- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Репликация и экспрессии геномов ДНК-вирусов бактерий и эукариот презентация
Содержание
- 1. Репликация и экспрессии геномов ДНК-вирусов бактерий и эукариот
- 4. Роджер и Артур Корнберги
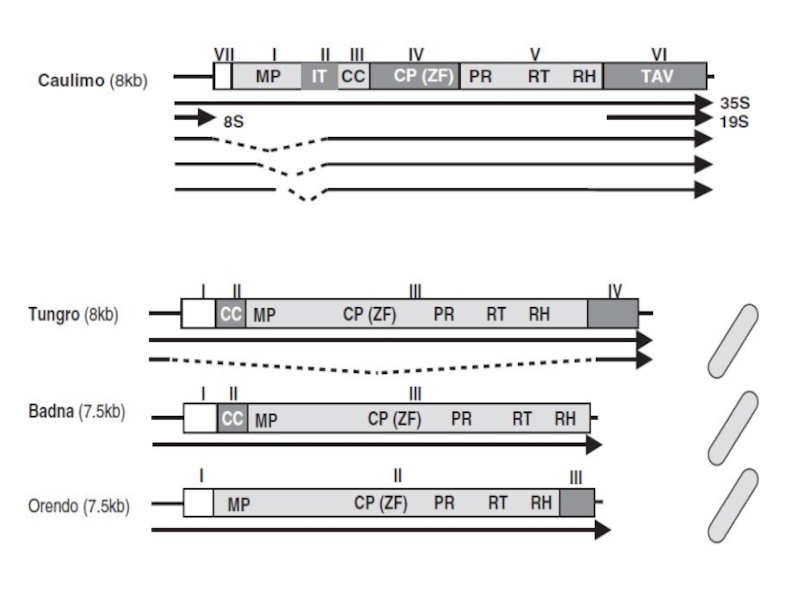
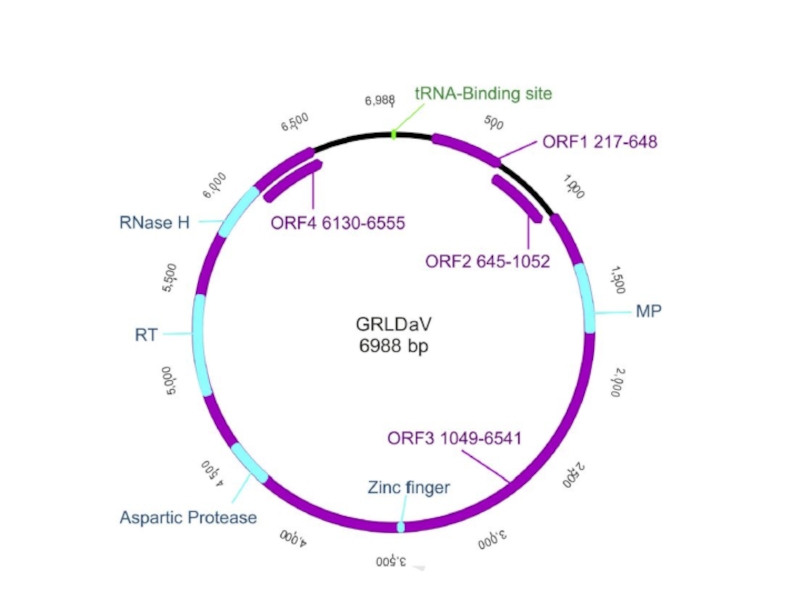
- 5. ДНК-вирусы растений. Каулимовирусы.
- 10. Каулимовирусы (Caulimoviruses)
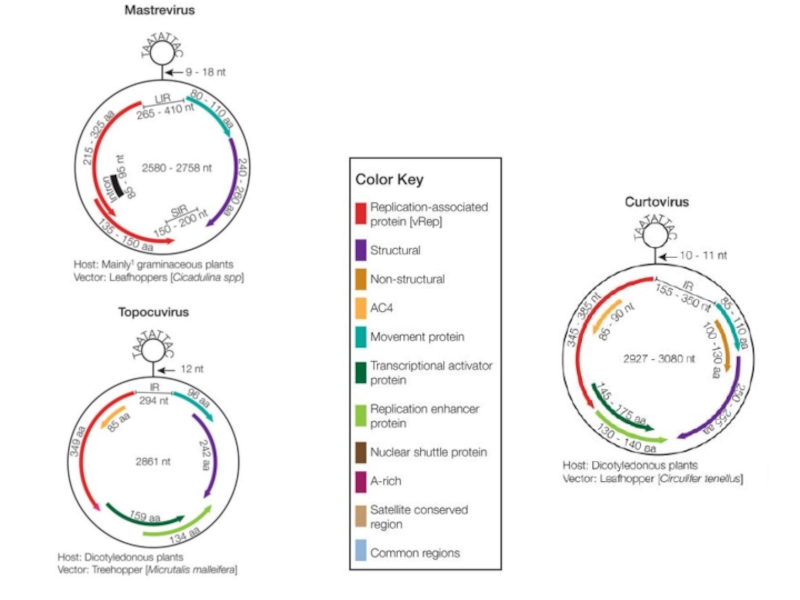
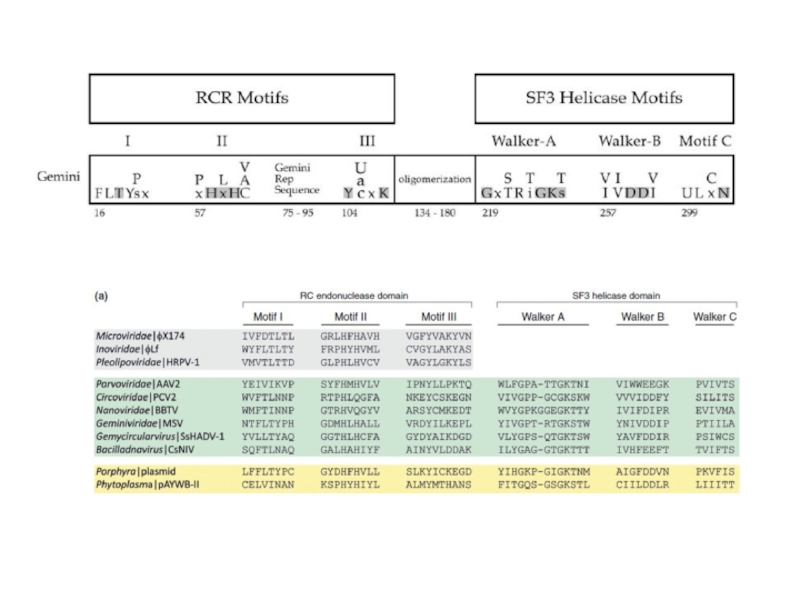
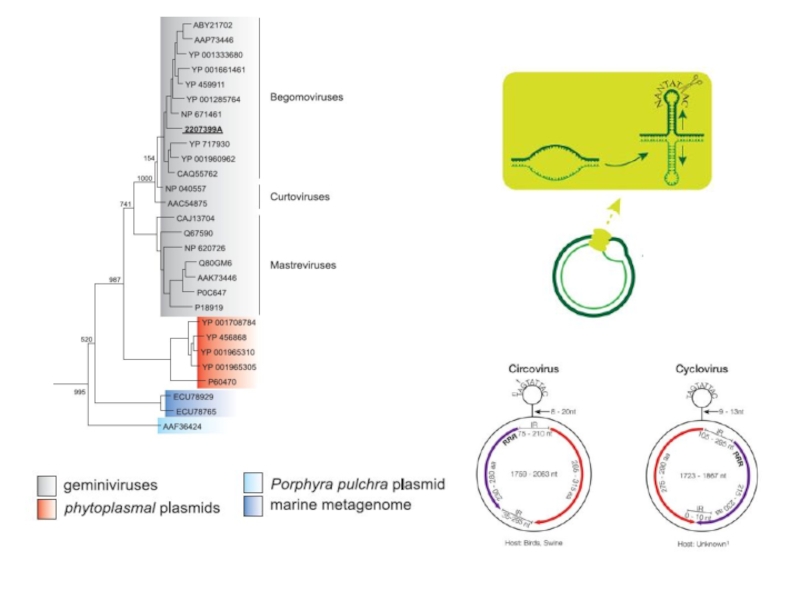
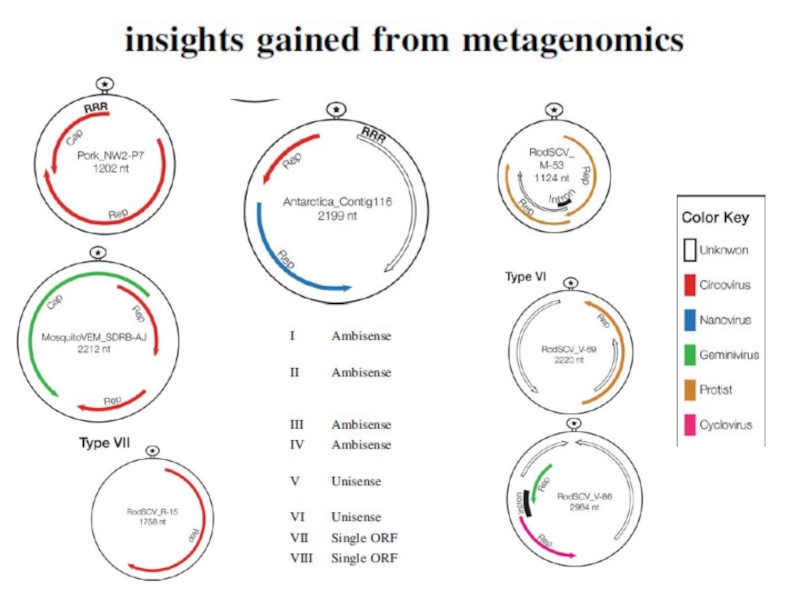
- 14. ДНК-вирусы растений. Геминивирусы.
- 15. Bean golden mosaic virus genus
- 19. Banana bunchy top virus
- 20. Faba bean necrotic yellows virus
- 23. Репликация и экспрессии геномов ДНК-вирусов бактерий и эукариот
- 25. Роджер и Артур Корнберги
- 27. Бактериофаги
- 28. РНК-содержащие бактериофаги
- 29. Механизмы синтеза РНК довольно консервативны и
- 30. Принципы регуляции транскрипции: 1. Системность. Один
- 31. Как правило, регулируемыми стадиями
- 32. Можно различить несколько уровней сложности систем
- 33. Бактериальная ДНК-зависимая РНК-полимераза включает “коровую“ часть, включающую
- 34. Частота инициации зависит от силы промотора.
- 35. В комплексе с другими субъединицами σ-фактор, включающий
- 36. Домены, расположенные на дистальных концах σ-субъединицы, узнают
- 37. Сродство промотора и σ-субъединицы зависит прежде
- 38. РНК-полимераза связывает промотор поэтапно. Сначала формируется
- 39. Терминация транскрипции. У прокариот имеется два
- 41. Итак, комбинацией промоторов и терминаторов различной
- 42. Бактериофаги
- 44. В геноме фагов есть две транскрипционные единицы.
- 45. Такой принцип количественной транскрипционной регуляции называется
- 47. Итак, фаг φХ174 дает нам прекрасный
- 48. Регуляция экспрессии генов фага λ Фаг
- 49. Жизненный цикл литических фагов
- 50. Явление лизогении
- 51. Индукция профага
- 52. Устройство генома лямбдоидных фагов
- 53. БАКТЕРИОФАГ ЛЯМБДА
