- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Введение в нейрофизиологию презентация
Содержание
- 1. Введение в нейрофизиологию
- 2. План лекции Введение История развития нейрофизиологии Основные
- 3. РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА ДЛЯ САМОСТОЯТЕЛЬНОЙ ПОДГОТОВКИ
- 4. Крылова Н.В., Искренко И.А. Мозг и проводящие
- 5. Достижения человечества существуют благодаря разуму, вместилищем которого
- 6. Введение Нейрофизиология относится к разделу физиологии, который
- 7. Разделы нейрофизиологии Общая нейрофизиология - изучает закономерности
- 8. История открытия и развитие представлений о биопотенциалах головного мозга
- 9. История развития нейрофизиологии Зависимость сознания от мозга
- 10. Первые исследования около 620-550 до н.э. –
- 11. История электрофизиологии началась с изучения влияния электричества
- 12. История развития нейрофизиологии Первые представления о рефлекторном
- 13. Начало электрофизиологии обычно связывают со знаменитыми опытами
- 15. Карло Маттеучи (Carlo Matteucci 1811 – 1868)
- 17. Опыт по электрофизиологии нервов и мышц
- 18. В 1875 году английский хирург и физиолог
- 19. Василий Яковлевич Данилевский В 1875 г.
- 20. Иван Михайлович Сеченов 1864 г. «Рефлексы
- 21. Иван Михайлович Сеченов 1829 – 1905
- 22. Он обосновал возможность распространения принципа рефлекса как
- 23. В 1884 году Н.Е. Введенский в работе
- 24. Б.Ф. Вериго В 1889 г. В журнале
- 25. 1890 год - работа А. Бека, посвященная
- 26. 1898 год - В.Е. Ларионов производит исследования
- 27. 1900 год – работа С.А. Тривуса, который
- 28. 1912 год - обширное исследование П.Ю. Кауфмана,
- 29. Первая запись биотоков головного мозга человека получена
- 30. Нейрофизиологические методы: Внутриклеточная регистрация активности нервных клеток
- 32. Регистрация импульсной активности нервных клеток Фотографии нейронов, окрашенных флуоресцентным красителем Подведение электрода к телу нейрона
- 33. Два подхода к изучению мозга Гемодинамика (метаболизм) – ПЭТ, ЯМРИ Электрофизиология – ЭЭГ, МЭГ
- 35. Основные ритмы ЭЭГ Альфа-ритм, 8-13 Гц 50-100
- 38. Современные методы исследования мозга человека, позволяющие локализовать функции Компьютерная томография Ядерно-магнитный резонанс Магнитоэнцефалография Позитронно-эмиссионная томография
- 41. Компьютерная томография
- 42. Компьютерная томограмма
- 43. Позитронно-эмиссионная томография
- 45. Результаты ПЭТ-сканирования ПET - изображение мозга, показывающее зрительное и слуховое распознавание
- 46. Ядерно-магнитный резонанс
- 49. Ядерная магнитно-резонансная интроскопия (ЯМРИ, MRI)
- 50. Послойное сканирование ЯМРИ
- 51. Возникновение современной нейронауки На современном этапе функции
- 52. Задачи нейронауки Объяснить поведение человека в
- 54. Молекулярный уровень рассматриваются вопросы морфологии и физиологические
- 55. Клеточный уровень Оценка механизмов восприятия сигналов дендритами,
- 56. Системный уровень Использование анатомических и физиологических знаний
- 57. Когнитивный уровень Оценка психологических функций с точки зрения формирования нервной системы.
- 58. Основные процессы в нервной системе Трансдукция.
- 59. Основные процессы в нервной системе Моделирование.
- 60. Виды модуляции 1. Активация (возбуждение). Повышение
- 61. Виды модуляции 3. Пластическая перестройка нервной структуры.
- 62. Виды модуляции 4. Активация исполнительного органа для
- 65. Нейрон и его элементы
- 67. Нейрон – структурно-функциональная единица центральной-нервной системы. Структурно-функциональная организация нервной системы
- 68. Тело (сома) нейрона - центральная часть
- 69. По отношению к отросткам тело клетки выполняют
- 70. Аксон – одиночный, обычно длинный выходной
- 72. Строение нейрона Дендриты – древовидно-ветвящиеся
- 73. Строение нейрона На дендритах имеются микроскопических
- 74. Виды нейронов А – униполярные: имеют один
- 75. Виды нейронов В –биполярные клетки: имеют
- 76. Виды нейронов По форме: Пирамидные, Веретенообразные, Звездчатые, Корзинчатые.
- 77. Виды нейронов По локализации: Центральные -
- 78. Виды нейронов по выполняемой функции Афферентные (чувствительные)
- 79. Виды нейронов по выполняемой функции Вставочные нейроны
- 80. Тела нейронов образуют серое вещество головного мозга:
- 81. Глиальные клетки Глиальные клетки в совокупности называют
- 82. Анатомическое расположение клетки нейроглии в головном мозге; шванновские клетки в периферической нервной системе.
- 83. Функции нейроглии Опорная – поддерживает нервные
- 84. Функции нейроглии Участие в регенерации. В
- 85. Виды глиальных клеток Эпиндемоциты - выстилают полости
- 86. Астроциты Функции Гомеостатическая (поддержание ионного и химического
- 87. Олигодендроциты Находятся преимущественно в белом веществе мозга, где они образуют миелин вокруг крупных аксонов.
- 88. Радиальная глия Участвует в миграции нейронов во время внутриутробного развития плода
- 89. Тело нейрона покрыто слоистой мембраной, которая
- 91. Строение нейрона Белки мембраны: Интегральные – ионные каналы Поверхностные - рецепторы
- 92. Структуры, поддерживающие гомеостаз нейрона: Мембрана – отделяет
- 93. Схема ионного канала
- 94. Функция канала
- 95. Основные функции мембраны Образует полупроницаемый, избирательный барьер
- 96. Ионные каналы 1. Пассивные (для калия) 2. Управляемые: потенциалзависимые; хемозависимые; механические
- 98. Мембрана называется полупроницаемой, потому что: Мембрана ограничивает
- 99. Благодаря этим свойствам мембрана создает и поддерживает
- 100. В покое мембрана более проницаема для ионов К+
- 101. Концентрационный градиент Различная концентрация ионов является движущей
- 102. Возникает и поддерживается МП МП = ПП
- 103. ГЕНЕРАЦИЯ ВОЗБУЖДЕНИЯ В НЕРВНОЙ СИСТЕМЕ
- 104. Основной принцип функционирования центральной нервной системы –
- 105. Мембранный потенциал это разность электрических потенциалов (напряжение) между внутренней и наружной поверхностью мембраны.
- 106. Мембрана живой клетки поляризована — её внутренняя поверхность
- 108. Потенциал покоя нейрона Если ввести внутрь живой
- 109. Потенциал покоя нейрона Снаружи — преобладают ионы натрия, кальция
- 110. Под воздействием стимула происходит открытие натриевых каналов.
- 111. Потенциал действия нейрона Исходную концентрацию ионов восстанавливает натрий-калиевый
- 112. Как распространяется возбуждение (потенциал действия) в нервной
- 113. Согласно этой теории биоэлектрических потенциалы обусловлены неодинаковой различной проницаемостью для них поверхностной мембраны
- 118. Потенциал действия (ПД) или Нервный импульс
- 119. Потенциал действия нейрона
- 120. ПД и фазы
- 121. Распространение ПД по волокну
- 122. В немиелинизированных нервных волокнах возбуждение распространяется посредством
- 123. В миелинизированных волокнах, где поперечное сопротивление увеличивается
- 125. Потенциал действия Возникновению потенциала действия предшествуют активные
- 127. В практической медицине используют эти фундаментальные знания
- 128. Закономерности проведения возбуждения по нервным волокнам Двустороннее
- 129. Контрольные вопросы
- 131. Кто впервые определил наличие
- 132. Чему равен потенциал покоя
- 133. Благодарю за внимание!

