пространственно разделены с ионными каналами и обеспечивают непрямую синаптическую передачу.
Синаптический эффект таких рецепторов состоит в инициации метаболических каскадов, управляющих состоянием ионных каналов.
Эти рецепторы вызывают эффекты посредством взаимодействия с ГТФ-связывающими белками (G-белками), поэтому их еще называют рецепторами, сопряженными с G-белками (англ., G-protein coupled receptors).
В некоторых случаях непрямые механизмы модулируют эффективность прямой синаптической передачи (процесс нейромодуляции) – через ауторецепторы и эндоканабиноидные рецепторы на пресинаптических мембранах.
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Метаботропные рецепторы презентация
Содержание
- 1. Метаботропные рецепторы
- 2. Мембранные рецепторы, активируемые медиаторами По механизму активации
- 3. Метаботропные рецепторы В настоящее время в геноме
- 4. Метаботропные рецепторы
- 5. The main five GPCR families: Glutamate (G),
- 6. OR- olfactory receptor ланцетник
- 7. Метаботропные рецепторы Рецепторы этих подсемейств не демонстрируют
- 8. Эволюционное родство родопсинподобных рецепторов To assemble this
- 9. Родопсинподобные метаботропные рецепторы Acetylcholine (M1, M2, M3,
- 10. Секретинподобные метаботропные рецепторы Brain-specific angiogenesis inhibitor (1,
- 11. Метаботропные глутаматные рецепторы Calcium-sensing receptor GABA B
- 12. Метаботропные рецепторы Первым метаботропным рецептором, который был
- 13. Семейства метаботропных рецепторов Красными кружками отмечены характерные
- 14. Родопсинподобные метаботропные рецепторы наиболее изучены и включают
- 15. Родопсинподобные метаботропные рецепторы Цистеиновый остаток на участке
- 16. Родопсинподобные метаботропные рецепторы Amino acid sequence and
- 17. Секретинподобные метаботропные рецепторы связываются с секретином, кальцитонином,
- 18. Глютаматные метаботропные рецепторы включают восемь типов метаботропных
- 19. Олигомеризация метаботропных рецепторов Метаботропные рецепторы являются олигомерами.
- 20. Рецептор-олигомер представлен комплексом из нескольких рецепторов-субъединиц, связанных
- 21. Димеризация метаботропных рецепторов Some GPCRs can function
- 23. When 5-HT2A and mGlu2 are prevented from
- 26. a. Schematic representation of ligand-induced heterodimerization by
- 27. Десенситизация метаботропных рецепторов Десенситизация рецепторов заключается в
- 28. Десенситизация метаботропных рецепторов включает три фазы:
- 29. Десенситизация метаботропных рецепторов При фосфорилировании протеин киназой
- 30. Десенситизация метаботропных рецепторов Вторая фаза заключается в
- 31. Десенситизация метаботропных рецепторов В третьей фазе интернализованный
- 32. Структура G-белков G-белки подразделяют на два типа
- 33. Структура G-белков G-белки являются гетеротримерами. G-белки
- 34. Функции G-белков-тримеров При связывании метаботропного рецептора с
- 35. Функции G-белков-мономеров G-белки мономеры гомологичны α-субъединицам гетеротримерных
- 36. Завершение процесса сигнализации G-белков определяется гидролизом
- 37. Подразделение G-белков-гетеротримеров Выделяют четыре группы по структуре
- 38. Подразделение G-белков-гетеротримеров Семейство Gi/Go-белков включает также
- 39. Подразделение G-белков-гетеротримеров Семейство Gi/Go-белков включает также
- 40. Подразделение G-белков-гетеротримеров Сигналы могут конвергировать (B, C)
- 41. Прямая модуляция активности ионных каналов G-белками Наряду
- 42. Прямая модуляция активности ионных каналов G-белками Прямое
- 43. Активация G-белками внутриклеточных метаболических каскадов Активированные G-белки
- 44. Пути внутриклеточной сигнализации, опосредованные G-белками первичный
- 45. Общая схема внутриклеточных метаболических каскадов с участием
- 46. Три уровня усиления в каскадах внутриклеточной сигнализации
- 47. Вторичные посредники В нейронах и многих других
- 48. Вторичные посредники: Са2+ является наиболее общим внутриклеточным
- 49. Вторичные посредники: Са2+ Одним из наиболее тщательно
- 50. Вторичные посредники: Са2+ поддержание внутриклеточной концентрации Высокий
- 51. Вторичные посредники: Са2+ источники Са2+-сигнала Са2+ поступает
- 52. Рианодиновый рецептор Рианодин – алкалоид растительного происхождения,
- 53. Рианодиновый рецептор The primary structure of a
- 54. Рианодиновый рецептор The primary structure of a
- 55. Са2+-токи рианодинового рецептора Modal gating of the
- 56. Вторичные посредники: Са2+ В цитоплазме Са2+ активирует
- 57. Вторичные посредники: цАМФ и цГМФ цАМФ синтезируется
- 58. Синтез и деградация цАМФ и цГМФ
- 59. Вторичные посредники: цАМФ и цГМФ При увеличении
- 60. Активация Са2+-каналов β-адренорецепторами Регуляция адренорецептором состояния
Слайд 1Метаботропные рецепторы
Метаботропные рецепторы представляют собой встроенные в мембрану отдельные белки, которые

Слайд 2Мембранные рецепторы, активируемые медиаторами
По механизму активации различных процессов (в том числе
и изменения ионной проводимости) мембранные рецепторы подразделяют на
ионотропные и метаботропные.
ионотропные и метаботропные.
")
Слайд 3Метаботропные рецепторы
В настоящее время в геноме млекопитающих идентифицировано более 1000 генов,
кодирующих молекулы метаботропных рецепторов.
По гомологии кодирующих генных последовательностей у млекопитающих выделяют три «главных» подсемейства:
1) родопсинподобные
2) секретинподобные,
3) метаботропные глутаматные рецепторы.
См. далее таблицу
По гомологии кодирующих генных последовательностей у млекопитающих выделяют три «главных» подсемейства:
1) родопсинподобные
2) секретинподобные,
3) метаботропные глутаматные рецепторы.
См. далее таблицу


Слайд 5The main five GPCR families:
Glutamate (G),
Rhodopsin (R),
Adhesion (A),
Frizzled (F)
Secretin (S)
Two independent receptor families:
Taste 2 receptors (T2Rs)
Vomeronasal 1 receptors (V1Rs)
,Rhodopsin (R),Adhesion (A),Frizzled (F) Secretin (S) Two independent receptor families:Taste")

Слайд 7Метаботропные рецепторы
Рецепторы этих подсемейств не демонстрируют значимого сходства между собой, однако
имеют сходное строение.
Кроме классических медиаторов некоторые метаботропные рецепторы активируются также
протонами,
Са2+,
аденозином,
АТФ,
простагландинами и другими липидами,
опиоидами и другими пептидами,
разнообразными белками,
светом,
различными одорантами
и вкусовыми веществами.
Кроме классических медиаторов некоторые метаботропные рецепторы активируются также
протонами,
Са2+,
аденозином,
АТФ,
простагландинами и другими липидами,
опиоидами и другими пептидами,
разнообразными белками,
светом,
различными одорантами
и вкусовыми веществами.