- 54. На ранней стадии происходит активация транскрипции с
- 55. С промотора PL происходит транскрипция гена N.
- 56. Мотивы nut состоят из последовательности box A
- 58. Важнейшая роль на стадии выбора пути инфекции
- 59. В клетке активность системы Hfl регулируется системой
- 60. Белок CII длиной 97 аминокислот действует как
- 62. Белок CII длиной 97 аминокислот действует как
- 63. БАКТЕРИОФАГ ЛЯМБДА
- 64. Нужны внешние воздействия для выхода фага из
- 65. БАКТЕРИОФАГ ЛЯМБДА
- 66. Белок cro длиной 66 остатков, содержит только
- 67. Белок Q, как и N, является антитерминатором
- 68. БАКТЕРИОФАГ ЛЯМБДА
- 69. Сборка головки фага
- 70. Сборка хвостового отростка
- 71. БАКТЕРИОФАГ Т4
- 72. Гены Т4 делятся на истинно ранние,
- 73. Среди продуктов истинно ранних генов фага
- 74. Для транскрипции средних генов нужны вирусные
- 76. Промоторы поздних генов содержат элементы, существенно
- 77. Известно, что gp55 является фактором, обеспечивающим
- 78. Alt (ModA, ModB) АДФ-рибозилтрансферазы
- 79. БАКТЕРИОФАГ Т7
- 80. Все гены фага Т7 транскрибируются с одной
- 81. Первоначально в клетку бактерии попадает левый конец
- 82. Ранние транскрипты у фага Т7 подвергаются
- 83. Важнейшую роль в инфекционном цикле фага
- 84. Пять фагоспецифических промоторов для генов класса III
- 85. Регуляция транскрипции у фага N4 Геном
- 88. Средние гены транскрибируются N4 РНК-полимеразой II,
- 90. Поздние гены транскрибируются клеточной σ70-содержащей РНК-полимеразой.
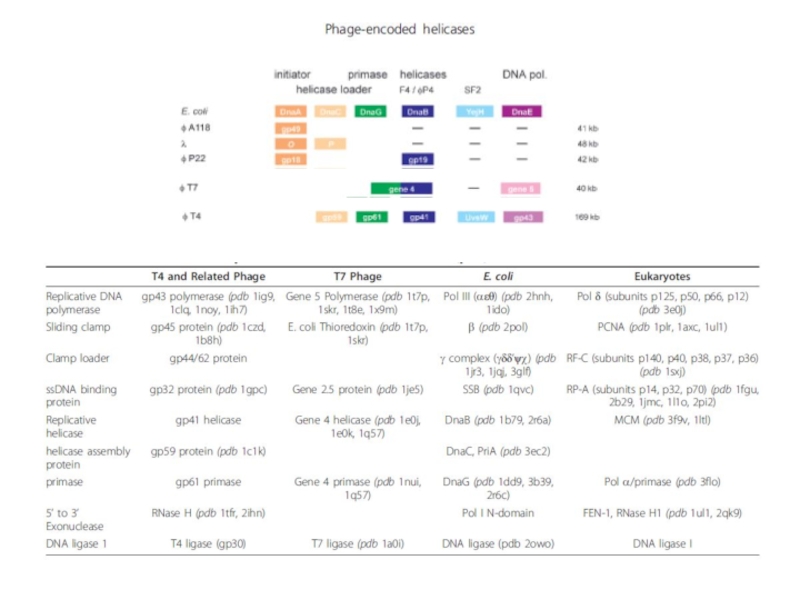
- 93. ДНК полимераза III E.coli Фермент состоит из
- 95. Enterobacteria phage P2 33.5 Kbp
- 96. Репликация ДНК бактериофага λ Инициация репликации. Участок
- 97. Если ассоциация белка О с
- 99. В ходе двунаправленной репликации по схеме Кернса
- 100. Созревание ДНК-конкатемеров В итоге поздней
- 101. Bacillus subtilis bacteriophage Φ29 21.1 Kbp
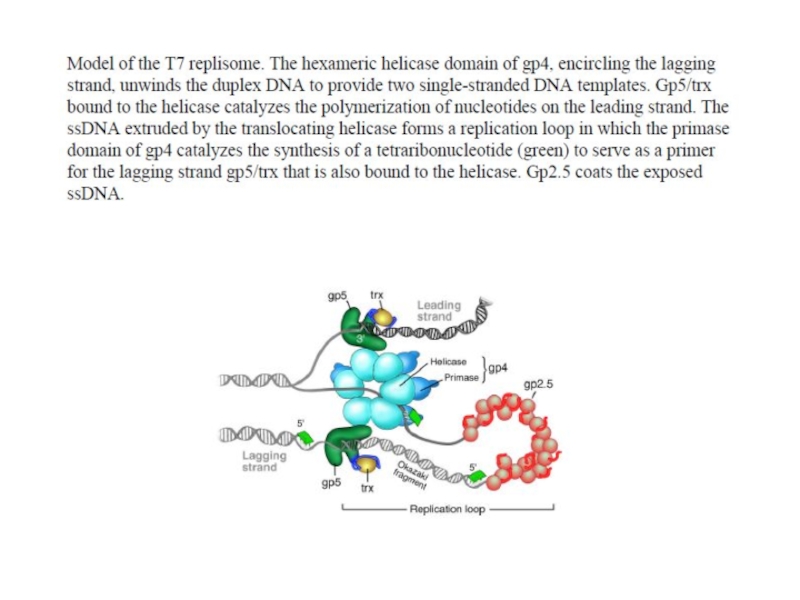
- 104. T7 phage
- 108. T7 phage
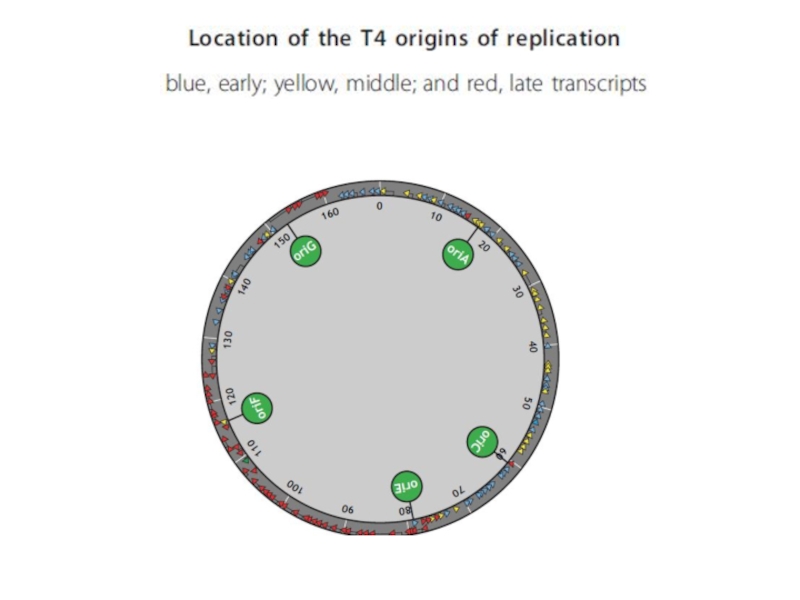
- 113. T4 bacteriophage





