catene şi permite celor 2 catene să se rotească una faţă de alta)
ARN primază - sintetizează primerul în direcţia 5'- 3‘ .

Слайд 53
d. ADN polimeraza ( ADNp I, II, III)- sinteza catenei fiice
ADN p III:
– acţiune polimerazică (5'- 3‘) - sintetizează în direcţia 5‘- 3' lanţul polidezoxiribonucleotidic, preluînd instrucţii de la ADN-matriţă,
- acţiune exonucleazică (3'- 5‘)
ADN pII - rol neclar.
ADN pI - posedă activitate 5'- 3‘ exonucleazică, înlătură primerul şi-l înlocuieşte cu fragmente de ADN
e. ADN ligaza- uneşte fragmentele Okazaki de pe catena întîrziată. Catalizează formarea unei legături fosfat diesterice între 3'-OH a unui fragment de ADN şi extremitatea 5' monofosfat al altuia.
- sinteza catenei fiice în direcţia 5'→3' .")
Слайд 54Mecanismul replicării
3 etape: iniţierea, elongarea, terminarea
Originea replicării este reprezentată de o
Replisoma (complex proteic) recunoaşte punctul de origine.
Iniţierea parcurge două etape:
Formarea furcii de replicaţie- ataşarea replisomului la punctul de origine al replicării şi sub acţiunea helicazelor are loc desfacerea duplexului parental pe anumite porţiuni – replicatori (la scindarea leg. de H dintre BA - se utilizează min 2 mol. de ATP). La desfacerea duplexului parental apar regiuni superhelicoidale, care se reglează cu ajutorul girazei (topoizomerazei).
Topoizomeraza efectuează rupturi monocatenare apoi sudează legătura fosfodiesterică şi favorizează relaxarea structurii DNA

Слайд 55
b. Sinteza primerului - sub acţiunea primazei se sintetizează o porţiune

Слайд 56Elongarea
ADN polimeraza III unindu-se la capătul 3' OH al primerului începe
Elongarea decurge în direcţia 5'→ 3‘, şi parcurge cu aceeaşi viteză pe ambele catene
Catena de bază se va sintetiza continuu, iar cea întîrziată - discontinuu: va fi formată din fragmente Okazaki (dimensiuni de 1000 – 2000 nucleotide la procariote şi 150-200 la eucariote).
ADN polimeraza I exclude primerii şi sintetizează complementar ADN.
Fragmentele sînt unite cu enzima ADN ligaza (necesită ATP la eucariote şi NAD la procariote).

Слайд 57Terminarea
Terminarea replicării are loc atunci, cînd cele două bifurcaţii de replicare

Слайд 60Replicarea la eucariote
Particularităţi:
ADN polimerazele: αβγδ
α- implicată în replicarea ADN nuclear- responsabilă
δ – răspunde de sinteza catenei lider. Ea manifestă acţiune exonucleazică
β – implicată în reparaţia ADN
γ - implicat în replicarea ADN mitocondrial, acţiune exonucleazică
Bifurcaţia replicii este de 3000 baze pe minut comparativ cu 16.000 la procariote
Pe o moleculă de ADN există mai multe origini de replicare (3X104 - 3X105 separate prin perechi de baze). În aceste origini multiple de replicare se organizează bifurcaţii- ce se deplasează biderecţional pe cromosomul eucariot în curs de replicare.
Fragmentele Okazaki 150-200 nucleotide

Слайд 61Telomer Telomeraza
Replicarea capetelor 5’ ale catenelor este incompletă (teoria lui Olovnicov,
Aceasta nu afectează informaţia genetică deoarece catenele conţin fragmente repetitive neinformative – telomere.
Telomerele sunt replicate de o E specifică – telomeraza
Telomeraza - reprezintă o ribonucleoproteidă: ARN şi proteină
Subunitatea proteică TRT (telomeraze revers transcriptase) posedă activitate catalitică
, deoarece după înlăturarea")
Слайд 62
Telomeraza – fiind o revertază (ADN polimeraza ARN dependentă) foloseşte ca
I etapă – are loc asocierea telomerazei la capătul 3’ al catenei lider din regiunea telomerică- TTAGGG
II – E extinde catena, utilizând ca matriţă ARN telomeric (se repetă)
III – Catena complementară a ADN telomeric e sintetizată după principiul catenei întârziate de ADNp
 foloseşte ca matriţă propria coenzimă –")


Слайд 65Mecanismul elongării capetelor cromozomului la eucariote
cromozoma
AUCCCAAUC 5’
Fixarea telomerei TTAGGG
elongarea GGGTTAGGGTTAG
5’ AUCCCAAUC
translocarea GGGTTAGGGTTAG
5’ AUCCCAAUC

Слайд 66
Structura şi funcţia RNA telomerazice.
Structura primară: la majoritatea RNA telomerice, regiunea
Structura secundară e compusă din 4 bucle şi un fragment unicatenar, ce conţine matriţa pentru sinteza DNA telomerice.

Слайд 67Inhibitorii telomerazei
oligonucleotidele modificate, complementare regiunii matrice a RNA – telomerazice.
Ca inhibitori au fost testaţi şi inhibitorii reverstranscriptazelor – azidotimidina, didezoxiguanozina.

Слайд 68
La om telomeraza e activă numai în celulele embrionale, în epiteliul

Слайд 69
Numărul telomerilor determină durata vieţii fiecărei celule şi condiţionează reducerea critică
lungimea telomerei este marcherul biologic al îmbătrînirii.