Слайд 8Эволюционное родство родопсинподобных рецепторов
To assemble this tree, sequence homologies in the
transmembrane domains were compared for each receptor. Distance determines the degree of relatedness.
r, rat; d, dog; h, human; tur, turkey; SRL, a putative serotonin receptor; and 5-HT, 5-hydroxytryptamine (serotonin).
Adapted from Linden (1994). Original tree construction was by William Pearson and Kevin Lynch, University of Virginia
r, rat; d, dog; h, human; tur, turkey; SRL, a putative serotonin receptor; and 5-HT, 5-hydroxytryptamine (serotonin).
Adapted from Linden (1994). Original tree construction was by William Pearson and Kevin Lynch, University of Virginia

Слайд 9Родопсинподобные метаботропные рецепторы
Acetylcholine (M1, M2, M3, M4, M5) - Adrenergic (α1
(A, B, D), α2 (A, B, C), β1, β2, β3) - Adrenomedullin - Anaphylatoxin (C3a, C5a) - Angiotensin (1, 2) - Apelin - Bile acid - Bombesin (BRS3, GRPR, NMBR) - Bradykinin (B1, B2) - Cannabinoid (CB1, CB2) - Chemokine - Cholecystokinin (A, B) - Dopamine (D1, D2, D3, D4, D5) - Eicosanoid (CysLT (1, 2), LTB4 (1, 2), FPRL1, OXE, Prostaglandin ((DP (1, 2), EP (1, 2, 3, 4), PGF, Prostacyclin, Thromboxane) - Endothelin (A, B) - Estrogen - Formyl peptide (1, L1, L2) - Free fatty acid (1, 2, 3, 4) - FSH - Galanin (1, 2, 3) - Gonadotropin-releasing hormone (1, 2) - GPR (1, 3, 4, 6, 12, 15, 17, 18, 19, 20, 21, 22, 23, 25, 26, 27, 31, 32, 33, 34, 35, 37, 39, 42, 45, 50, 52, 55, 61, 62, 63, 65, 68, 75, 78, 79, 82, 83, 84, 87, 88, 92, 101, 119, 120, 132, 135, 139, 141, 142, 146, 148, 149, 150, 151, 152, 153, 160, 161, 162, 171, 172, 173, 174, 182) - Ghrelin - Histamine (H1, H2, H3, H4) - Kisspeptin - Luteinizing hormone/choriogonadotropin - Lysophospholipid (1, 2, 3, 4, 5, 6, 7, 8) - MAS (1, 1L) - Melanocortin (1, 2, 3, 4, 5) - MCHR (1, 2) - Melatonin (1A, 1B)- Motilin - Neuromedin (B, U (1, 2)) - Neuropeptide (B/W (1, 2), FF (1, 2), S, Y (1, 2, 4, 5)) - Neurotensin (1, 2) - Opioid (Delta, Kappa, Mu, Nociceptin, but not Sigma) - Olfactory - Opsin (1SW) - Orexin (1, 2) - Oxytocin - PAF - Prokineticin (1, 2) - Prolactin-releasing peptide - Protease-activated (1, 2, 3, 4) - Purinergics (Adenosine (A1, A2a, A2b, A3), P2Y - Relaxin (1, 2, 3, 4) - Somatostatin (1, 2, 3, 4, 5) - Serotonin (5-HT1 (A, B, D, E, F), 5-HT2 (A, B, C), 5-HT (4, 5A, 6, 7) - SREB - TAAR (1, 2, 3, 5, 6, 8, 9) - Tachykinin (1, 2, 3) - Thyrotropin - Thyrotropin-releasing hormone - Urotensin-II - Vasopressin (1A, 1B, 2)
 - Adrenergic (α1 (A, B, D), α2")
Слайд 10Секретинподобные метаботропные рецепторы
Brain-specific angiogenesis inhibitor (1, 2, 3)
Cadherin (1, 2, 3)
Calcitonin
- CD97
Corticotropin-releasing hormone (1, 2)
EMR (1, 2, 3)
Glucagon (GR, GIPR, GLP1R, GLP2R)
Growth hormone releasing hormone
PACAPR1
GPR (56, 64, 97, 98, 110, 111, 112, 113, 114, 115, 116, 123, 124, 125, 126, 128, 133, 143, 144, 157)
Parathyroid hormone (1, 2)
Secretin
Vasoactive intestinal peptide (1, 2)
Corticotropin-releasing hormone (1, 2)
EMR (1, 2, 3)
Glucagon (GR, GIPR, GLP1R, GLP2R)
Growth hormone releasing hormone
PACAPR1
GPR (56, 64, 97, 98, 110, 111, 112, 113, 114, 115, 116, 123, 124, 125, 126, 128, 133, 143, 144, 157)
Parathyroid hormone (1, 2)
Secretin
Vasoactive intestinal peptide (1, 2)
Cadherin (1, 2, 3)Calcitonin - CD97Corticotropin-releasing hormone (1,")
Слайд 11Метаботропные глутаматные рецепторы
Calcium-sensing receptor
GABA B (1, 2)
Glutamate receptor (1, 2, 3,
4, 5, 6, 7, 8)
GPR (156, 158, 179) – orphan* G-protein-coupled receptors
RAIG (1, 2. 3. 4) - retinoic Acid-Inducible orphan G-protein-coupled receptors
Taste receptors (TAS1R, TAS2R)
GPR (156, 158, 179) – orphan* G-protein-coupled receptors
RAIG (1, 2. 3. 4) - retinoic Acid-Inducible orphan G-protein-coupled receptors
Taste receptors (TAS1R, TAS2R)
* An orphan receptor is an apparent receptor that has a similar structure to other identified receptors but whose endogenous ligand has not yet been identified. If a ligand for an orphan receptor is later discovered, the receptor is referred to as an "adopted orphan”
Glutamate receptor (1, 2, 3, 4, 5, 6, 7,")
Слайд 12Метаботропные рецепторы
Первым метаботропным рецептором, который был клонирован в начале 1990-х г.г.
и всесторонне изучен методами молекулярной биологии и классической фармакологии с широким использованием радиоактивных меченых лигандов, оказался β–адренорецептор.
В 2000 г. методом рентгеновской кристаллографии с высоким разрешением была получена трехмерная конфигурация родопсина, прототипа семейства родопсинподобных рецепторов, включающего β–адренорецептор.
В 2000 г. методом рентгеновской кристаллографии с высоким разрешением была получена трехмерная конфигурация родопсина, прототипа семейства родопсинподобных рецепторов, включающего β–адренорецептор.
Фосфорилирование нескольких оснований на третьей цитоплаз-матической петле и на карбоксиль-ном конце (например, цАМФ-зави- симой протеин киназой) вызывает десенситизацию рецептора.
Фосфорилирование карбоксильного конца (например, β-адрено-рецеп-тор-киназой) также приводит к десенситизации рецептора.