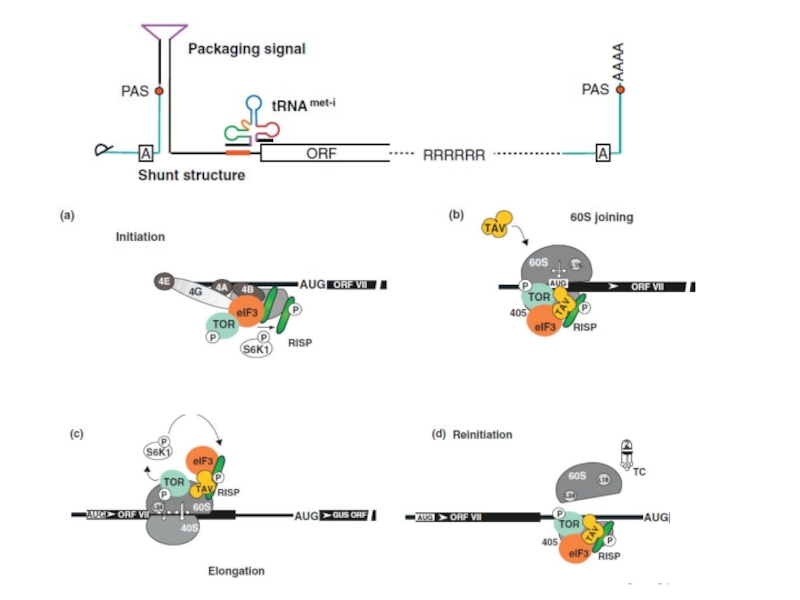
")