Слайд 2План лекции
Введение
История развития нейрофизиологии
Основные процессы в нервной системе
Нейрон – как структурно-функциональная
Заключение


Слайд 4Крылова Н.В., Искренко И.А. Мозг и проводящие пути: Анатомия человека в
Коган А.Б. Функциональная организация нейронных механизмов мозга. – Л., 1979.
Котляр Б.И., Шульговский В.А. Физиология ЦНС. – М., 1985.

Слайд 5Достижения человечества существуют благодаря разуму, вместилищем которого является головной мозг.
Все продукты
Решения: эффективное использование новых методов исследования головного мозга.
Мозг – основа существования человека и общества

Слайд 6Введение
Нейрофизиология относится к разделу физиологии, который изучает функции нервной системы, в
Нейрофизиология – это наука о жизнедеятельности головного мозга, о его взаимодействии с внешней средой и динамике процессов, протекающих на уровне мембраны нервных клеток, синапсов и нервных тканей.

Слайд 7Разделы нейрофизиологии
Общая нейрофизиология - изучает закономерности функционирования нервной системы на разных
Возрастная нейрофизиология –это раздел, посвященный изучению возрастных и индивидуальных особенностей мозгового обеспечения высших нервных и психических процессов.
Клиническая нейрофизиология изучает особенности функционирования мозга при патологических процессах.


Слайд 9История развития нейрофизиологии
Зависимость сознания от мозга утверждал еще отец медицины -
Ученики Гиппократа, следуя своему учителю, рассматривали мозг как центр всех центров и главный орган ума.

Слайд 10Первые исследования
около 620-550 до н.э. – Фалес Милетский - считается первооткрывателем
около 1600 года - Вильям Гильберт (William Gilbert) начал изучать электрические свойства различных веществ, а Отто ван Герик (Otto von Guericke 1602-1686) изобрел электрофорную машину для создания электрических полей.

Слайд 11 История электрофизиологии началась с изучения влияния электричества на организм. В 18
Раздражение вызывало своеобразное субъективное ощущение и некоторые воздействия на организм, например, непроизвольные и судорожные сокращения и подергивание мышц. Эти влияния привлекли внимание врачей и физиологов.