Слайд 13Семейства
метаботропных рецепторов
Красными кружками отмечены характерные консервативные остатки аминокислот трансмембранных сегментов, внутри-
и внеклеточных петель.
a) родопсинподобные
b) секретинподобные,
c) метаботропные глутаматные рецепторы
a) родопсинподобные
b) секретинподобные,
c) метаботропные глутаматные рецепторы
")
Слайд 14Родопсинподобные метаботропные рецепторы
наиболее изучены и включают родопсин, адренергические рецепторы, мускариновые ацетилхолиновые
(мАцХР), пуриновые, серотониновые рецепторы, а также многочисленные рецепторы пептидов и гормонов, подразделяемые на 19 подсемейств.
Относительно мелкие лиганды, такие как катехоламины, связываются в углублении, сформированном семью трансмембранными сегментами, а короткие пептиды взаимодействуют с внеклеточными петлями и N-терминалью, а также с трансмембранными сегментами.
Относительно мелкие лиганды, такие как катехоламины, связываются в углублении, сформированном семью трансмембранными сегментами, а короткие пептиды взаимодействуют с внеклеточными петлями и N-терминалью, а также с трансмембранными сегментами.
Цистеиновый остаток на участке С-терминали, примыкающем к ТМ7, обеспечивает привязку рецептора к мембране.
Дисульфидный мостик между цистеиновыми остатками внеклеточных петель между ТМ2-ТМ3 и ТМ4-ТМ5 сегментами является характерным для большинства рецепторов этого семейства.
, пуриновые, серотониновые рецепторы,")
Слайд 15Родопсинподобные метаботропные рецепторы
Цистеиновый остаток на участке С-терминали, примыкающем к ТМ7, обеспечивает
привязку рецептора к мембране.
Дисульфидный мостик между цистеиновыми остатками внеклеточных петель между ТМ2-ТМ3 и ТМ4-ТМ5 сегментами является характерным для большинства рецепторов этого семейства.
Дисульфидный мостик между цистеиновыми остатками внеклеточных петель между ТМ2-ТМ3 и ТМ4-ТМ5 сегментами является характерным для большинства рецепторов этого семейства.

Слайд 16Родопсинподобные метаботропные рецепторы
Amino acid sequence and predicted domain topology of the
M3 isoform of mAChR. The NH2 terminus of the protein is at the left and extends into the extracellular space. The COOH terminus is intracellular and is at the right; i1 to i4 are the four intracellular domains. The conserved disulfide bond (-S-S-) connects extracellular loop 2 to 3. Dashes in the amino acid sequence represent inserts of various lengths that are not shown. Conserved amino acids for all members of the G-protein-coupled receptor family of receptors are marked in purple. Amino acids taking part in ACh binding to the receptor are highlighted in yellow. Note that all amino acids associated with ligand binding lie in approximately the same horizontal plane across the receptor.

Слайд 17Секретинподобные метаботропные рецепторы
связываются с секретином, кальцитонином, паратироидными гормонами, глюкагоном, кортикотропин-релизинг фактором,
вазоактивными интестициальными пептидами и гипофизарными белками, активирующими аденилатциклазу.
Лиганд-связывающие участки этих рецепторов локализованы на внеклеточных доменах, а их относительно длинная N-терминаль включает несколько цистеиновых остатков, которые образуют сеть дисульфидных мостиков.
Лиганд-связывающие участки этих рецепторов локализованы на внеклеточных доменах, а их относительно длинная N-терминаль включает несколько цистеиновых остатков, которые образуют сеть дисульфидных мостиков.