Слайд 70Reparaţia ADN
Erorile în timpul replicării sunt reduse la minimum datorită DNA
Tipuri de deteriorări:
Formarea de breşe
Modificarea BA
Pierderea de BA
Formarea dimerilor de pirimidină sub acţiunea razelor ultraviolete

Слайд 71Reparaţia ADN
Incizia dimerului sub acţiunea endonucleazelor
Peticirea – sub acţiunea ADN polimerazei
Excizia fragmentului lezat sub acţiunea exonucleazei
Sudarea – sub acţiunea ADN ligazei




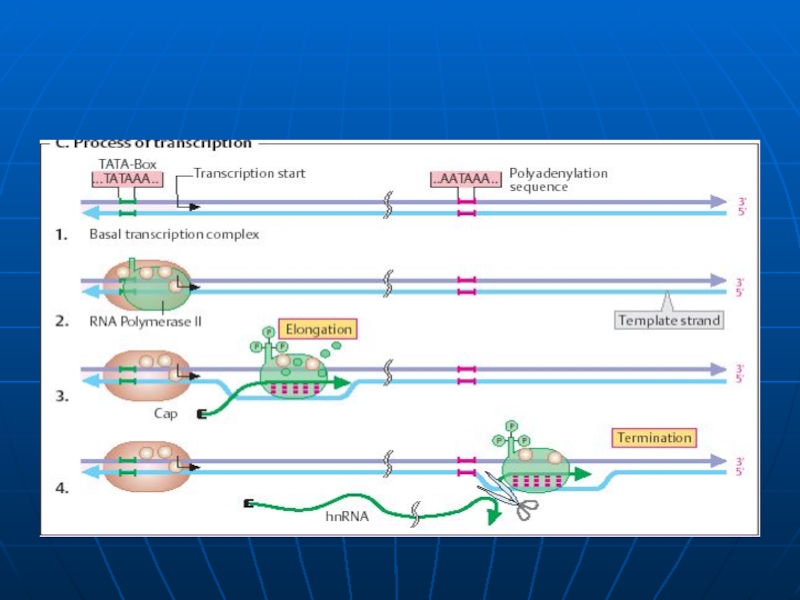
Слайд 75Transcripţia
biosinteza ARN pe matriţă de ADN
Particularităţi:
Matriţă - DNA dublu helicoidal (prezenţa
Substrat - ribonucleozidtrifosfaţi (ATP, GTP, CTP, UTP)
Sinteza are loc în direcţia 5’→3’
Este asimetrică – copierea catenei necodificătoare
Este incompletă –are loc copierea doar a unei porţiuni de ADN (transcripton: promotor, operator, GS, terminator)
Forţa motrice a procesului e hidroliza PP
Enzima - ARN polimeraza
")
Слайд 76ARN polimeraza
este o holoenzimă
la procariote - este oligomer din 5
α subunităţile – centre catalitice;
β - fixează substratul;
β 1 – se leagă de ADN,
δ - are rol în recunoaşterea secvenţelor matriţei numit promotor, unde aderă enzima
la eucariote:
RNApI sintetizeaza RNA ribozomal (28S si 18S)
RNApII sintetizeaza RNAm•
RNApIII sintetizeaza RNAt, RNAr 5S şi molecule mai mici
3. Nu necesită prezenţa primerului
4. Nu posedă funcţie nucleazică, doar polimerazică
5. ADN polimeraza conţine Zn2+ şi necesită prezenţa în mediu a ionilir de Mg2+, Mn2+

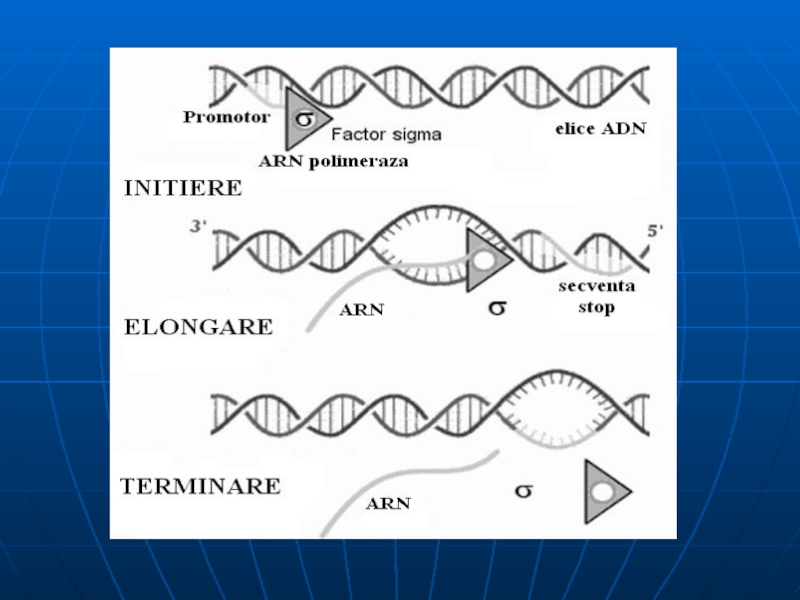
Слайд 77Etapele transcripţiei
Sinteza decurge în 3 etape: iniţierea, elongarea, terminarea.
Iniţierea – începe
Pe P deosebim 2 locusuri: de recunoaştere (depistat cu ajutorul sigmei) şi locusul de legare laxă a ARN polimerazei. Locusul de recunoaştere e situat la o distanţă de 25 nucleotide de locusul de legare şi 10 nucleotide de la punctul de iniţiere (+1)
Toţi P bacterieni au 2 secvenţe consens situate pe catena codificătoare:
Caseta Pribnow (-10) 5’ TATAAT 3’ –responsabilă de iniţierea denaturării locale a ADN
Caseta -35 5’ TTGACA 3’ - la care are loc asocierea primară a ARN polimerazei
δ subunitatea recunoaşte caseta -35 şi se leagă la ea. E. alunecă de-a lungul ADN şi în jurul casetei -10 (Pribnow) deschide dublul helix – formând complexul deschis de iniţiere
ARN p catalizează formarea primei leg. fosfodiesterice între nucleotidul +1 şi +2 -δ disociază, iar E-cor continuă sinteza.