Слайд 12История развития нейрофизиологии
Первые представления о рефлекторном принципе действия нервной системы человека
Рене Декарт
Ответная реакция по Декарту

Слайд 13Начало электрофизиологии обычно связывают со знаменитыми опытами итальянского врача, анатома и
История развития нейрофизиологии

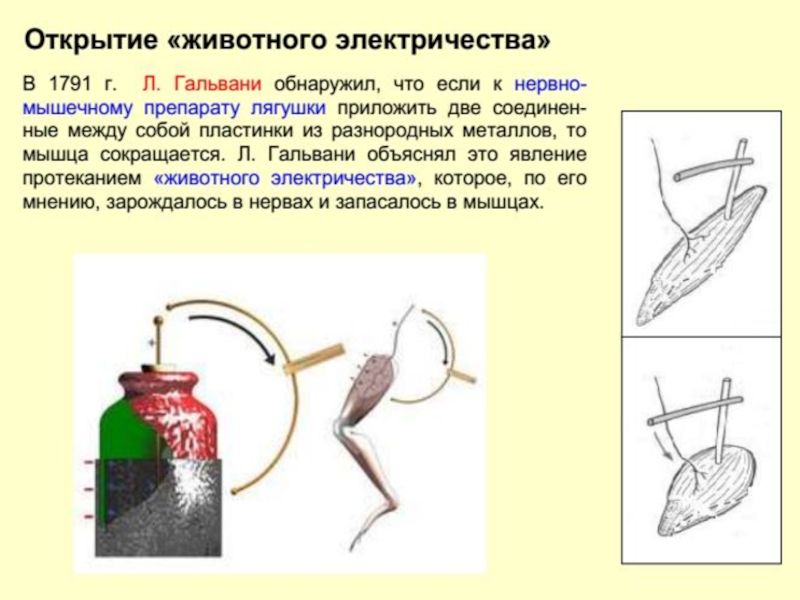
Слайд 15Карло Маттеучи (Carlo Matteucci 1811 – 1868) в 1830-1840 годах показал,
Распределение электрических зарядов в нервно-мышечном препарате (А) и разность потенциалов (Б), регистрируемая между неповрежденной и поврежденной поверхностями мышцы (по Маттеучи)
 в 1830-1840 годах показал, что в мышце всегда")

Слайд 17Опыт по электрофизиологии нервов и мышц
Эмиль Дюбуа Реймон
Первая публикация о наличии токов в центральной нервной системе (ЦНС) была сделана основоположником электрофизиологии нервов и мышц Эмилем Дюбуа Реймоном.
История развития нейрофизиологии

Слайд 18 В 1875 году английский хирург и физиолог Ричард Кэтон на заседании
Таким образом, было показано, что мозг является генератором электрической активности, то есть были открыты биотоки мозга.
Richard Caton
(1842 – 1926)

Слайд 19Василий Яковлевич
Данилевский
В 1875 г. В.Я. Данилевский изложил данные по изучению
Выделил две основные формы активности – фоновую, то есть спонтанно возникающую в спокойном состоянии животного, при видимом отсутствии раздражений рецепторов и проводящих путей и вызванную – на свет, звук, при электрической стимуляции седалищного нерва, а так же на запах аммиака, амилнитрита и пищи.

Слайд 20Иван Михайлович
Сеченов
1864 г. «Рефлексы головного мозга»
В 1882 году И.М.Сеченов опубликовал

Слайд 21Иван Михайлович Сеченов
1829 – 1905
В 1863 г. опубликовал книгу

Слайд 22Он обосновал возможность распространения принципа рефлекса как принципа организации поведения на

Слайд 23В 1884 году Н.Е. Введенский в работе “Исследования над нервными центрами”
Николай Евгеньевич
Введенский
(1852 – 1922)

Слайд 24Б.Ф. Вериго
В 1889 г. В журнале «Вестник клинической и судебной психиатрии

Слайд 251890 год - работа А. Бека, посвященная использованию метода регистрации электрической

Слайд 261898 год - В.Е. Ларионов производит исследования электрических явлений в головном
Соответствие между высотой музыкального тона и участком мозга, где локализованы, по мнению Ларионова, тоновые центры, обнаруживалось появлением отрицательного колебания; несоответствие – положительного колебания.

Слайд 271900 год – работа С.А. Тривуса, который получил определенные изменения потенциалов
Схема экспериментальной установки С.А. Тривуса

Слайд 281912 год - обширное исследование П.Ю. Кауфмана, который в тщательных опытах

Слайд 29Первая запись биотоков головного мозга человека получена Гансом Бергером в 1925
Ганс Бергер
Первая запись биотоков головного мозга человека

Слайд 30Нейрофизиологические методы:
Внутриклеточная регистрация активности нервных клеток (ПП, ПД)
Внеклеточная регистрация нейронной активности
Гистологические (гистохимические) методы – оценка изменения активности клеток по накоплению в них различных веществ (аминокислот, белков и др.)
Внеклеточная регистрация нейронной активности (ПД, ЭЭГ)Гистологические (гистохимические) методы")

Слайд 32Регистрация импульсной активности нервных клеток
Фотографии нейронов, окрашенных флуоресцентным красителем
Подведение электрода к

Слайд 33Два подхода к изучению мозга
Гемодинамика (метаболизм) – ПЭТ, ЯМРИ
Электрофизиология – ЭЭГ,
 – ПЭТ, ЯМРИЭлектрофизиология – ЭЭГ, МЭГ")