Слайд 18Глютаматные метаботропные рецепторы
включают восемь типов метаботропных рецепторов (mGluR), Са2+-чувствитель-ные рецепторы и
ГАМКВ рецепторы.
Характеризуются длинными N- и С-терминалями. Лиганд-связывающий участок у mGluR локализован на N-терминалях двух субъединиц рецептора, которые связаны между собой дисульфидным мостиком.
Два цистеиновых остатка на внеклеточных петлях образуют дисульфидный мостик. Уникальной особенностью этого семейства рецепторов является короткая и высоко консервативная внутриклеточная петля ТМ5-ТМ6.
Характеризуются длинными N- и С-терминалями. Лиганд-связывающий участок у mGluR локализован на N-терминалях двух субъединиц рецептора, которые связаны между собой дисульфидным мостиком.
Два цистеиновых остатка на внеклеточных петлях образуют дисульфидный мостик. Уникальной особенностью этого семейства рецепторов является короткая и высоко консервативная внутриклеточная петля ТМ5-ТМ6.
, Са2+-чувствитель-ные рецепторы и ГАМКВ рецепторы.Характеризуются длинными N-")
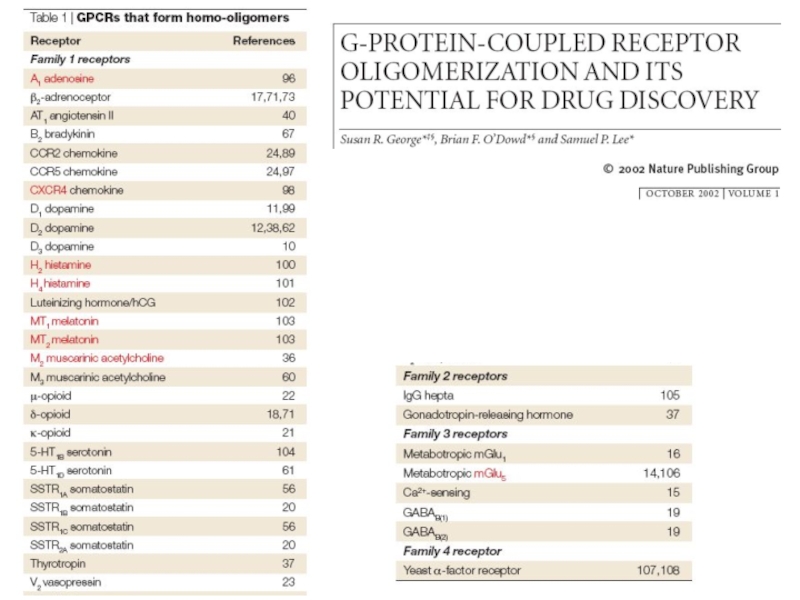
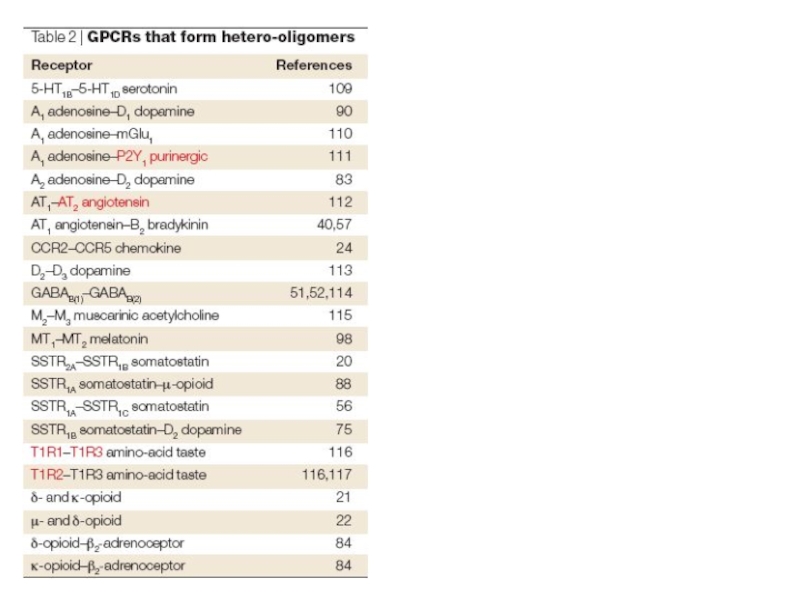
Слайд 19Олигомеризация метаботропных рецепторов
Метаботропные рецепторы являются олигомерами.
Пример рецептора (гетеро)димера
ГАМКВ рецептор состоит из
двух субъединиц GABABR1 и GABABR2.
Субъединица-мономер GABABR1 локализована в ЭПР (его удерживает С-терминаль).
Субъединица-мономер GABABR1 локализована в ЭПР (его удерживает С-терминаль).
Субъединица-мономер GABABR2 экспрессирована в клеточной мембране, но в мономерной конфигурации не функциональна (не связывается с агонистом).
Гетеродимер GABABR1/GABABR2 экспрессирован в клеточной мембране и является функциональным (способен связываться с агонистом).
димераГАМКВ рецептор состоит из двух субъединиц GABABR1 и")
Слайд 20Рецептор-олигомер представлен комплексом из нескольких рецепторов-субъединиц, связанных ковалентными мостиками или межмолекулярными
связями.
Субъединица, входящая в состав комплекса, называется протомером. Субъединица вне комплекса называется мономером.
Рецепторы-гомо(олиго)меры состоят из идентичных протомеров,
рецепторы-гетеро(олиго)меры состоят из разных протомеров.
Субъединица, входящая в состав комплекса, называется протомером. Субъединица вне комплекса называется мономером.
Рецепторы-гомо(олиго)меры состоят из идентичных протомеров,
рецепторы-гетеро(олиго)меры состоят из разных протомеров.
Олигомеризация метаботропных рецепторов

Слайд 21Димеризация метаботропных рецепторов
Some GPCRs can function as dimers.
In this example, chimeric
receptors were produced between α2AR (α2) and mAChR (M3) by swapping certain transmembrane domains through genetic engineering.
When α2/M3 or M3/α2 is expressed separately, they are not active. However, if both chimeric molecules are expressed in the same cells, they form receptors that can be activated by either epinephrine or muscarine.
Adapted from Salahpour et al., (2000).
A top-down view of how this domain swapping might occur when two molecules dimerize to produce receptors that can respond to both transmitters. Adapted from Lee et al., (2000).
When α2/M3 or M3/α2 is expressed separately, they are not active. However, if both chimeric molecules are expressed in the same cells, they form receptors that can be activated by either epinephrine or muscarine.
Adapted from Salahpour et al., (2000).
A top-down view of how this domain swapping might occur when two molecules dimerize to produce receptors that can respond to both transmitters. Adapted from Lee et al., (2000).


Слайд 23When 5-HT2A and mGlu2 are prevented from forming a receptor heterocomplex,
activation of 5-HT2A by LSD elicits signaling characteristic of Gq/11-protein subtypes. In contrast, LSD acting at the 5-HT2A-mGlu2 receptor heterocomplex activates both Gq/11- and Gi/o-dependent signaling. Head-twitch behavior is reliably and robustly elicited by hallucinogenic 5-HT2A agonists, and is absent in mGlu2 knockout mice.
Димеризация метаботропных рецепторов
G protein-dependent signaling and behavioral responses that require a GPCR heterocomplex. Hallucinogenic drugs, such as lysergic acid diethylamide (LSD), mescaline and psilocybin, all have in common a high affinity for the serotonin 5-HT2A receptor. Metabotropic glutamate 2 receptor (mGlu2) and serotonin 5-HT2A receptor form a specific functional GPCR heterocomplex in mammalian brain and in tissue culture preparations. mGlu2 transmembrane domains 4/5 mediate association with the 5-HT2A receptor.
судорожное движение головой

Слайд 26a. Schematic representation of ligand-induced heterodimerization by a bivalent peptide ligand.
A bivalent peptide with two distinct receptor-recognition sites might bind two different receptors simultaneously and induce receptor heterodimerization. If the two receptors pre-exist in an oligomeric state, the ligand might enhance the heteromeric conformation by crosslinking two receptors.
b. Dimeric ligands generated from two monovalent ligands. Similar to a bivalent ligand, a dimeric ligand that has an appropriate linker might induce or enhance dimerization of two receptors. It is plausible that homodimeric and heterodimeric ligands might selectively bind homomeric and heteromeric receptors, respectively.
b. Dimeric ligands generated from two monovalent ligands. Similar to a bivalent ligand, a dimeric ligand that has an appropriate linker might induce or enhance dimerization of two receptors. It is plausible that homodimeric and heterodimeric ligands might selectively bind homomeric and heteromeric receptors, respectively.