Слайд 79Etapele transcripţiei
Sinteza decurge în 3 etape: iniţierea, elongarea, terminarea.
Iniţierea – începe
Toţi P bacterieni au 2 secvenţe consens situate pe catena codificătoare:
Caseta Pribnow 5’ TATAAT 3’
Caseta 35 5’ TTGACA 3’
P la eucariote:
GC casete GGGCG
CAAT casete CCAAT
Caseta Hogness –TATAT/AT
Primele 2 sunt responsabile de frecvenţa transcripţiei (când începe), a 3 - - implicată în iniţiere – semnal (unde). Toate sunt responsabile de exacitatea iniţierii
Pe P deosebim 2 locusuri: de recunoaştere (depistat cu ajutorul sigmei) şi locusul de legare laxă a ARN polimerazei. Locusul de recunoaştere e situat la o distanţă de 25 nucleotide de locusul de legare şi 10 nucleotide de la punctul de iniţiere (+1)

Слайд 80
Sigma subunitatea găseşte punctul de iniţiere şi:
Activează identificarea secvenţelor de RNA
Ia parte la desfacerea dublului helix de ADN
Ia parte la formarea primei legături fosfosiesterice
Astfel complexul de iniţiere este format, sigma subunitatea e disociată de la holoenzimă şi ia parte la iniţierea unui alt ciclu de transcriere.

Слайд 81Elongarea şi Terminarea
Elongarea - alunecarea ARN polimerazei pe matrţa de ADN
Terminarea – RNAp recunoaşte secvenţele nucleotidice specifice de pe ADN, ce conţin un număr mare de G, C . Proteina ρ - se asociază la E şi se mişcă împreună cu ea, însă la identificarea semnalelor de terminare coboară de pe matriţă şi încetineşte acţiunea E,producând transcriptul cu folosirea energiei – ATP

Слайд 83Transcripţia la eucariote
RNA p alcătuită din 9-11 subunităţi
Folosesc mai multe tipuri
ARN polimeraza I – ARNr (18S, 28S, 5,8S; 45S) –în nucleoli;
ARN polimeraza II – ARNm;
ARN polimeraza III –ARNt şi ARNr (5S)
ARN polimeraza IV (mitocondrială)- toate tipurile de ARN mitocondrial
P la eucariote:
GC casete GGGCG (-90)
CAAT casete – 5-GGCCAATCT -3 (-75)
Caseta Hogness –TATAT/AT (-35)
Primele 2 sunt responsabile de frecvenţa transcripţiei (când începe), a 3 - - implicată în iniţiere – semnal (unde). Toate sunt responsabile de exacitatea iniţierii
Secvenţele alcătuite din 10-20 nucleotide “enhancers” şi “silencers” – cresc şi scad respectiv V transcrierii; pot fi situate la distanţe mari de gena transcrisă.

Слайд 84Procesingul
Toţi precursorii de ARN în nucleu trec etapa de maturizare posttranscripţională.
Procesingul înclude:
1. Modificarea fragmentelor terminale 5’ şi 3’ ale ARN:
a. “Cap”-area: la capătul 5’ -este adiţionată guanozina metilată (5’- 5’ trifosfat - protejarea mARN de atacul 5’-exonucleazelor şi pentru recunoaşterea de către ribosomi ca semnal de iniţiere);
b. la capătul 3’ – se adaugă o secvenţă mare de poli A (200 A -coadă). Ea serveşte la exportul moleculelor de ARN din nucleu în citoplasmă.


Слайд 86
2.Splisingul - excizia intronilor şi sudarea exonilor. Aşa numitul splising are
a.ARN nuclear (ARN U) identifică secvenţele de baze la joncţiunea intron – exon,
b. se fixează complementar la ele, buclează intronul, astfel apropiind capetele exonilor.
c. Are loc scindarea legăturilor fosfodiesterice dintre exoni şi introni, capetele exonilor sunt juxtapuse, apoi sudate de RNA ligazele



Слайд 90Transcripţia inversă
sinteza ADN pe catena de ARN
Matriţa – ARN
Substrat –
Enzima – revers transcriptaza
Caracteristic viruşilor oncogeni
Mecanismul:
a. Revers transcriptaza sintetizează pe ARN viral catena de ADN- hibrid: ADN_ARN
b. Scindarea ARN viral de o nuclează
c. Autoreplicarea ADN – cu formarea unui duplex de ADN


Слайд 92Obiectivele:
Codul genetic. Proprietăţile.
Ribozomii - sediul sintezei proteinelor, structura lor.
Procesul de translare
Reglarea biosintezei proteinelor. Inducţia şi represia enzimelor.
Inhibitorii sintezei proteice.
Polimorfismul proteinelor (variantele hemoglobinei, enzimelor, grupelor sanguine).
Bolile ereditare şi diagnosticul lor biochimic.
. Modificările posttranslaţionare")
Слайд 93Codul genetic
Informaţia genetică referitor la biosinteza proteinelor se transmite cu ajutorul