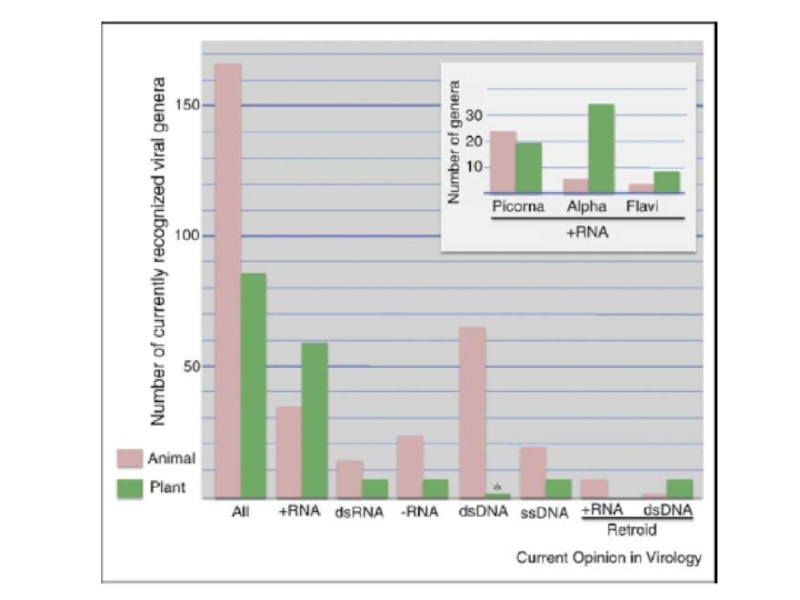

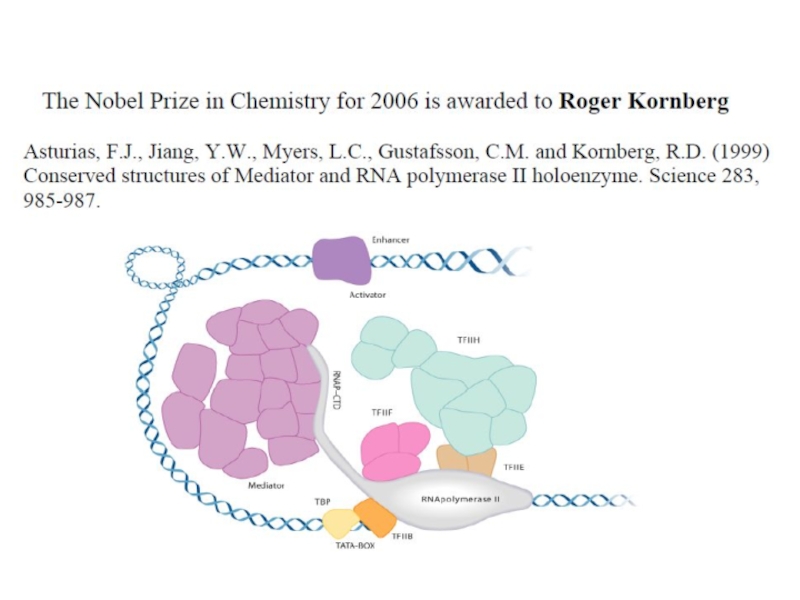


Слайд 29
Механизмы синтеза РНК довольно консервативны и однообразны среди вирусов. Однако, большое
Для чего нужна регуляция транскрипции геномной ДНК?
1. на разных стадиях цикла репродукции требуются разные белки.
2. белки нужны в разных количествах (требуется много структурных белков и меньше энзимов).
3. избыток ранних белков может подавлять активность белков поздних стадий.
Уровни регуляции экспрессии генов:
1. транскрипционный
2. посттранскрипционный (чаще трансляционный)

Слайд 30
Принципы регуляции транскрипции:
1. Системность. Один механизм регуляции транскрипции может контролировать экспрессию
2. Каскадность. Включение экспрессии одних генов может быть связано с репрессией или активацией других генов.
Белки вирусов условно делят на:
1. ранние (истинно ранние).
2. средние (отсроченно ранние).
3. поздние.

Слайд 31
Как правило, регулируемыми стадиями транскрипции являются инициация и терминация. В
Выход продукта экспрессии гена зависит от:
1. частоты инициации транскрипции
2. длины гена (положение терминатра)

Слайд 32
Можно различить несколько уровней сложности систем транскрипции фаговых геномов в зависимости
В наиболее простых системах, когда комбинацией промоторов и терминаторов различной силы регулируется количество синтезируемой мРНК, используются лишь белки аппарата транскрипции хозяина.
Другие вирусы кодируют регуляторные белки активации и репрессии транскрипции, третьи модифицируют отдельные субъединицы хозяйской РНК-полимеразы.
И наконец, существуют варианты полного замещения хозяйского аппарата транскрипции на фагоспецифический.

Слайд 33Бактериальная ДНК-зависимая РНК-полимераза включает “коровую“ часть, включающую две α субъединицы, β- и

Слайд 34
Частота инициации зависит от силы промотора. Чем сильнее промотор, тем выше
За узнавание промотора и определение стартовой точки транскрипции у прокариот отвечает σ-субъединица РНК-полимеразы. Основной тип σ-субъединиц – это σ70, отвечающая за транскрипцию большинства генов бактерий. Другие σ-факторы используются лишь при специфических условиях роста или при стрессе.

Слайд 35В комплексе с другими субъединицами σ-фактор, включающий 613 аминокислотных остатков имеет

Слайд 36Домены, расположенные на дистальных концах σ-субъединицы, узнают два элемента промотора. Стандартный

Слайд 37
Сродство промотора и σ-субъединицы зависит прежде всего от оптимального расстояния между
Целый ряд σ70-зависимых промоторов обладают далеко не оптимальной для узнавания структурой –35 элемента и, тем не менее, весьма активно узнаются РНК-полимеразой. Такие промоторы содержат удлиненный –10 консенсус типа 5’-TGNTATAAT-3’.
Известно, что в некоторых случаях CTD-домен α-субъединиц узнает последовательности в области от -40 до –60 (UP элементы), что может резко увеличивать силу промотора.

Слайд 38 РНК-полимераза связывает промотор поэтапно. Сначала формируется достаточно слабый так называемый