Слайд 27Десенситизация метаботропных рецепторов
Десенситизация рецепторов заключается в затухании эффекторной активности и прекращении
физиологического ответа, несмотря на то, что лиганд остается связанным с рецептором.
Описано два механизма десенситизации:
- через фосфорилирование
- физическое удаление рецептора из мембраны
Описано два механизма десенситизации:
- через фосфорилирование
- физическое удаление рецептора из мембраны

Слайд 28Десенситизация метаботропных рецепторов
включает три фазы:
Первая фаза десенситизации (от секунд до минут)
заключается в расцеплении рецептора и G-белка, что происходит в результате фосфорилирования рецептора протеин киназами С и А и G-белок-рецептор-киназой. Киназы активируются вторичными посредниками, которые синтезируются в результате функционирования рецепторов при их связывании с агонистами.
Рецептор может фосфорилироваться протеин киназой А даже если он не связан с агонистом (т.н. гетерогенная десенситизация).
Рецептор может фосфорилироваться протеин киназой А даже если он не связан с агонистом (т.н. гетерогенная десенситизация).
 заключается в расцеплении рецептора")
Слайд 29Десенситизация метаботропных рецепторов
При фосфорилировании протеин киназой А происходит расцепление рецептора и
G-белка.
При фосфорилировании G-белок-рецептор-киназой происходит связывание рецептора с белком аррестином, который устраняет связывание рецептора с G-белком.
При фосфорилировании G-белок-рецептор-киназой происходит связывание рецептора с белком аррестином, который устраняет связывание рецептора с G-белком.

Слайд 30Десенситизация
метаботропных рецепторов
Вторая фаза заключается в изоляции, или так называемой интернализации рецептора.
При фосфорилировании G-белок-рецептор-киназой происходит связывание рецептора с белком аррестином.
Затем рецепторы подвергаются эндоцитозу и удаляются с поверхности мембраны.
Затем рецепторы подвергаются эндоцитозу и удаляются с поверхности мембраны.

Слайд 31Десенситизация
метаботропных рецепторов
В третьей фазе интернализованный рецептор либо:
дефосфорилируется и возвращается (рециклируется)
на поверхность клеточной мембраны и в дальнейшем продолжает функционировать (рециклирование/ ресенситизация рецептора);
2) секвестируется (изолируется) в мембране эндосом, при дефосфорилировании освобождается от аррестина и рециклируется;
3) транспортируется в лизосомы, где происходит его деградация. При этом необходимо пополнение пула рецепторов путем их синтеза.
2) секвестируется (изолируется) в мембране эндосом, при дефосфорилировании освобождается от аррестина и рециклируется;
3) транспортируется в лизосомы, где происходит его деградация. При этом необходимо пополнение пула рецепторов путем их синтеза.
(1)
(2)
(3)
 на поверхность клеточной")
Слайд 32Структура G-белков
G-белки подразделяют на два типа – гетеротримеры и мономеры.
G-белки гетеротримеры
состоят из трех отдельных субъединиц (α, β и γ). Существует большое количество разновидностей каждой из субъединиц (20 α, 6 β и 12 γ), что создает основу для большого количества их комбинаций.
α-субъединица связывается с гуаниновыми нуклеотидами - либо с ГТФ, либо с ГДФ.
При связывании с ГДФ α-субъединица соединяется с β- и γ-субъединицами, формируя неактивный тример (αβγ).
α-субъединица связывается с гуаниновыми нуклеотидами - либо с ГТФ, либо с ГДФ.
При связывании с ГДФ α-субъединица соединяется с β- и γ-субъединицами, формируя неактивный тример (αβγ).

Слайд 33Структура G-белков
G-белки являются гетеротримерами.
G-белки состоят из трех отдельных субъединиц (α, β
и γ).
α-субъединица связывается с гуаниновыми нуклеотидами - либо с ГТФ, либо с ГДФ.
При связывании с ГДФ α-субъединица соединяется с β- и γ-субъединицами, формируя неактивный тример (αβγ).
GRK - G protein-coupled receptor kinases
α-субъединица связывается с гуаниновыми нуклеотидами - либо с ГТФ, либо с ГДФ.
При связывании с ГДФ α-субъединица соединяется с β- и γ-субъединицами, формируя неактивный тример (αβγ).
GRK - G protein-coupled receptor kinases
.α-субъединица связывается с")
Слайд 34Функции G-белков-тримеров
При связывании метаботропного рецептора с внеклеточным сигналом G-белок связывается с
рецептором, и в этом состоянии ГДФ на α-субъединице замещается на ГТФ.
В конфигурации с ГТФ (активированный G-белок) α-субъединица отделяется от димера βγ-субъединиц.
В конфигурации с ГТФ (активированный G-белок) α-субъединица отделяется от димера βγ-субъединиц.
Вслед за активацией α-субъединица, связанная с ГТФ, и комплекс из βγ-субъединиц могут связываться с молекулами-эффекторами следующего порядка, обуславливающими разнообразные ответы клеток-мишеней.
GAP - GTPase-Activating Proteins

Слайд 35Функции G-белков-мономеров
G-белки мономеры гомологичны α-субъединицам гетеротримерных G-белков и называются также малыми
G-белками
Передают сигнал от активируемых внеклеточными агентами метаботропных рецепторов цитоплазматическим мишеням, таким как элементы цитоскелета и системы везикулярного транспорта. Такие малые G-белки впервые были открыты в вирусах. Ras – от rat sarcoma.
Передают сигнал от активируемых внеклеточными агентами метаботропных рецепторов цитоплазматическим мишеням, таким как элементы цитоскелета и системы везикулярного транспорта. Такие малые G-белки впервые были открыты в вирусах. Ras – от rat sarcoma.
При связывании с ГТФ они активируют протеин киназы, которые передают сигнал в ядро. Это приводит к неконтролируемому росту клеток и развитию опухолей.
После этих открытий было идентифицировано большое число малых G-белков с разнообразными функциями. Например, некоторые из них задействованы в транспорте везикул в пресинаптических терминалях или в других частях нейронов. Другие играют главную роль в транспорте белков и РНК в ядро и из ядра.