Слайд 94Proprietăţile codului genetic
Este triplet -64 codoni: 3 nonsens:UAG; UGA; UAA;
este degenerat - unui AA poate să-i corespunda mai mulţi codoni (Ex. Arg, Leu, Ser - codificate de 6 codoni; Met- şi Trp - un codon). Codonii unui aminoacid sint sinonime. Specificitatea codonului e determinată de primele două litere. Degenerarea se referă la nivelul nucleotidului 3 din codon sau 1 din anticodon care oscileaza.
nu este ambiguu- acelaşi triplet nu semnifică 2 AA diferiţi
Are o structură liniară (colinear) – o concordanţă liniară între genă şi proteina codificătoare
Nu se suprapune (excepţie- viruşii)

Слайд 95
Este universal – toate veţuitoarele utilizează acelaşi mecanism de traducere (abatere
nu are virgule, semne de punctuatie - ce ar indica începutul şi sfîrşitul fiecarui codon.
AUG - este codonul de initiere
UAG, UAA,UGA - codoni stop (non sens)
Toţi codonii cu U (în pozitia 2) codifica AA hidrofobi
codonii cu A în pozitia -2 codifică AA polari
Uracilul în poziţia 1 prezintă codonul nonsens
dacă în anticodon în directia (5'->3') prima bază nucleotidică e: a) Citozina sau Adenina, el va citi un singur codon; b) Uracilul sau Guanina el va citi 2 codoni; c) inozina - respectiv va citi 3 codoni

Слайд 96Ribozomii
Reprezintă sediul de traducere a ARNm şi sinteza proteinelor.
Structura- complexe ribonucleoproteice
Structura ribozomilor procariotici:
subunitatea 30 S – conţine ARNr 16S şi 21 proteine.
subunitatea 50S – ARN r 5S, 23S şi 31 proteine.
Sinteza ARNr şi formarea subunităţilor are loc în citoplasmă.
Structura ribosomilor eucariotici:
subunitatea 40S – ARNr 18S şi 33 proteine.
subunitatea 60S – ARN r 5S, 5,8S, 28S şi 49 proteine. ARNr – se formează în nucleol.
Ribozomul va avea constanta de sedimentare 70S la procariote si 80S la eucariote.
S – este coeficientul de sedimentare Svedberg, care depinde de forma, densitatea şi dimensiunea particulelor.

Слайд 97Centrele catalitice ale ribosomilor
Situsul A - aminoacil – responsabil de unirea
Situsul P – peptidil – găzduieşte ARNt legat de un lanţ polipeptidic deja sintetizat
Situsul E – e responsabil de eliminarea ARNt
Procesul de sinteză proteică poate fi schiţat sumar prin interacţiunea celor 3 tipuri de ARN - informaţia din ARNm este citită in ribozom si transpusă în proteine, AA necesari fiind aduşi de ARNt.
În starea complet nedisociată ribozomii sunt activi.
Deplasarea lberă a ribozomilor în diferite sectoare ale celulei, sau combinarea lor în diferite locuri cu membranele reticulului endoplasmatic oferă posibilitatea de asamblare a proteinei în celulă.

Слайд 98
Mai mulţi ribozomi pot citi simultan acelaş ARN mesager pe care

Слайд 99Translaţia
Translaţia sau biosinteza proteinelor propriu zisă.
Bazele moleculare ale translaţiei:
m-RNA ca
aminoacil – tRNA;
ribozomii ca maşini moleculare pentru unirea succesivă a AA în catena polipeptidică conform programului mRNA;
GTP ca sursă de energie;
“factorii” proteici care vin în ajutor în diferite etape ale asamblării proteinei în ribozomi;
unii ioni ca cofactori (Mg 2+, K+).

Слайд 100Etapele
se realizeaza in 5 etape:
Activarea AA.
Iniţierea lanţului polipeptidic.
Elongarea lanţului polipeptidic.
Terminarea
Prelucrări post traducere ale proteinei sintetizate.

Слайд 101Activarea AA
are loc în citozol
Sunt necesare:
AA (procariote – Nformil-Met; la eucariote
ARNt
ATP, Mg, K
E – aminoacil ARNt sintetaza (există nu mai puţin de 20 – indentificate 32 de tipuri de ARNt):
Posedă 4 centre: pentru AA, ATP, ARNt, H2O
Specificitatea E e determinată de structura ARNt
Fidelitatea e asigurată de capacitatea de autocontrol
Conţine grupări libere sulfhidrilice
sunt ligazele, care au o specificitate absolută.
ARNtATP, Mg, KE")
Слайд 102
Se desfăşoară în două etape:
1. NH2-CH-COOH NH2-CH-CO –O-AMP
I I
R R Aminoacil Adenilat
2. NH2-CH-CO –O-AMP+ARNt NH2-CH-CO –O-RNAt +AMP
I I
R R
Aminoacil RNAt
I etapă - AA reacţionează cu ATP rezultând aminoacil-AMP. PP eliberat este hidrolizat şi in acest fel reacţia ireversibilă.
II etapă - complexul cedează AA moleculei de ARNt, specifică acelui AA
Activarea AA
Aminoiacil ARNt
sintetaza


Слайд 104Activarea AA
Esenţa procesului de activare este fixarea AA la ARNt propriu
Enzima poate recunoaste daca un AA gresit s-a fixat pe ARNt, situatie in care il elimina si il inlocuieste cu AA corespunzator deoarece prezinta si un locus hidrolitic.

Слайд 105Activarea AA
Activarea AA consumă 2 legături macroergice
ARNt pe calea difuziunii

Слайд 107Translaţia propriu zisă
Citirea ARNm se face în direcţia 5‘- 3' iar
se desting trei etape:
Iniţierea
Elongarea
terminarea.