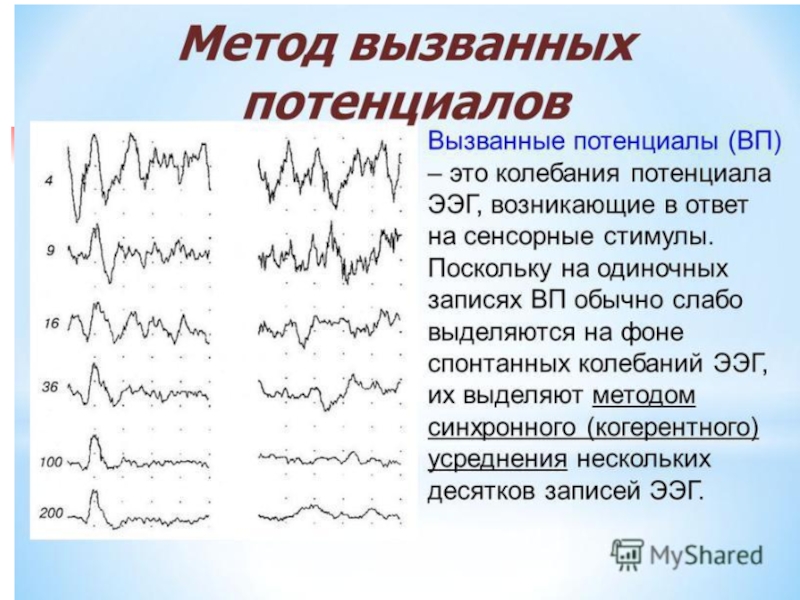
Слайд 35Основные ритмы ЭЭГ
Альфа-ритм, 8-13 Гц
50-100 мкВ
Дельта-ритм, 0.5-4 Гц
20-200 мкВ
Бета-ритм, 14-30
5-30 мкВ
Гамма-ритм, 30-60 (150, 500) Гц
2-15 мкВ
Тета-ритм, 4-7 Гц, 20-100 мкВ


Слайд 38Современные методы исследования мозга человека, позволяющие локализовать функции
Компьютерная томография
Ядерно-магнитный резонанс
Магнитоэнцефалография
Позитронно-эмиссионная томография






Слайд 45Результаты ПЭТ-сканирования
ПET - изображение мозга, показывающее зрительное и слуховое распознавание



")

Слайд 51Возникновение современной нейронауки
На современном этапе функции нейрофизиологии построены на изучении интегративной

Слайд 52Задачи нейронауки
Объяснить поведение человека в терминах активности мозга.
Понять, как
Нейронаука изучает нервную систему на различных уровнях: от молекулярного к клеточному, затем к системному уровню и, наконец, когнитивному (познавательному).

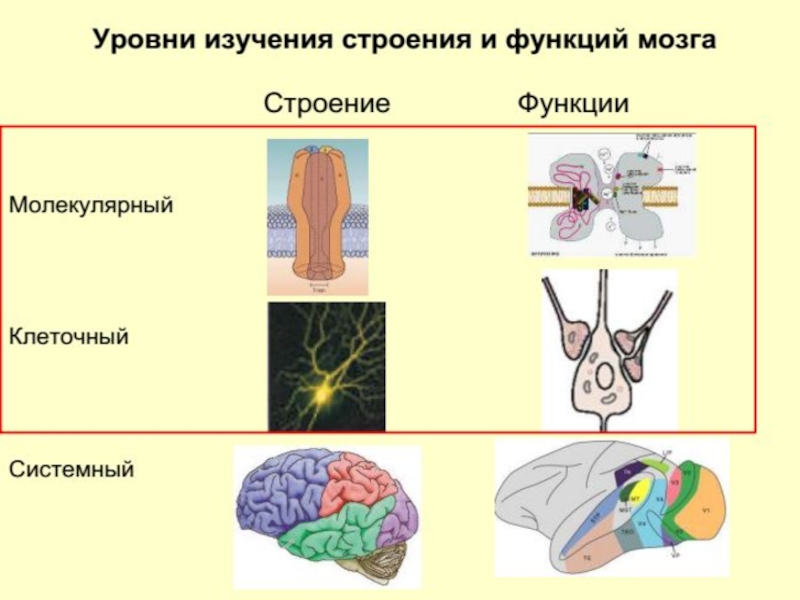
Слайд 54Молекулярный уровень
рассматриваются вопросы морфологии и физиологические особенности нейронов, а также взаимосвязь

Слайд 55Клеточный уровень
Оценка механизмов восприятия сигналов дендритами, сомами и аксонами нейронов, а

Слайд 56Системный уровень
Использование анатомических и физиологических знаний для изучения таких физиологических функций,

Слайд 57Когнитивный уровень
Оценка психологических функций с точки зрения формирования нервной системы.

Слайд 58Основные процессы в нервной системе
Трансдукция. Превращение раздражения, внешнего по отношению к
Трансформация. Переделка, преобразование входящего потока возбуждения в выходящий поток с отличающимися характеристиками.
Распределение. Распределение возбуждения и направление его по разным путям, по разным адресам.

Слайд 59Основные процессы в нервной системе
Моделирование. Построение нервной модели раздражения и/или раздражителя,
Модуляция. Нервная система под влиянием раздражения изменяет себя и/или свою деятельность.