Слайд 39
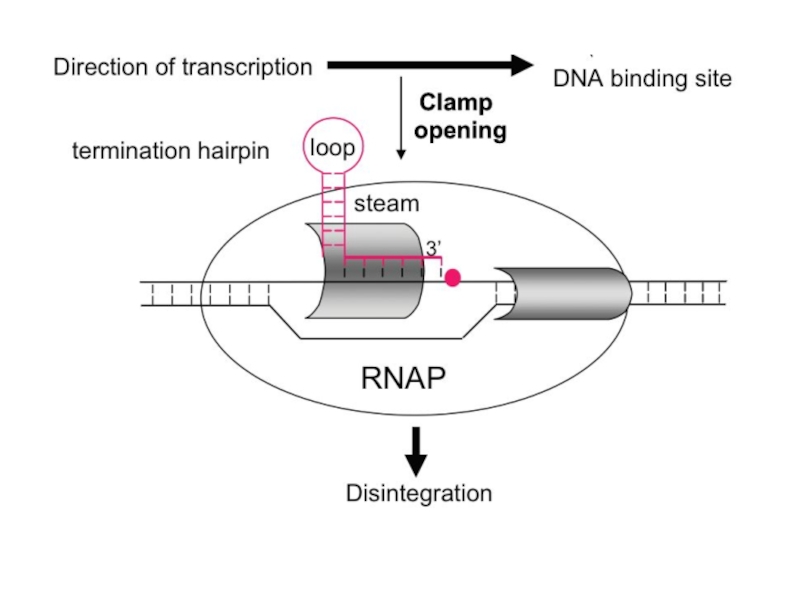
Терминация транскрипции.
У прокариот имеется два механизма регуляции терминации транскрипции. Это ρ
ρ-фактор представляет собой олигомерный белок (гексамер) функционирующий как РНК-ДНК хеликаза (3'-5'), которая в АТФ-зависимой реакции расплетает гетеродуплекс РНК-ДНК.
В ρ-независимой терминации участвуют специальные сигналы РНК, задерживающие работу РНК-полимеразы. Чаще всего сигналами ρ-независимой терминации являются GC-богатые шпилечные участки мРНК, за которыми следует U-богатая область.
В результате возможны остановки полимеразы. В момент паузы возможен стохастический выбор между терминацией синтеза и продолжением процесса транскрипции. Это создает возможность снижения в той или иной степени эффективности транскрипции отдельных генов и оперонов в зависимости от встречаемости сегментов ДНК, способствующих паузам.
-зависимая терминация и ρ-независимая")
Слайд 41
Итак, комбинацией промоторов и терминаторов различной силы можно регулировать количество синтезируемой
Примером простой системы, в которой отсутствует временная, но есть количественная регуляция транскрипции, являются фаги с кольцевой одноцепочечной ДНК.


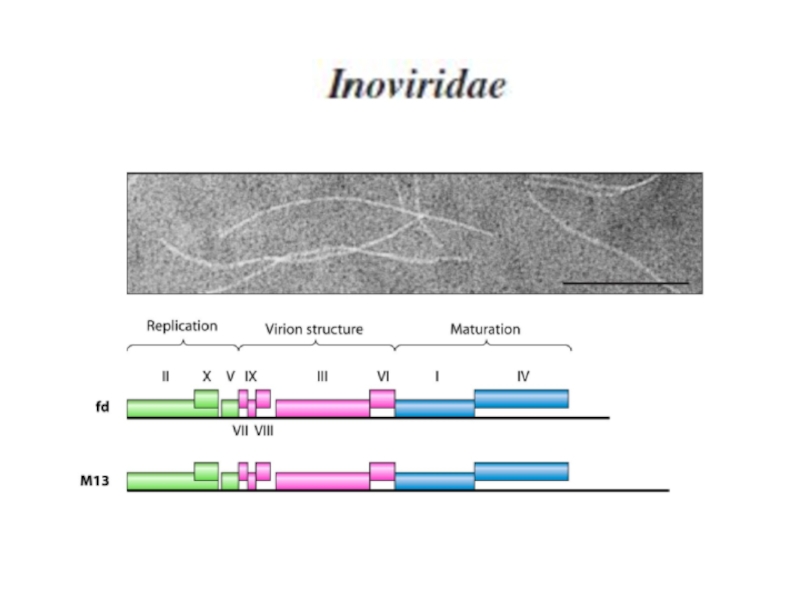
Слайд 44В геноме фагов есть две транскрипционные единицы. Одна из транскрипционных единиц

Слайд 45
Такой принцип количественной транскрипционной регуляции называется каскадным (не путать с темпоральной
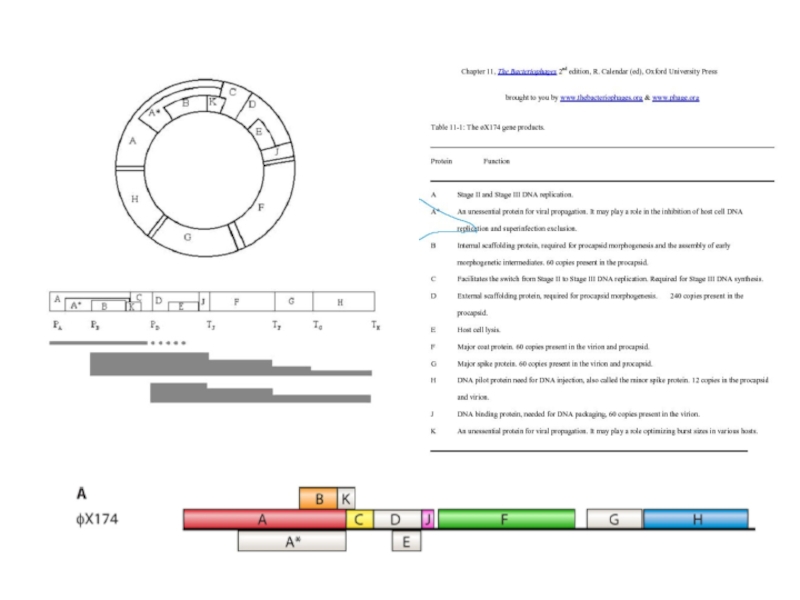
Возможно, еще более яркий пример сочетания принципа каскадности на уровнях инициации и терминации транскрипции представляет фаг φХ174.
.Возможно, еще более")
Слайд 47
Итак, фаг φХ174 дает нам прекрасный пример позитивной каскадной количественной регуляции,
С другой стороны, столь же отчетливо наблюдается негативная каскадная регуляция в случае, если ген отделен от промоторов одним или несколькими терминаторами транскрипции.

Слайд 48Регуляция экспрессии генов фага λ
Фаг λ - Умеренный бактериофаг
48,502 bp
Система регуляции транскрипции фага лямбда несравненно сложнее, чем у фагов М13 и φХ174.






Слайд 54На ранней стадии происходит активация транскрипции с помощью клеточных белков. Белок

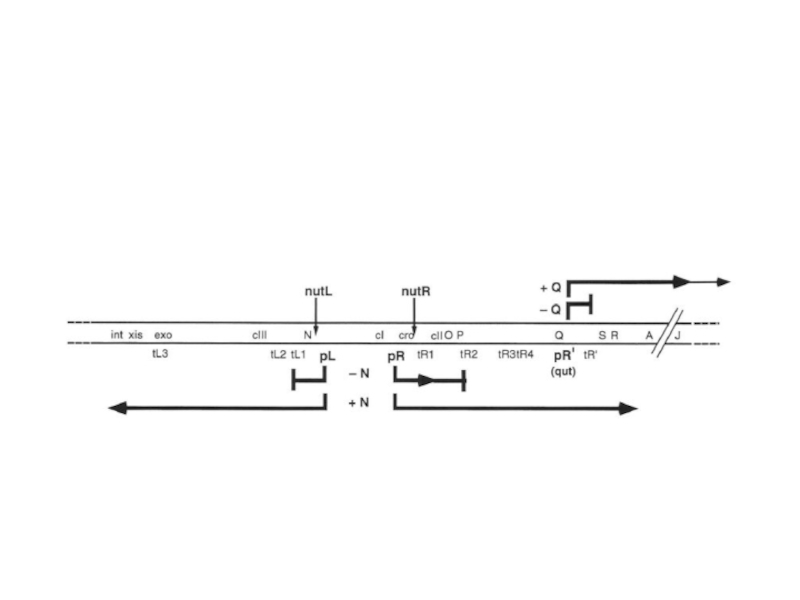
Слайд 55С промотора PL происходит транскрипция гена N. Белок N – это