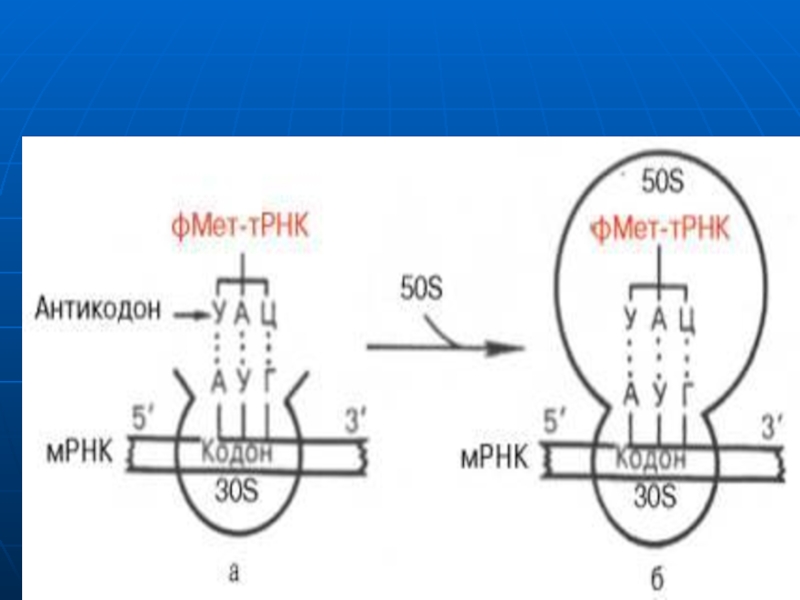
Слайд 108Iniţierea
Necesar:
ARNm (AUG)
Ribosomul cu subunităţile disociate
ARNt f-met (Met)
GTP, Mg
IF1, IF2, IF3
Scopul: formarea
Ribosomul cu subunităţile disociateARNt f-met (Met)GTP, MgIF1, IF2, IF3Scopul: formarea complexului de iniţiere")
Слайд 109Formarea complexului de iniţiere:
Subunitatea mică leagă IF3 şi previne reasocierea ribosomilor
La
La complex adiţionează IF1, mai apoi IF2 legat de GTP şi formil Met-ARNt
Îndată cum are loc fixarea anticodonului fMet-ARNt cu codonul AUG (ARNm), are loc hidroliza GTP, eliberarea FI şi unirea subunităţilor
Formil Met-ARNt –e fixată în centrul P
30S
FI3
P 50S A
AUGCCGGUAAUAAAGGGCCCAGG
FI1
FMET
GTP
FI2
GDP Pi
UAC
")
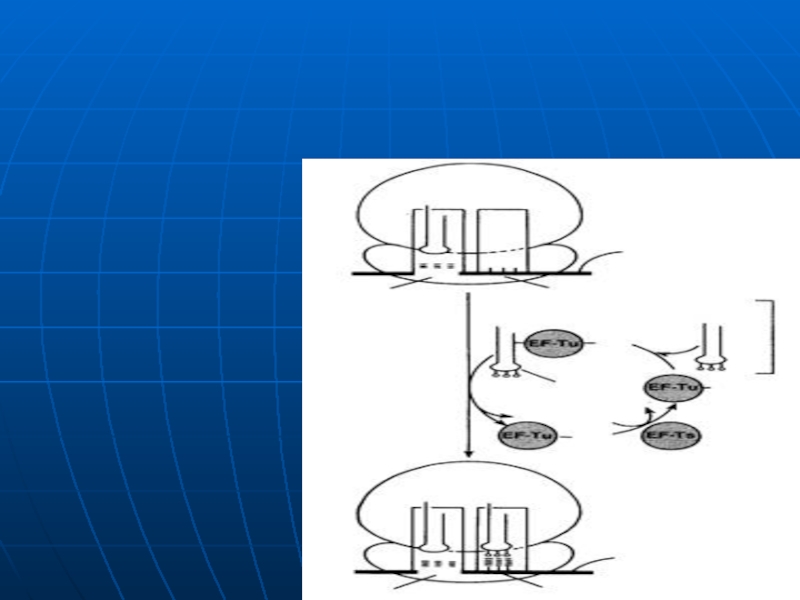
Слайд 111Elongarea
Necesar:
ARNm cu următorul codon
ARNt cu următorul AA
GTP
FE: Tu, Ts, G
Elongarea translaţiei
Legarea aminoacil – ARNt;
Transpeptidarea- formarea legăturii peptidice,
Translocarea (deplasarea ARNm cu un codon).

Слайд 1121. Adaptarea (legarea)AA
are loc după principiul codon- anticodon în centrul A
a.
b. AA se fixează în centrul A. Simultan are loc hidroliza GTP în GDP şi P
Tu-GDP+GTP-Ts------Tu-GTP
AAare loc după principiul codon- anticodon în centrul Aa. Aminoacil-ARNt se fixează cu")
Слайд 1142. Transpeptidarea
este formarea legăturii peptidice între doi aminoacizi.
AA din centrul P
Se formează dipeptida
În centrul P rămîne ARNt liber