Слайд 36Завершение процесса сигнализации G-белков
определяется гидролизом ГТФ в ГДФ у α-субъединицы.
Скорость
гидролиза ГТФ является важным свойством G-белков и регулируется ГТФаза-активируемыми белками (GAP, от GTPase-Activating Proteins). Активная α-субъединица, связанная с ГТФ, функционально является ГТФазой.
Замещая ГТФ на ГДФ, ГТФаза-активируемые белки возвращают α-субъединицу G-белков в неактивное состояние.
Замещая ГТФ на ГДФ, ГТФаза-активируемые белки возвращают α-субъединицу G-белков в неактивное состояние.
Сначала активируемые ГТФазой белки были открыты как регуляторы малых G-белков, затем их регулирующая функция была показана и для α-субъединиц G-белков-гетеротримеров.

Слайд 37Подразделение G-белков-гетеротримеров
Выделяют четыре группы по структуре и мишеням α-субъединиц:
1) Gs стимулирует
аденилатциклазу (или гуанилатциклазу);
2) Gi ингибирует аденилатциклазу (или гуанилатциклазу),
Go ингибирует потенциал-зависимые Са2+- и К+-каналы;
3) Gq/11 (или Gq) активирует фосфолипазу С;
4) G12/13 не участвует в процессах внутриклеточной сигнализации,
имеющих отношение к непрямой синаптической передаче.
Димер, образованный комплексом из β- и γ-субъединиц (Gβγ), также проявляет свою самостоятельную функцию, например, регулирует проводимость ионных каналов (например, Са2+-каналов).
2) Gi ингибирует аденилатциклазу (или гуанилатциклазу),
Go ингибирует потенциал-зависимые Са2+- и К+-каналы;
3) Gq/11 (или Gq) активирует фосфолипазу С;
4) G12/13 не участвует в процессах внутриклеточной сигнализации,
имеющих отношение к непрямой синаптической передаче.
Димер, образованный комплексом из β- и γ-субъединиц (Gβγ), также проявляет свою самостоятельную функцию, например, регулирует проводимость ионных каналов (например, Са2+-каналов).
 Gs стимулирует аденилатциклазу (или гуанилатциклазу);2) Gi")
Слайд 38Подразделение G-белков-гетеротримеров
Семейство Gi/Go-белков включает также
Gt (трансдуцин), который активирует фосфодиэстеразу, катализирующую превращение
цГМФ в нециклическую форму 5’3’-ГМФ в фоторецепторах сетчатки,
, который активирует фосфодиэстеразу, катализирующую превращение цГМФ в нециклическую форму")
Слайд 39Подразделение G-белков-гетеротримеров
Семейство Gi/Go-белков включает также
G-белок густодуцин (передатчик вкуса), который активирует фосфолипазу
С (на рис. PLCβ2) в мембранах вкусовых рецепторных клеток.
, который активирует фосфолипазу С (на рис. PLCβ2)")
Слайд 40Подразделение G-белков-гетеротримеров
Сигналы могут конвергировать (B, C) и дивергировать (D, E) в
зависимости от взаимодействия между рецепторами (Ri), G-белками (Gi) и эффекторами (Ei).
 и дивергировать (D, E) в зависимости от взаимодействия между")
Слайд 41Прямая модуляция активности ионных каналов G-белками
Наряду с активацией ферментных молекул-мишеней G-белки
могут также напрямую связываться с ионными каналами.
При активации мускаринового ацетилхолинового рецептора в кардиомиоцитах происходит диссоциация G-белка. Димер β/γ напрямую связывается с K+-каналом и открывает его, в результате чего выходящий ток катионов вызывает гиперполяризацию клетки.
В специальных экспериментах методом patch clamp на кардимиоцитах показано, что при аппликации β/γ с внутренней стороны мембраны происходит открытие K+-канала.
При активации мускаринового ацетилхолинового рецептора в кардиомиоцитах происходит диссоциация G-белка. Димер β/γ напрямую связывается с K+-каналом и открывает его, в результате чего выходящий ток катионов вызывает гиперполяризацию клетки.
В специальных экспериментах методом patch clamp на кардимиоцитах показано, что при аппликации β/γ с внутренней стороны мембраны происходит открытие K+-канала.

Слайд 42Прямая модуляция активности ионных каналов G-белками
Прямое взаимодействие G-белков и ионных каналов
показано в механизме ауторегуляции выделения норадреналина из нейронов симпатического ганглия лягушки. Выделяясь из аксонной терминали, норадреналин связывается со специальными пресинаптическими ауторецепторами.
В результате диссоциации G-белка димер β/γ напрямую действует на Ca2+-канал N-типа, снижая вероятность его открытия. Уменьшение Ca2+-тока в пресинаптическую терминаль приводит к уменьшению выделения медиатора.
В результате диссоциации G-белка димер β/γ напрямую действует на Ca2+-канал N-типа, снижая вероятность его открытия. Уменьшение Ca2+-тока в пресинаптическую терминаль приводит к уменьшению выделения медиатора.

Слайд 43Активация G-белками внутриклеточных метаболических каскадов
Активированные G-белки влияют на функции многих эффекторов
следующего порядка.
Большинство таких эффекторов являются ферментами, которые при их активации G-белками синтезируют внутриклеточные вторичные посредники, а при ингибировании – прекращают синтез последних.
Эти ферменты включают аденилат- и гунилатциклазы, фосфолипазу С и некоторые другие.
Вторичные посредники инициируют каскады следующего порядка.
Поскольку каждый из этих каскадов запускается специфическими субъединицами G-белков, пути внутриклеточной сигнализации, опосредованные отдельными рецепторами, зависят от специфичности ассоциированных с этими рецепторами субъединиц G-белков.
Большинство таких эффекторов являются ферментами, которые при их активации G-белками синтезируют внутриклеточные вторичные посредники, а при ингибировании – прекращают синтез последних.
Эти ферменты включают аденилат- и гунилатциклазы, фосфолипазу С и некоторые другие.
Вторичные посредники инициируют каскады следующего порядка.
Поскольку каждый из этих каскадов запускается специфическими субъединицами G-белков, пути внутриклеточной сигнализации, опосредованные отдельными рецепторами, зависят от специфичности ассоциированных с этими рецепторами субъединиц G-белков.