Слайд 56Мотивы nut состоят из последовательности box A длиной 8 н.т., спейсера
nut | N | NusA | NusG | S 10 | RP

Слайд 58Важнейшая роль на стадии выбора пути инфекции принадлежит белку CII. Основные

Слайд 59В клетке активность системы Hfl регулируется системой катаболитной репрессии, включающей сAMP
.")
Слайд 60Белок CII длиной 97 аминокислот действует как транскрипционный фактор, открывающий новые

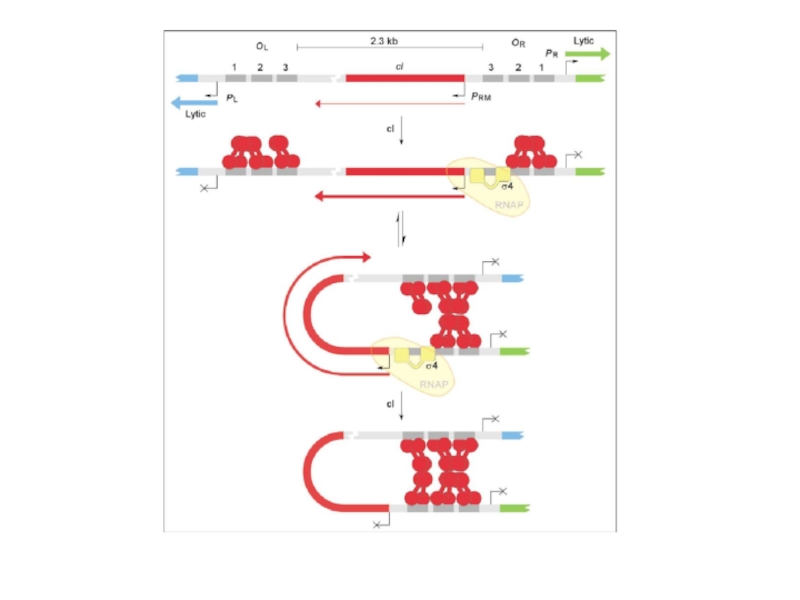
Слайд 62Белок CII длиной 97 аминокислот действует как транскрипционный фактор, открывающий новые


Слайд 64Нужны внешние воздействия для выхода фага из лизогенного состояния. Таким событием,


Слайд 66Белок cro длиной 66 остатков, содержит только один домен и узнает

Слайд 67Белок Q, как и N, является антитерминатором транскрипции и абсолютно необходим





Слайд 72
Гены Т4 делятся на истинно ранние, ранние, средние и поздние. Около

Слайд 73
Среди продуктов истинно ранних генов фага обнаруживаются еще две АДФ-рибозил-трансферазы. Соответствующие

Слайд 74
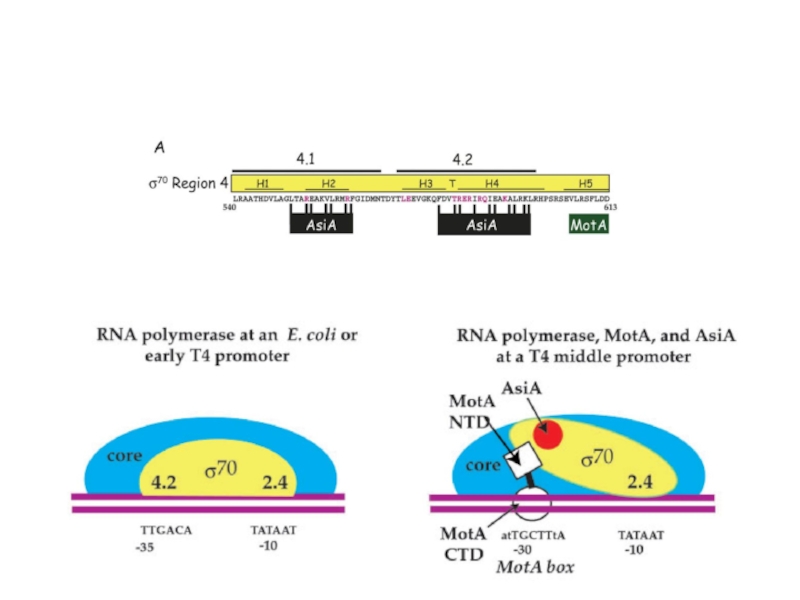
Для транскрипции средних генов нужны вирусные белки Asi A (antisigma) и
 и Mot (Modification of transcription).")
Слайд 76
Промоторы поздних генов содержат элементы, существенно отличающие их от клеточных промоторов.

Слайд 77
Известно, что gp55 является фактором, обеспечивающим весьма слабое сродство к ДНК

Слайд 78
Alt
(ModA, ModB)
АДФ-рибозилтрансферазы
для α-субъединиц РНК-полимеразы
Asi A (antisigma), Mot
Mod
 АДФ-рибозилтрансферазы для α-субъединиц РНК-полимеразы Asi A (antisigma), MotMod - АДФ-рибозилтрансфераза")

Слайд 80Все гены фага Т7 транскрибируются с одной и той же цепи

Слайд 81Первоначально в клетку бактерии попадает левый конец генома размером около 850

Слайд 82
Ранние транскрипты у фага Т7 подвергаются процессингу РНКазой III. Точек разрезания

Слайд 83 Важнейшую роль в инфекционном цикле фага играет ген 0.3, который
Среди ранних продуктов экспрессии появляется и РНК–полимераза фага, которая первоначально инициирует транскрипцию на 10 специфических промоторах, расположенных перед генами класса II. К этому классу относится и ген 2, кодирующий ингибитор клеточной РНК-полимеразы, который образует с ней прочный комплекс. Терминация транскрипции фаговой полимеразой происходит специфично на терминаторе, расположенном в районе поздних генов, и неспецифично при достижении правого конца генома

Слайд 84Пять фагоспецифических промоторов для генов класса III являются гораздо более сильными,