Слайд 60Виды модуляции
1. Активация (возбуждение). Повышение активности нервной структуры, повышение её
2. Угнетение (торможение, ингибиция). Понижение активности нервной структуры, торможение.
. Повышение активности нервной структуры, повышение её возбуждения и/или возбудимости. Доминантное")
Слайд 61Виды модуляции
3. Пластическая перестройка нервной структуры.
Варианты пластических перестроек:
1) Сенситизация -
 Сенситизация - улучшение передачи")
Слайд 62Виды модуляции
4. Активация исполнительного органа для совершения действия.
Таким способом нервная


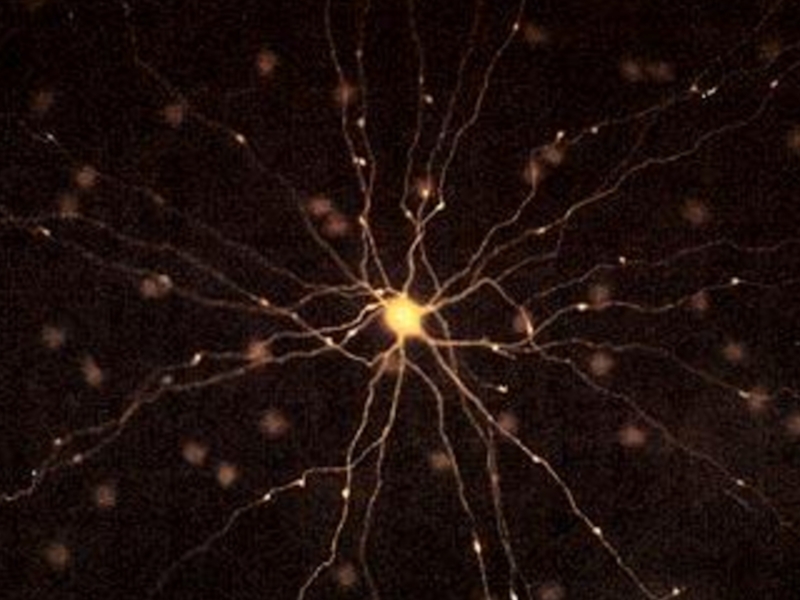


Слайд 67Нейрон – структурно-функциональная единица центральной-нервной системы.
Структурно-функциональная организация нервной системы

Слайд 68Тело (сома)
нейрона - центральная часть нейрона, обеспечивающая питанием все остальные
Строение нейрона
 нейрона - центральная часть нейрона, обеспечивающая питанием все остальные части клетки. Строение нейрона")
Слайд 69По отношению к отросткам тело клетки выполняют трофическую функцию -регулирует в

Слайд 70
Аксон – одиночный, обычно длинный выходной отросток нейрона, служащий для быстрого
По аксону транспортируются вещества, необходимые для синаптической передачи, пептиды, продукты нейросекреции.
Начальный сегмент аксона нейронов (аксональный холмик) является триггерной зоной – местом первоначальной генерации возбуждения.
Строение нейрона

Слайд 72Строение нейрона
Дендриты – древовидно-ветвящиеся отростки нейрона, его главное рецептивное поле,
От одной клетки может отходить от 1 до 1000 дендритов.

Слайд 73Строение нейрона
На дендритах имеются микроскопических размеров выросты (шипики), которые значительно увеличивают
Особого развития шипики достигают на клетках больших полушарий головного мозга.
На каждом шипике может быть до 8 синапсов (межклеточных контактов).
, которые значительно увеличивают поверхность соприкосновения с другими")
Слайд 74Виды нейронов
А – униполярные: имеют один отросток – аксон. Может быть
Б – псевдоуниполярные: места отхождения аксона и дендрита очень близки, затем Т-образно делится на два отростка.

Слайд 75Виды нейронов
В –биполярные клетки: имеют два отростка – аксон и дендрит.
Г


Слайд 77Виды нейронов
По локализации:
Центральные - лежат в пределах ЦНС.
Периферические нейроны

Слайд 78Виды нейронов по выполняемой функции
Афферентные (чувствительные) нейроны обеспечивают восприятие раздражения и
Эфферентные (двигательные) нейроны обеспечивают передачу информации от ЦНС на периферию.
 нейроны обеспечивают восприятие раздражения и передачу информации в ЦНС.Эфферентные")
Слайд 79Виды нейронов по выполняемой функции
Вставочные нейроны обеспечивают передачу информации внутри ЦНС.
В
возбуждающие – оказывают возбуждающее влияние на эфферентные нейроны.
тормозные – оказывают тормозное влияние на эфферентные нейроны.

Слайд 80Тела нейронов образуют серое вещество головного мозга: кора, подкорковые ядра
Отростки нейронов

Слайд 81Глиальные клетки
Глиальные клетки в совокупности называют нейроглией. Это клетки, окружающие нейроны

Слайд 82Анатомическое расположение
клетки нейроглии в головном мозге;
шванновские клетки в периферической нервной системе.