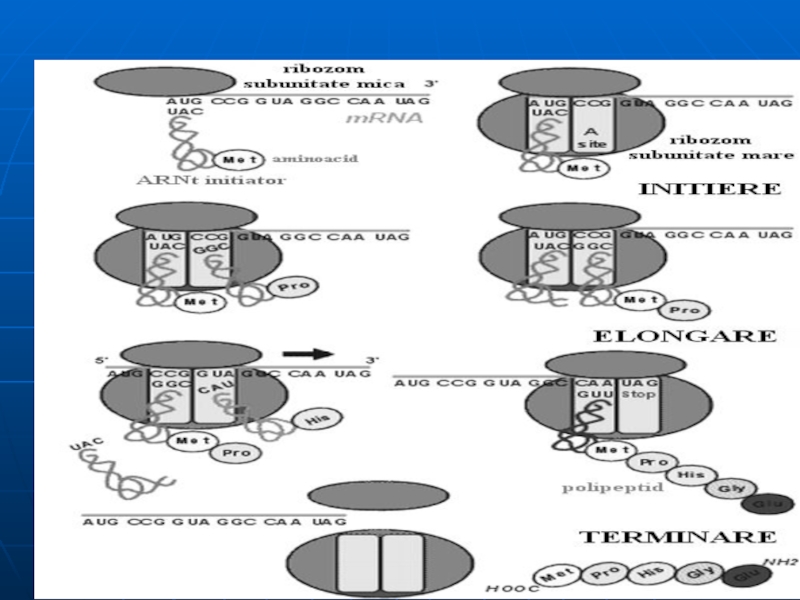
Слайд 1163. Translocarea
deplasarea ARNm cu un triplet în direcţia 5‘- 3' .
Dipeptida
ARNt din P părăseşte ribosomul

Слайд 117Elongarea
Decurge în 3 etape :1 fixarea noului Aminoacil ARNt
complexul: aminoacil-ARNt,
se fixează pe situsul A, după ce are loc hidroliza GTP la GDP care se eliberează împreună cu FE-T.
2. formarea legăturii peptidice.
Enzima peptidil-transferaza catalizează formarea legăturii peptidice între doi AA din situsul A şi P.Peptida rămîne ataşată de RNAt de pe situsul A.
translocaţia
ribozomul se deplasează la următorul codon de pe ARNm şi peptidilARNt trece de pe situsul A pe P
această etapă necesită factorul de elongare G (FE-G) şi GTP (necesar pentru realizarea modificărilor conformaţionale care deplasează ribozomul).
30S
P 50S A
FMET
UAC
AUGCCGGUAAUAAAGGGC
FE-T
GTP
GDP P
E-PT
FE-G
GTP
FE-T
GTP

Слайд 118Terminarea
are loc cînd sunt întîlniţi codonii UAA, UGA, UAG şi factorii
Nici un tRNA nu se poate lega cu codonii de terminare.
Factorii de terminare:
eliberează lanţul polipeptidic
Elimină ARNt din centrul P
Disocierea ribosomului în subunităţile respective

Слайд 119
La formarea unei legături peptidice se consumă patru legături macroergice:
2
2 în elongare: legare şi translocare - GTP.

Слайд 120Prelucrările posttraducere
Modificarea capătului N- şi C-terminal; capătul N se acetilează;
Îndepărtarea secvenţei
modificarea unor AA:
hidroxilarea enzimatică a Pro, Lyz – obţinerea hidroxiprolinei, hidroxilizinei .
Metilarea (Lyz în muşchi)
Carboxilarea Glu - γ-carboxil-glutamatului (protrombină)
oxidarea reziduuriilor de Cis - cistinei;
iodurarea reziduurilor de Tir ale tireoglobulinei.
ataşarea unor gr. funcţionale: fosfat, glicozil, metalelor pentru formarea fosfoproteinelor,glicoproteinelor, metaloproteinelor ş.a.
Formarea punţilor disulfurice
Proteina se autoasamblează – formând conformaţia nativă – structura tridimensională

Слайд 121Inhibitori ai sintezei proteinei
la nivelul replicării:
Mitomicina –împiedica separarea catenelor de
Acid nalidixic –inhiba ADN giraza
la nivelul transcriptiei:
- Actinomicina D - se fixeaza pe ADN
– Rifampicina - inhiba ARN polimeraza

Слайд 122Inhibitorii sintezei proteinei
la nivelul translatiei:
Streptomicina –inhiba legarea ARNt initiator la
Cloramfenicol -inhiba peptidil transferaza
Tetraciclina - inhiba legarea ARNt la ribozomi
Eritromicina,Azitromicina – blocheaza subunitatea 50S
Puromicina – blocheaza elongarea inhibând competitiv ARNt
Streptomicina - interferă cu legarea formil-Met-ARNt la locul de iniţiere.
Neomicina, Kanamicină - erori în reproducerea codului genetic
Toxina difterică - inhibă translocaza

Слайд 123Reglarea sintezei proteinelor
Sinteza proteinelor nu e constantă – ea trebuie să
Celulel dispun de 3 tipuri de enzime :
Constitutive - se sintetizează în celulă cu o viteză constantă.
Inductible - sunt E a căror concentraţie depinde de prezenţa sau absenţa din mediu a unui compus denumit inductor. Sunt implicate în căile catabolice,
Represible - sunt E a căror concentraţie depinde de prezenţa sau absenţa din mediu a unui compus denumit corepresor. Sunt implicate în căile anabolice.
În mod normal cantitatea de E inductibile în celule este foarte mică,dar ea poate creşte atunci cînd apare necesitatea utilizării substratului E respective ( S care se comportă ca inductor).

Слайд 124Teoria lac-operonului
Schema reglării biosintezei proteinei la procariote a fost descrisă în
Modelul e bazat pe studiul reglării mtabolismului lactozei în Escheria coli.
Exprimarea GS (conţin informaţia cu privire la biosinteza E impicate în utilizarea lactozei:β-galactozidaza, permeaza şi transacetilaza –1,2,3) e controlată de un fragment de ADN denumit genă reglatoare (GR) - codifică represorul (R). R se leagă de un fragment de ADN denumit operator (O).
Legarea R la O blochează accesul ARN – polimerazei la promotor avînd ca rezultat suprimarea transcrierii GS.