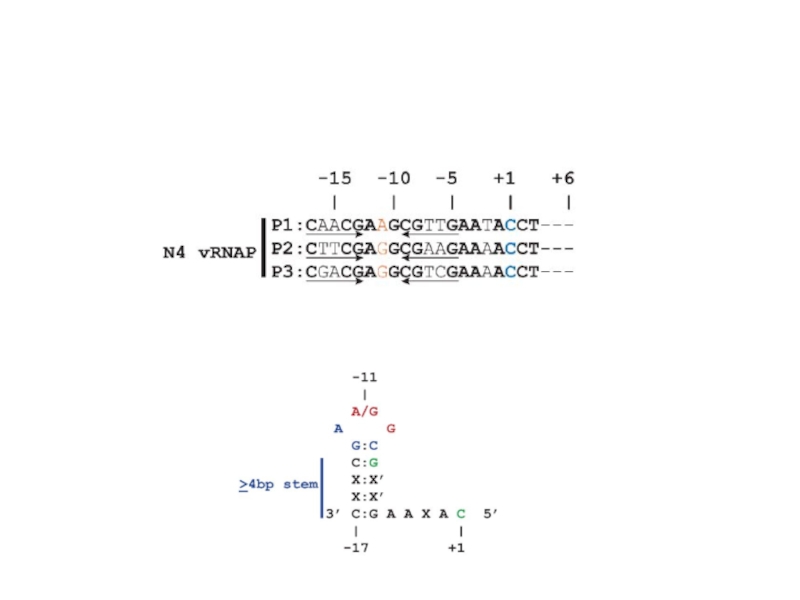
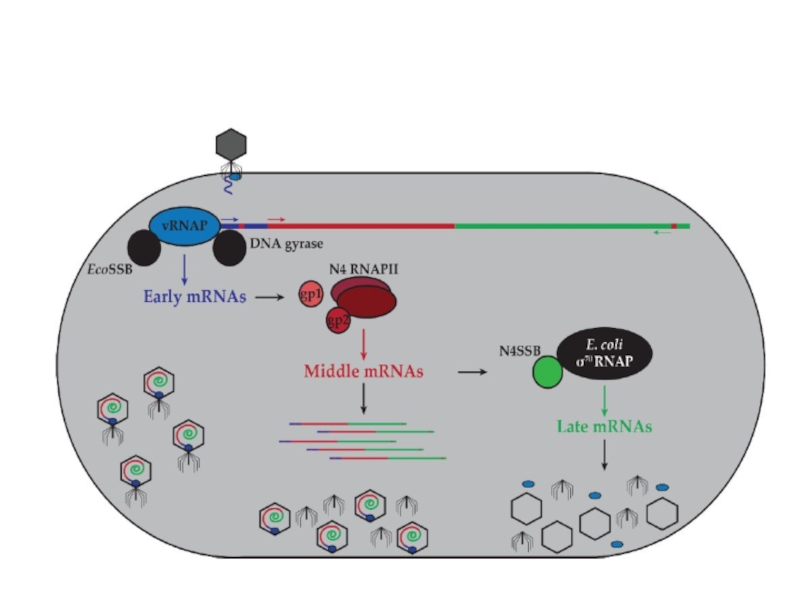
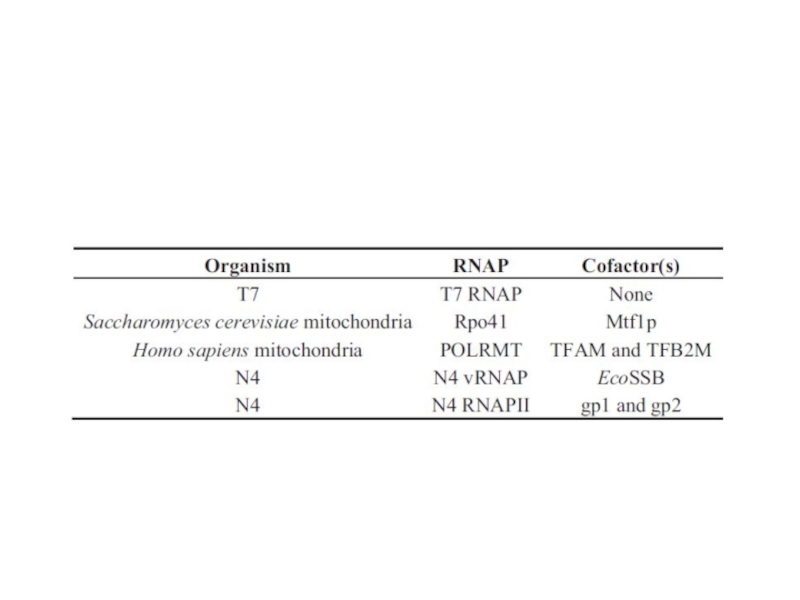
Слайд 85Регуляция транскрипции у фага N4 Геном этого фага представлен линейной двуцепочечной ДНК
Ранние гены транскрибируются фаговой РНК-полимеразой длиной 3500 аминокислот, которая находится в вирионе в количестве 1-2 копий и вносится в клетку при инфекции. Этот фермент эволюционно отдаленно близок Т7 РНК-полимеразе. Вирионная РНК-полимераза не способна узнавать промоторы на двуцепочечной ДНК, и использует только сверхспирализованную ДНК вируса, образовавшуюся при действии ДНК-гиразы E.coli. Ранние промоторы фага представляют собой консервативные шпилечные структуры, включающие концевую петлю длиной 3 остатка и двуспиральный стебель длиной 5-7 пар оснований. Такие шпильки стабилизируются ssb-белком E.coli, связывающим прилежащие односпиральные участки.


Слайд 88
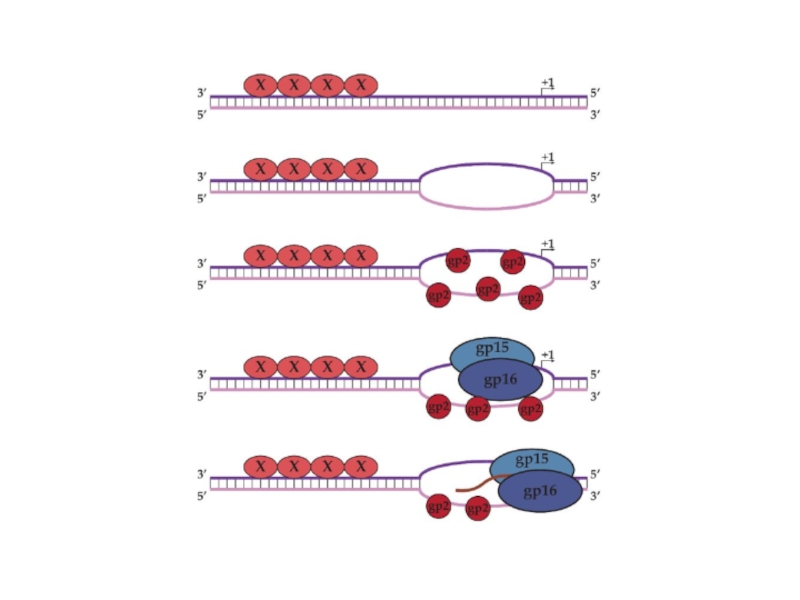
Средние гены транскрибируются N4 РНК-полимеразой II, являющейся продуктом ранних генов и

Слайд 90
Поздние гены транскрибируются клеточной σ70-содержащей РНК-полимеразой. Однако, поздние промоторы N4 проявляют

Слайд 93ДНК полимераза III E.coli
Фермент состоит из нескольких субъединиц и представляет собой
1) каталитический “соre” включает каталитические субъединицы α2 (синтез ДНК) и ε2 (экзонуклеазная 3’-5’активность);
2) β2-фактор процессивности полимеразы (аналог PCNA);
3) τ2, γ2,δ2,δ’2, χ, ψ – clamp loader (загружает фактор процессивности на ДНК с затратой АТФ). τ в нужный момент способствует посадке кольца β-субъединицы.
Наоборот, δ,γ,δ’ как «гаечный ключ» раскрывает β2-фактор.
Фактор процессивности снимается с ДНК, когда полимераза сталкивается с затравкой. Затравку затем удаляют РНКазаН и ДНК полимераза I, которая обладает 5’-3’ экзонуклеазной активностью и способна съедать последний рибонуклеотид затравки, связанный с дезоксирибонуклеотидом.