Слайд 83Функции нейроглии
Опорная – поддерживает нервные клетки
Изолирующая – препятствует переходу нервных импульсов
Регуляторная – участвует в регуляции работы ЦНС, в частности, обеспечивая передачу импульсов в нужном направлении
Трофическая – участвует в обменных процессах нейронов

Слайд 84Функции нейроглии
Участие в регенерации. В отличие от нейронов, глиальные клетки сохраняют

Слайд 85Виды глиальных клеток
Эпиндемоциты - выстилают полости внутри мозга и имеют цилии
Астроциты - самые многочисленные глиальные клетки.
Плазматические (в сером веществе)
Волокнистые (в белом веществе)
Олигодендроциты - формируют миелин – электрическая изоляция аксонов.
Радиальная глия - играет роль в миграции нейронов при развитии мозга.

Слайд 86Астроциты
Функции
Гомеостатическая
(поддержание ионного
и химического состава
среды)
Метаболическая
(синтез и разложение
веществ)
Трофическая
(влияние на рост и
развитие нейронов)
Метаболическая(синтез и разложениевеществ)Трофическая(влияние на рост иразвитие нейронов)")
Слайд 87Олигодендроциты
Находятся преимущественно в белом веществе мозга, где они образуют миелин вокруг


Слайд 89
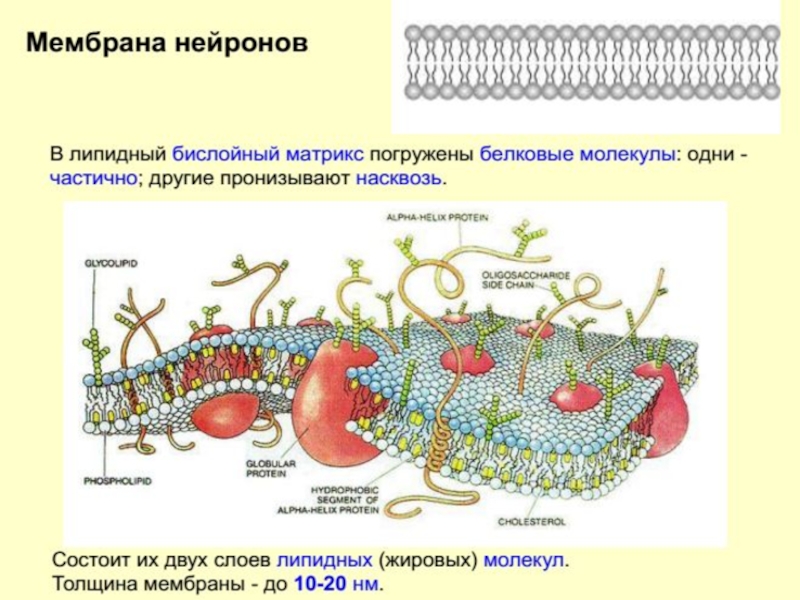
Тело нейрона покрыто слоистой мембраной, которая представляет собой два слоя липидов
Строение нейрона


Слайд 92Структуры, поддерживающие гомеостаз нейрона:
Мембрана – отделяет внутриклеточную среду от внеклеточной и
Ионные каналы – пассивный транспорт ионов по градиенту концентрации
Ионные каналы насосы – активный транспорт ионов против градиента концентрации с затратой энергии АТФ



Слайд 95Основные функции мембраны
Образует полупроницаемый, избирательный барьер между цитоплазмой и внешней средой
Регулирует обмен веществ между клеткой и окружающей средой.
Уникальный детектор и трансдуктор, приходящих стимулов.
Соединяет между собой клетки (межклеточные контакты) и прикрепляет клетки к внеклеточному матриксу.

Слайд 96Ионные каналы
1. Пассивные (для калия)
2. Управляемые:
потенциалзависимые;
хемозависимые;
механические
2. Управляемые:потенциалзависимые;хемозависимые;механические")
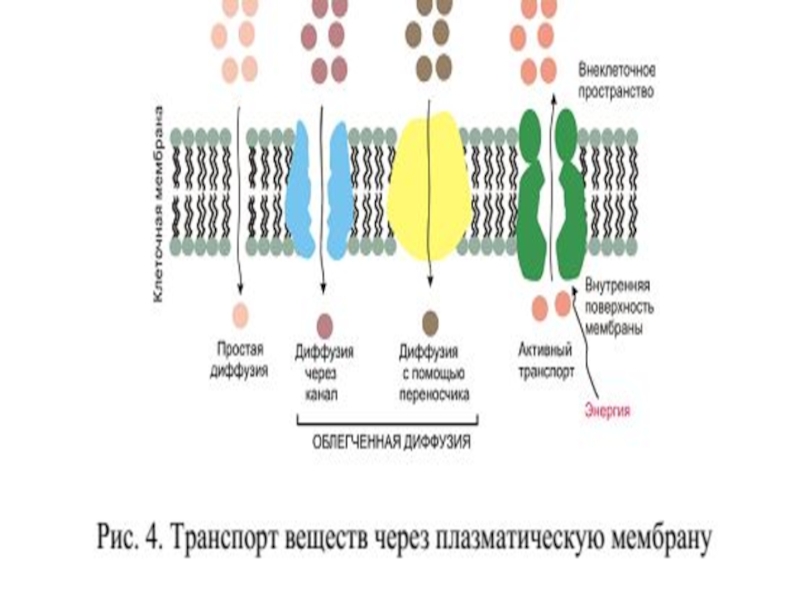
Слайд 98Мембрана называется полупроницаемой, потому что:
Мембрана ограничивает движение ионов какого-либо вида, т.е.
В результате этого возникает асимметрия в распределении ионов по обе стороны мембраны и (химический) градиент концентраций ионов

Слайд 99Благодаря этим свойствам мембрана создает и поддерживает
Химическую (концентрационную) асимметрию ионов по
Электрический градиент зарядов.
Поляризацию мембраны (диполи).
 асимметрию ионов по обе стороны мембраны.Электрический градиент")

Слайд 101Концентрационный градиент
Различная концентрация ионов является движущей силой для их диффузии в
Катионы калия диффундируют из клетки во внеклеточную жидкость (постоянно).
Катионы натрия и хлора диффундируют из внеклеточной жидкости в клетку (особенно интенсивно при открытии каналов в момент возбуждения нейрона).