Слайд 44Пути внутриклеточной сигнализации, опосредованные G-белками
первичный посредник (медиатор)
рецептор
G-белок
эффекторный белок (фермент)
вторичный посредник
протеин-киназа
целевое действие
рецепторG-белокэффекторный белок (фермент)вторичный посредник протеин-киназа целевое действие")
Слайд 45Общая схема внутриклеточных метаболических каскадов с участием G-белков состоит из следующих
этапов:
В результате активации рецептора ГДФ на α-субъединице G-белка (передатчик) замещается на ГТФ, что приводит к диссоциации α- и βγ-субъединиц.
Различные типы активированной α-субъединицы взаимодействуют с рядом первичных эффекторов - ферментов (циклазы, фосфолипазы, фосфодиэстеразы цГМФ, ГТФаза-активируемые белки и некоторые другие). В результате взаимодействия α-субъединица может либо активировать ферменты, что приводит к синтезу (циклазы) или деградации (фосфодиэстеразы) вторичных посредников, либо ингибировать ферменты (циклазы), прекращая синтез вторичных посредников. Димер βγ ингибирует и стимулирует (вместе с α-субъединицей) некоторые аденилатциклазы, а также регулирует активацию фосфолипаз Сβ и А2, К+- и Са2+-каналов и других эффекторов.
Вторичные посредники активируют различные белки-мишени (вторичные эффекторы) – протеинкиназы, фосфатазы, липоксигеназы, циклооксигеназы и др., воздействуя на их регуляторные субъединицы.
Каталические субъединицы различных белков-мишеней вызывают дальнейшие эффекты. Например, протеин киназы фосфорилируют канальные белки, что приводит к открытию ионных каналов, а фосфатазы дефосфорилируют канальные белки, что приводит к закрытию ионных каналов
Посредники следующего уровня либо активируют различные третичные эффекторы, либо влияют на ионную проводимость в мембранах, в том числе и в мембранах клеточных органелл.


Слайд 47Вторичные посредники
В нейронах и многих других клетках представлены многие вторичные посредники
как внутриклеточные сигнальные молекулы. Эти посредники различаются по механизмам их синтеза и деградации, а также по их молекулам-мишеням следующего порядка и эффектам.

Слайд 48Вторичные посредники: Са2+
является наиболее общим внутриклеточным посредником в нейронах и других
клетках.
Во всех случаях при участии Са2+ информация передается в результате быстрого возрастания его концентрации в цитоплазме (от 10-100 нМ до 0,5-1,0 мкМ), в результате чего Са2+ связывается с большим числом Са2+-связывающих белков, которые выступают в качестве молекул-мишеней.
Внутриклеточный Са2+ регулирует К+- и Cl--каналы, а также активность связанных с мембраной фосфолипаз С и А2.
В цитоплазме Са2+ активирует три основных мишени:
- протеинкиназу С,
- калмодулин
- и Са2+-зависимую протеазу (кальпаин).
Во всех случаях при участии Са2+ информация передается в результате быстрого возрастания его концентрации в цитоплазме (от 10-100 нМ до 0,5-1,0 мкМ), в результате чего Са2+ связывается с большим числом Са2+-связывающих белков, которые выступают в качестве молекул-мишеней.
Внутриклеточный Са2+ регулирует К+- и Cl--каналы, а также активность связанных с мембраной фосфолипаз С и А2.
В цитоплазме Са2+ активирует три основных мишени:
- протеинкиназу С,
- калмодулин
- и Са2+-зависимую протеазу (кальпаин).

Слайд 49Вторичные посредники: Са2+
Одним из наиболее тщательно изученных белков-мишеней Са2+ является белок
калмодулин, который содержится в цитозоле в большой концентрации.
При связывании с Са2+ этот белок активируется, и комплекс Са2+/калмодулин инициирует разнообразные эффекты, активируя эффекторы следующего порядка:
Са2+/калмодулин-зависимую протеин киназу II (СаМК II),
аденилатциклазу,
фосфодиэстеразу циклических нуклеотидов,
протеин фосфатазу (кальцинейрин)
и NO-синтазу.
При связывании с Са2+ этот белок активируется, и комплекс Са2+/калмодулин инициирует разнообразные эффекты, активируя эффекторы следующего порядка:
Са2+/калмодулин-зависимую протеин киназу II (СаМК II),
аденилатциклазу,
фосфодиэстеразу циклических нуклеотидов,
протеин фосфатазу (кальцинейрин)
и NO-синтазу.

Слайд 50Вторичные посредники: Са2+
поддержание внутриклеточной концентрации
Высокий градиент Са2+ поддерживается системами активного транспорта
(Са2+-насосом и Na+/Са2+-обменником).
Цитоплазматический Са2+ закачивается специальными Са2+-насосами в эндоплазматический ретикулум и митохондрии. Эти органеллы выступают в роли внутриклеточных Са2+-депо.
В клетках присутствует Са2+-связывающий белок калбиндин (от англ., Са2+-binding protein), который выступает в качестве Са2+-буфера.
Такие буферные системы, обратимо связывающие Са2+, демпфируют амплитуду и кинетику Са2+-сигнала внутри нейронов.
Цитоплазматический Са2+ закачивается специальными Са2+-насосами в эндоплазматический ретикулум и митохондрии. Эти органеллы выступают в роли внутриклеточных Са2+-депо.
В клетках присутствует Са2+-связывающий белок калбиндин (от англ., Са2+-binding protein), который выступает в качестве Са2+-буфера.
Такие буферные системы, обратимо связывающие Са2+, демпфируют амплитуду и кинетику Са2+-сигнала внутри нейронов.
.Цитоплазматический")
Слайд 51Вторичные посредники: Са2+
источники Са2+-сигнала
Са2+ поступает в клетку из внеклеточной среды по
потенциал- и лиганд-зависимым Са2+-каналам плазматической мембраны.
Са2+ высвобождается в цитозоль из цистерн эндоплазматического ретикулума. Эти каналы активируются в ответ на различные внутриклеточные сигналы:
- Инозитол-трифосфатный (ИФ3-) рецептор. ИФ3 является вторичным посредником, синтезируемым в результате активации метаботропных рецепторов.
- Рианодиновый рецептор. Рианодин является экзогенным лигандом, который связывается с этим рецептором и частично открывает Са2+-канал. Этот рецептор активируется внутриклеточным Са2+, а в мышечных клетках – при деполяризации плазматической мембраны.
Са2+ высвобождается в цитозоль из цистерн эндоплазматического ретикулума. Эти каналы активируются в ответ на различные внутриклеточные сигналы:
- Инозитол-трифосфатный (ИФ3-) рецептор. ИФ3 является вторичным посредником, синтезируемым в результате активации метаботропных рецепторов.
- Рианодиновый рецептор. Рианодин является экзогенным лигандом, который связывается с этим рецептором и частично открывает Са2+-канал. Этот рецептор активируется внутриклеточным Са2+, а в мышечных клетках – при деполяризации плазматической мембраны.