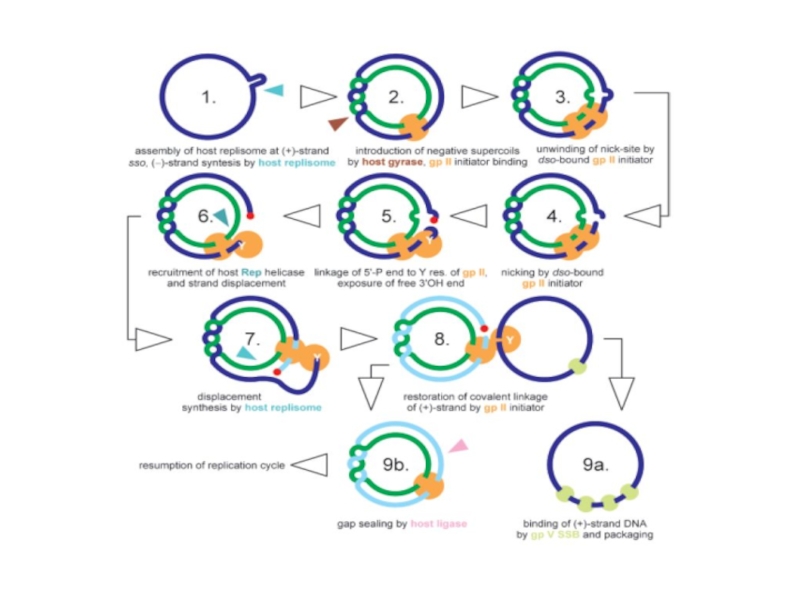

Слайд 96Репликация ДНК бактериофага λ
Инициация репликации. Участок инициации репликации Ori первоначально узнается

Слайд 97 Если ассоциация белка О с комплексом DnaB-Р-белок не произойдет (в случае

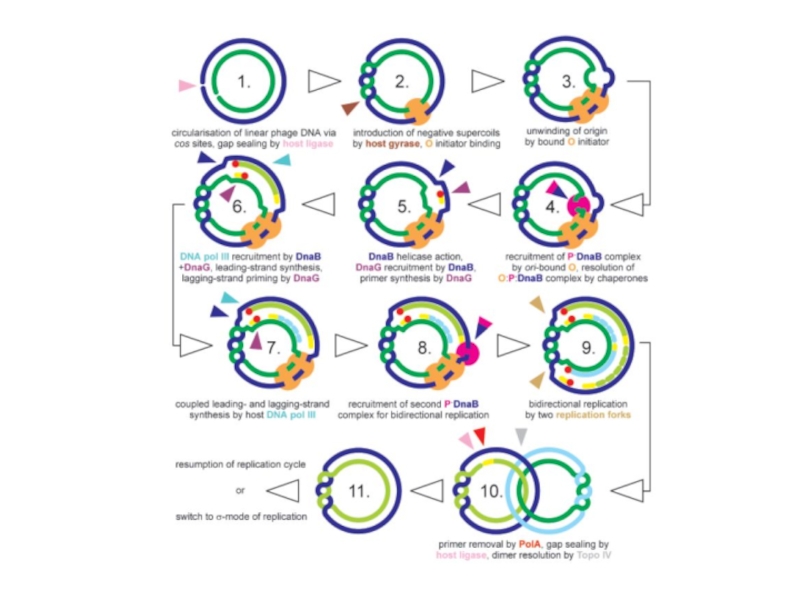
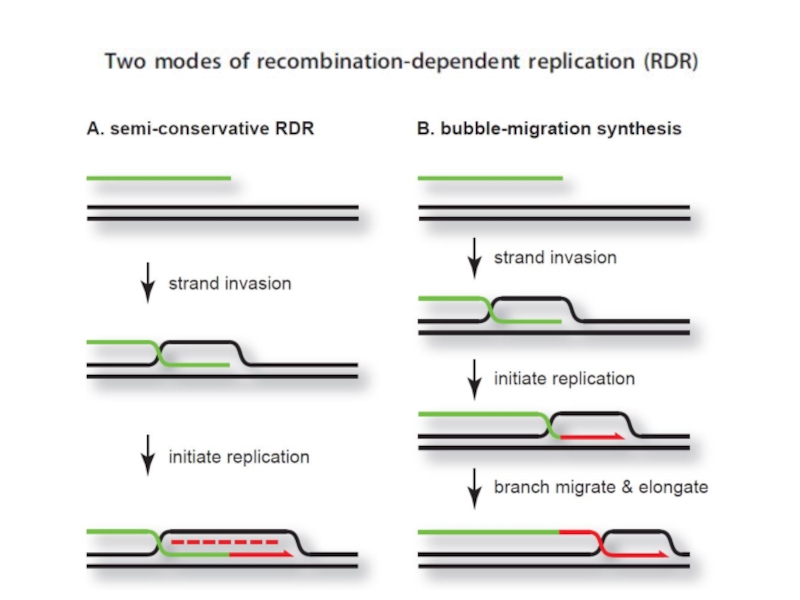
Слайд 99В ходе двунаправленной репликации по схеме Кернса в клетках, зараженных фагом
Превращение схемы Кернса в схему разматывающегося рулона, возможно, связано с ослаблением траскрипции в участке ori с промотера P® и, как следствие, возникновением односторонней репликации. Причиной ослабления транскрипции c этого промотера может быть действие возрастающих количеств cro-репрессора. Кроме того, присутствие в клетке ограниченного количества белка DnaA, приводит к тому, что при увеличении числа копий фаговой ДНК, он вытитровывается, и транскрипция с промотера P® ослабляется. Действительно, показано, что в клетках бактерий, продуцирующих температурочувствительный мутант DnaA при 43 градусах, даже на очень ранних этапах инфекции репликация ДНК фага происходит почти исключительно по схеме разматывающегося рулона.

Слайд 100Созревание ДНК-конкатемеров
В итоге поздней репликации образуются конкатемерные молекулы, которые надо разрезать
Структура cos.
Белок А в виде димера присоединяется к CosB и вносит разрыв в CosN. С этого участка начинается упаковка ДНК. CosQ нужен для правильного процессирования противоположного конца ДНК, чтобы упаковывался только один эквивалент генома. Для действия терминазы необходимо также взаимодействие с cos районом (участки I1 и I2) клеточного фактора IHF, который сильно изгибает ДНК, а также вирусного белка Nu1 (узнает R1 , R2 и R3). Возможно, что процесс упаковки регулируется также путем взаимодействия белков капсида с терминазой.