Слайд 102Возникает и поддерживается МП
МП = ПП = Пассивный К+ ток.
МП регистрируют
Линг и Джерард впервые так сделали микроэлектрод


Слайд 104Основной принцип функционирования центральной нервной системы – способность мембраны нейронов генерировать
В формировании мембранного потенциала основную роль играет различная проницаемость мембраны нейрона для ионов калия и натрия.

Слайд 105Мембранный потенциал
это разность электрических потенциалов (напряжение) между внутренней и наружной поверхностью
 между внутренней и наружной поверхностью мембраны.")
Слайд 106Мембрана живой клетки поляризована — её внутренняя поверхность заряжена отрицательно по отношению
Мембрана обладает избирательной проницаемостью — её проницаемость для различных частиц (атомов или молекул) зависит от их размеров, электрического заряда и химических свойств.
Мембрана возбудимой клетки способна быстро менять свою проницаемостъ для определённого вида катионов, вызывая переход положительного заряда с внешней стороны на внутреннюю.

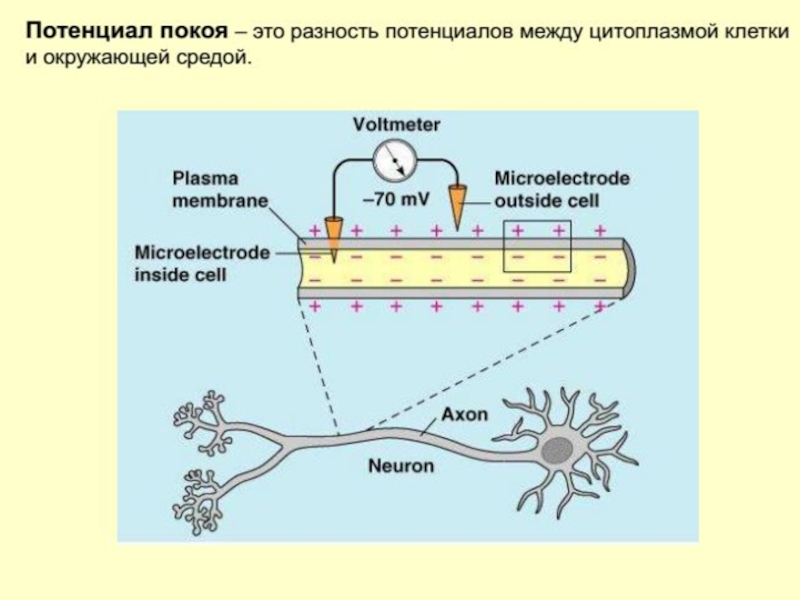
Слайд 108Потенциал покоя нейрона
Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя,

Слайд 109Потенциал покоя нейрона
Снаружи — преобладают ионы натрия, кальция и хлора, внутри — ионы калия и отрицательно

Слайд 110Под воздействием стимула происходит открытие натриевых каналов. По градиенту концентраций положительно
Открытие потенциал-зависимых натриевых каналов – активный ток ионов натрия внутрь клетки и смещение мембранного потенциала до + 20 мВ, что обеспечивает генерацию ПД.

Слайд 111Потенциал действия нейрона
Исходную концентрацию ионов восстанавливает натрий-калиевый насос.
Длительность ПД составляет около
Амплитуда ПД составляет 110-130 мВ.

Слайд 112Как распространяется возбуждение (потенциал действия) в нервной ткани?
Развития современных представлений
 в нервной ткани? Развития современных представлений о биоэлектрических явления способствовали")
Слайд 113Согласно этой теории биоэлектрических потенциалы обусловлены неодинаковой различной проницаемостью для них