Слайд 125
Ce se întâmplă dacă bacteria dispune simultan de glucoză şi lactoză?
Bacteria creşte pe seama glucozei - şi numai atunci când c% acesteea devine minimă începe să utilizeze lactoza. Metabolizarea simultană a glucozei şi lactozei sunt excluse.
Cum se comută activitatea bacteriei pe utilizarea lactozei când c% glucozei scade?


Слайд 127Reglarea sintezei proteinei prin inducţie
În prezenţa lactozei:
Inductorul (în acest caz lactoza)
 se leagă specific la")

Слайд 129
În lipsa glucozei se măreşte c% AMPc – ce reprezintă semnalul
Ca urmare, AMPc se leagă de o proteină receptoare specifică (proteina activatoare a genei catabolice – CAP) - formează complex (CAP- AMPc), apt să se lege de promotor (p-locus)
Acest proces favorizează pătrunderea ARNp în locusul de reglare. Dacă lactoza este prezentă în mediu, operatorul este liber şi ARNp efectuează transcrierea genelor lac.
CAP dispune de 2 centre: pentru AMPc şi pentru ADN

Слайд 130Reglarea lac-operonului într-un mediu ce conţine glucoză
Cu cât c% glucozei e

Слайд 131Ilustrarea mecanismului de reglare a sintezei proteinei prin represie
Teoria operonului explică
Ex: sinteza His: la c% mari de His (corepresor) – se leagă de R, modificându-i conformaţia – activându-l – în rezultat favorizează legarea R la O.
His – produs final, CoR- sistează transcrierea genelor ce codifică E implicate în propria sa sinteză


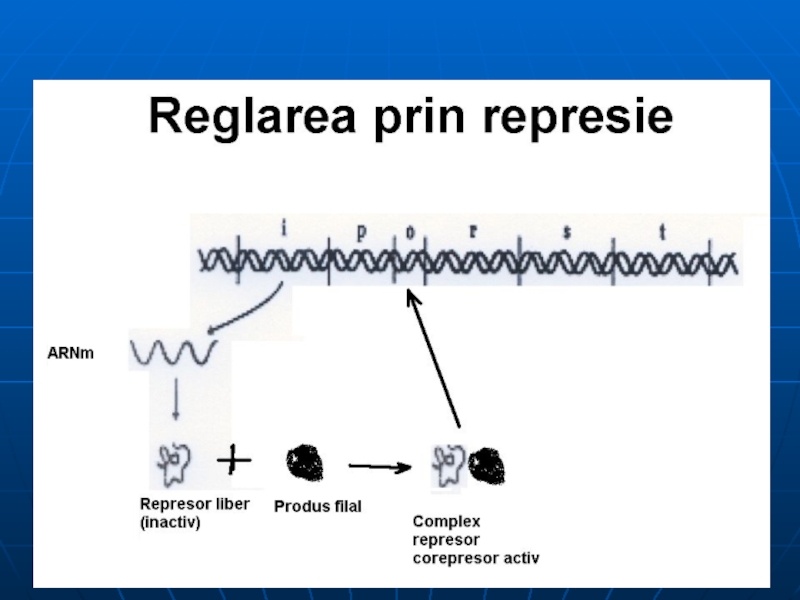
Слайд 134REZUMĂM:
GR controlează exprimarea anumitor GS prin intermediul unei proteine – R
Ra
E inductibile – Ra – nu are loc transcrierea. Când în mediul apare I – R se inactivează – are loc sinteza ARNm- proteinei
E represibile – R este inactiv – are loc transcripţia şi translaţia. Când în mediu se acumulează produsul final al căii anabolice (CoR)- R se activează, formarea complexului R-CoR – şi sistarea transcripţiei şi translaţiei.

Слайд 135Reglarea sintezei la eucariote
Atât la nivelul transcripţiei cât şi la nivelul
Reglarea hormonală (cortizol- sinteza E gluconeogenezei; estrogenii, androgenii, vitamina D – sinteză de proteine specifice)
Reglarea exspresiei genetice prin moleculele proteice legate de ADN (histonele) – sinteza ARN pe ADN e inhibată prin adaosul de histone
Reglarea proteinei la nivelul translaţiei – e posibilă prin acţiunea factorilor proteici, care contribuie iniţierea, elongarea, terminarea.

Слайд 136Ingineria genetică
ştiinta, preocupată de crearea noilor fenotipuri prin transplantarea genei unui
Se obtin molecule hibride (himerice)
În linii marl procedura include etapele:
1. Căpătarea genei
2. Căpătarea ADN-ului recombinat
3.Clonarea ADN-ului recombinat
Căpătarea genei:Stiind structura primară a proteinei în laborator se poate obline gena respectivă (se oblin gene pînă la 250 codoane)- mai greu e obtinerea genei din genomul celulei (genele se despart prin introni)- mai uşor e căpătarea genelor din virusuri cu enzima revertaza.

Слайд 137Căpătarea ADN-ului recombinat-
gena necesară se întroduce în celulă pentru a se
De regulă se foloseşte E Coli, ce contine un cromozom şi plasmide, ce plutesc în citozol (plasmida este de 1000 ori mai mica decît cromozomul). Plasmidele se replica independent de replicarea materialului genetic. Unele plasmide se pot include în cromozom şi apoi din nou să-l părăsească. Plasmidele pot trece dintr-o celulă în alta în procesul deconjugare. Plasmidele se separă din E.Coli şi li se înlătură o parte de ADN inelar cu ajutorulenzimelor restrictaze, care recunosc şi taiediferite sectoare. Folosind restrictaze diferite se poate de tăiat ADN în locusurile necesare. Inrezultat se formează capete lipicioase (sectoaremonocatenare, capabile de a uni nucleotide complimentare. La fel se procedează şi cu gena,care trebuie întrodusă (se formează capete lipicioase complimentare capetelor plasmidei). Dacă se amestecă gena şi plasmida ele se vor uni cu capetele lipicioase. Enzima ligaza va uni capetele şi se va căpăta molecula ADN inelara, care contine gena menită pentru transplantare.