Слайд 52Рианодиновый рецептор
Рианодин – алкалоид растительного происхождения, который связывается с этим рецептором
и частично открывает Са2+-канал.
Этот рецептор активируется внутриклеточным Са2+ (положительная обратная связь).
В сердце и поджелудочной железе активируется также другим вторичным посредником - циклической АДФ-рибозой (Cyclic ADP-ribose)
Этот рецептор активируется внутриклеточным Са2+ (положительная обратная связь).
В сердце и поджелудочной железе активируется также другим вторичным посредником - циклической АДФ-рибозой (Cyclic ADP-ribose)

Слайд 53Рианодиновый рецептор
The primary structure of a cardiac ryanodine receptor and binding
domains of protein phosphatases 1 and 2A, protein kinase A, calmodulin and FKBP12.6 are indicated. Protein phosphatases 1 and 2A and protein kinase A bind to the cardiac ryanodine receptor via their specific adaptor proteins. Three major divergent (nonhomologous) regions are also indicated. Six transmembrane segments are shown as previously proposed.
CaM, calmodulin; DR, divergent region; FKBP, calstabin-2; LIZ, leucine–isoleucine zipper; PKA, protein kinase A; PP, protein phosphatase; SR, sarcoplasmic reticulum.
CaM, calmodulin; DR, divergent region; FKBP, calstabin-2; LIZ, leucine–isoleucine zipper; PKA, protein kinase A; PP, protein phosphatase; SR, sarcoplasmic reticulum.
Yano M et al. (2006) Mechanisms of Disease: ryanodine receptor defects in heart failure and fatal arrhythmia Nat Clin Pract Cardiovasc Med 3: 43–52

Слайд 54Рианодиновый рецептор
The primary structure of a ryanodine receptor
6 трансмембранных сегментов представляют
Са2+-канал (ионная пора – между 5 и 6-м).

Слайд 55Са2+-токи рианодинового рецептора
Modal gating of the RyR2 channel. The Markovian scheme
shown in Figure 3 was used here to simulate the single-channel recordings shown. At the top, a series of high Po bursts or low Po bursts are marked. Trains of high or low Po bursts correspond to the high or low Po gating modes of the RyR2 channel.
At any steady-state calcium concentration, the RyR2 channel will spontaneously shift between its different gating modes. A sample spontaneous modal gating shift (at pCa 5) is shown in the middle panel. In this case, the channel shifted from the low Po mode to the high Po mode and back. Shifts in model RyR2 gating can also be evoked by fast calcium concentration changes. A sample evoked modal gating shift is shown on the bottom panel. A fast calcium concentration change (pCa 7 to 6) was introduced at the arrow. The channel immediately shifts into the high Po mode for a period of time before shifting again into the low Po mode.
At any steady-state calcium concentration, the RyR2 channel will spontaneously shift between its different gating modes. A sample spontaneous modal gating shift (at pCa 5) is shown in the middle panel. In this case, the channel shifted from the low Po mode to the high Po mode and back. Shifts in model RyR2 gating can also be evoked by fast calcium concentration changes. A sample evoked modal gating shift is shown on the bottom panel. A fast calcium concentration change (pCa 7 to 6) was introduced at the arrow. The channel immediately shifts into the high Po mode for a period of time before shifting again into the low Po mode.

Слайд 56Вторичные посредники: Са2+
В цитоплазме Са2+ активирует три основных мишени:
протеинкиназу С,
- калмодулин и Са2+-зависимую протеазу (кальпаин).
Са2+/калмодулин активирует:
Са2+/калмодулин-зависимую протеин киназу II, аденилатциклазу, фосфодиэстеразу циклических нуклеотидов, протеин фосфатазу (кальцинейрин) и NO-синтазу
Са2+/калмодулин активирует:
Са2+/калмодулин-зависимую протеин киназу II, аденилатциклазу, фосфодиэстеразу циклических нуклеотидов, протеин фосфатазу (кальцинейрин) и NO-синтазу
фосфатаза
протеаза

Слайд 57Вторичные посредники: цАМФ и цГМФ
цАМФ синтезируется из АТФ при активации аденилатциклазы,
локализованной в плазматической мембране. цГМФ синтезируется из ГТФ при активации гуанилатциклазы.
Циклические нуклеотиды деградируют в результате активации фосфодиэстераз, которые разрезают фосфодиэфирную связь в молекулах цАМФ и цГМФ и превращает их в линейную форму 3’5’-АМФ и 3’5’-ГМФ.
Уровень циклических нуклеотидов регулируется балансом их синтеза (циклазами) и деградации (фосфодиэстеразами). Каждый из этих ферментов может регулироваться независимо.
Циклические нуклеотиды деградируют в результате активации фосфодиэстераз, которые разрезают фосфодиэфирную связь в молекулах цАМФ и цГМФ и превращает их в линейную форму 3’5’-АМФ и 3’5’-ГМФ.
Уровень циклических нуклеотидов регулируется балансом их синтеза (циклазами) и деградации (фосфодиэстеразами). Каждый из этих ферментов может регулироваться независимо.


Слайд 59Вторичные посредники: цАМФ и цГМФ
При увеличении концентрации циклические нуклеотиды связываются с
двумя различными классами внутриклеточных мишеней.
Типичными их мишенями являются протеин киназы – цАМФ-зависимая протеин киназа (ПКА) и цГМФ-зависимая протеинкиназа (ПКГ).
Протеин киназы обеспечивают множество физиологических ответов, фосфорилируя различные мишени белки-мишени.
Циклические нуклеотиды могут связываться с цитоплазматическими участками С-терминалей определенных лиганд-зависимых ионных каналов, а также с цАМФ- потенциал-зависимыми неселективными катионными каналами.
Типичными их мишенями являются протеин киназы – цАМФ-зависимая протеин киназа (ПКА) и цГМФ-зависимая протеинкиназа (ПКГ).
Протеин киназы обеспечивают множество физиологических ответов, фосфорилируя различные мишени белки-мишени.
Циклические нуклеотиды могут связываться с цитоплазматическими участками С-терминалей определенных лиганд-зависимых ионных каналов, а также с цАМФ- потенциал-зависимыми неселективными катионными каналами.

Слайд 60Активация Са2+-каналов β-адренорецепторами
Регуляция адренорецептором состояния Са2+-каналов в кардиомиоцитах.
Связывание норадреналина (NE)
с β-адренорецептором приводит к активации Gs-белка, который диссоциируется на α- и βγ-субъединицы. Обе субъединицы связываются с аденилатциклазой, активируя ее. Аденилатциклаза продуцирует цАМФ, которая активирует цАМФ-зависимую протеин киназу. Протеин киназа фосфорилирует потенциал-зависимые Са2+-каналы, устраняя их инактивацию.
 с β-адренорецептором приводит к")