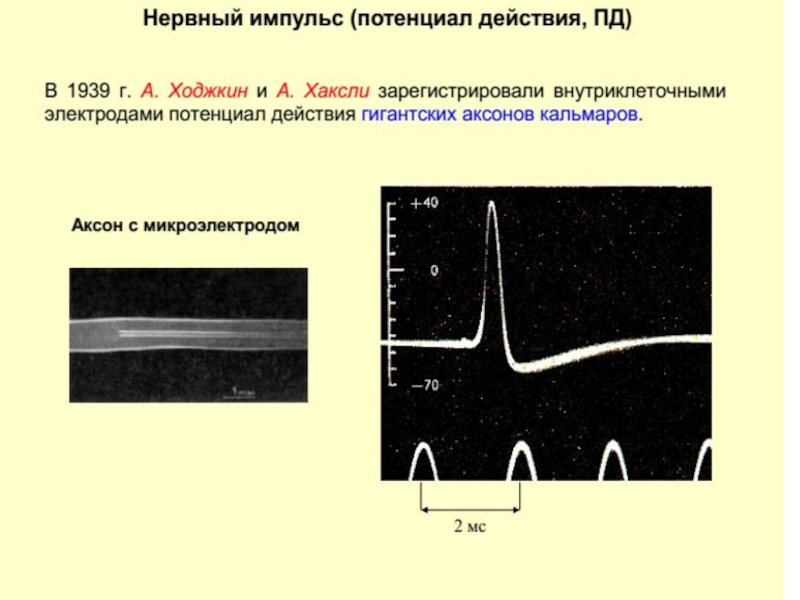
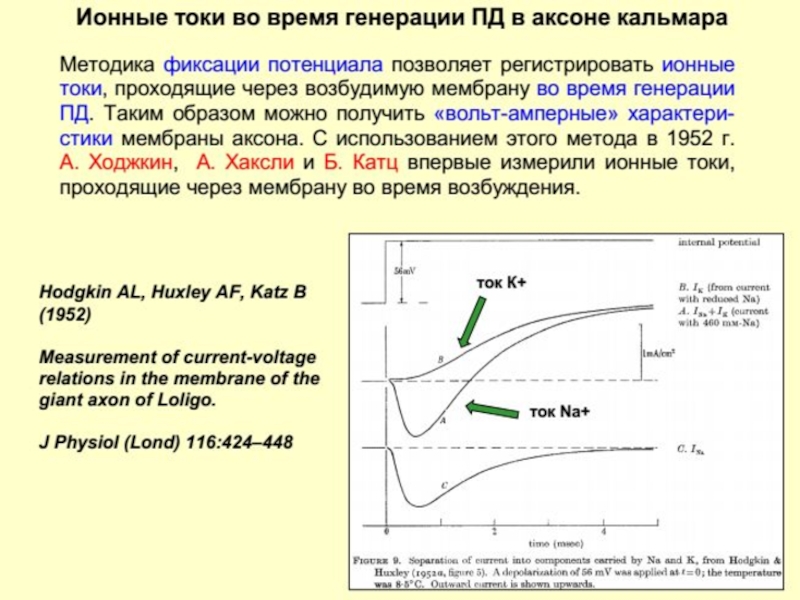
 или Нервный импульс")



Слайд 122В немиелинизированных нервных волокнах возбуждение распространяется посредством локальных токов очень маленькими
Распространение ПД по волокну

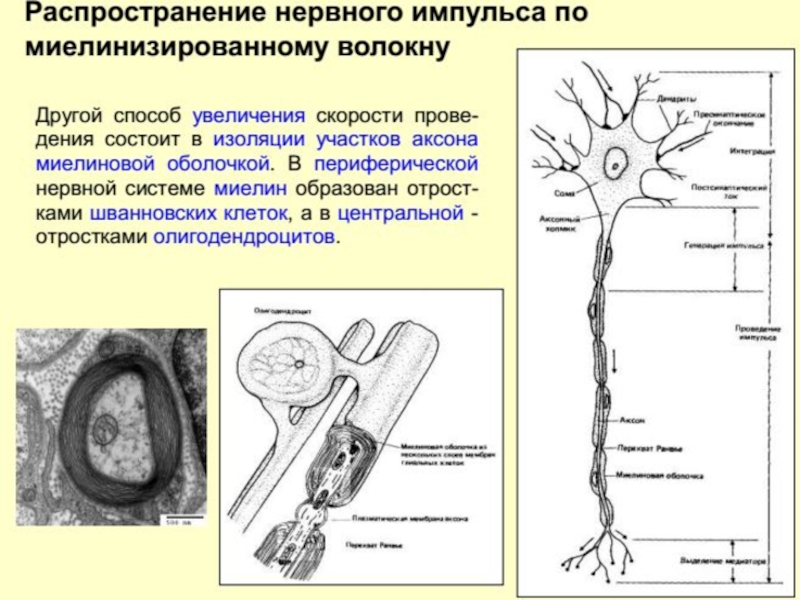
Слайд 123В миелинизированных волокнах, где поперечное сопротивление увеличивается за счет наличия дополнительной
Распространение ПД по волокну

Слайд 125Потенциал действия
Возникновению потенциала действия предшествуют активные изменения мембранного потенциала, которые обусловлены
Катионы натрия устремляются в клетку и уменьшают до нуля мембранный потенциал. На время возникает даже разность потенциалов с обратным знаком.
Внутренняя поверхность мембраны теряет отрицательный заряд и становится положительно заряженной, а внешняя, наоборот, теряет положительный заряд и заряжается отрицательно. Это фаза деполяризации.


Слайд 127В практической медицине используют эти фундаментальные знания
Местные анестетики блокируют натриевые каналы.
Натрий
Кардиологи управляют входом Кальция (лечение аритмий, гипертонии)

Слайд 128Закономерности проведения возбуждения по нервным волокнам
Двустороннее проведение возбуждения.
Изолированное проведение возбуждения в
Большая скорость проведения возбуждения.
Неутомляемость нервного волокна.
Возможность функционального блока проведения возбуждения при морфологической целостности нервных волокон.