Слайд 1383.Clonarea
ADN-ului recombinat- obtinerea cantităţilor dorite de proteină codificată de gena eucariotă

Слайд 139
Ranadamentul sintezei bacteriene este impresionabil: Ex- 100 celule E.Coli produc prin clonare
Diversitatea formelor în Iimita uneia şi aceaşi specie se datoreşte mutaţiilor şi într-o măsura mai mare recombinării genetice.

Слайд 140Mutaţiile.
Modificările genomului organismului, care se păstrează şi se transmit prin ereditate
se transmit apoi de la o generaţie la alta.
Modificările pot interesa o pereche de baze (mutatii punctiforme) sau un grup de baze pe una sau pe ambele catene ale unei molecule de DNA.

Слайд 141
Mutatiile punctiforme: pot decurge prin:
l. substitutie (misens mutatii, unde deosebim 2
a. Tranzitie - o BA purinică este înlocuită tot cu una purincă, una pirimidinică -tot cu una pirimidinică.
b.Transversie - o pereche de baze purinice este înlocuită cu una pirimidinică sau invers.
2. Inserţie - acest mecanism constă în întroducerea unei perechi de baze suplimentare în catena de DNA.
3. Deleţia constă în excluderea unei perechi de baze în aşa mod ca ea nu mai poate fî complementară şi la replicare apare "golul" în ambele catene. Unele modificări în secventa nucleotidică pot duce la formarea codonului sinonim şi succesiunea aminoacizilor nu se va schimba (mutatii benigne).
La afectarea segmentelor mari de genă apar mutatii întinse. In dependentă de consecinţele modificărilor deosebim mutaţie benignă, neutră, nocivă.
Agenţii mutageni pot provoca mutaţiile spontane cît şi mutaţiile induse.
:")
Слайд 142
şi două uşoare (L-214AA),
constituiti din:- 2 lanturi polipeptidice
grele identice (H446 AA)
fiecare dintre ele
contin cite o porţiune:
constantă „C"
varîabilă „V".
şi una
, constituiti din:- 2 lanturi")
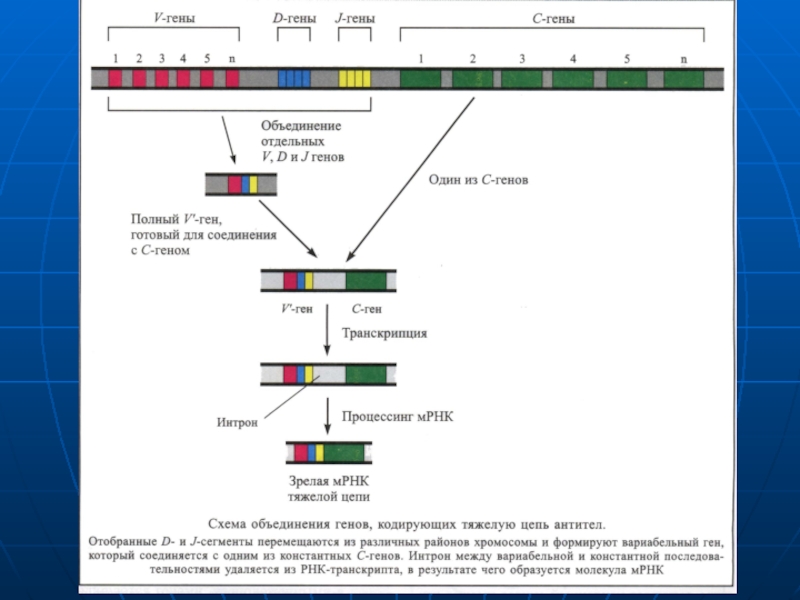
Слайд 143Secvenţa de AA în porţiunea variabilă este diferită pentru fiecare anticorp.
Genele ce corespund porţiunilor „V" şi „C“ ale unui anumit tip de lanţ uşor sunt foarte apropiate în ADN al imunocitelor care produc acest tip de lanţ uşor, dar se găsesc departe una de alta în ADN al celulelor ce produc alte tipuride anticorpi.
De aici reiese, că în imunocit se selectează un anumit segment de ADN, ce codifică porţiunea variabilă a unui anumit lant uşor, care se transferă prin transpoziţie în vecinătatea secvenţei codificatoare a porţiunii constante a lantului uşor. Deci ADN ce codifică sectoarele „V" ale lanţurilor „H" şi "L" constă din cîteva gene de tip diferit care-şi pot schimba locurile proprii şi asocia cu formarea imenselor combinaţii.

Слайд 144Sinteza Anticorpilor
Fiecare dintre milioanele de anticorpi produşi leagă unul dintre milioanele

Слайд 145
Gradul de diversificare în obţinerea lanţurilor „H" şi „L" este crescut
Porţiunea variabilă (V) constituită din - 400 de gene.
Porţiunea de diversitate (D) ce cuprinde -12 gene
Portiunea de articulare sau jonciune ţ(J) - 4 gene
Asamblarea acestor gene în diferite combinaţii permite construirea a 20000 de sectoare V - fapt ce asigură extrema varietate a anticorpilor.