- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Биотехнология растений. Трансгенные растения (часть 4) презентация
Содержание
- 1. Биотехнология растений. Трансгенные растения (часть 4)
- 2. Лекция 6: Трансгенные растения-4 Получение инсерционных мутантов
- 3. Т-ДНК инсерционный мутагенез интеграция Т-ДНК изменение
- 4. Схемы используемых плазмид для получения: рецессивных мутаций
- 5. Клонирование генов
- 6. Клонирование гена agamous с помощью Т-ДНК инсерционного
- 7. Ген, маркированный транспозоном (tagged gene), можно клонировать с помощью «вытягивания за транспозон»
- 8. Транспозонный мутагенез интеграция
- 9. При встраивании транспозона в кодирующую последовательность гена, наблюдается мутантный фенотип. При вырезании фенотип восстанавливается
- 10. Клонирование генов на основе транспозонного (инсерционного) мутагенеза
- 11. Клонирование генов на основе транспозонного (инсерционного) мутагенеза
- 12. Получение мутации flo613 и клонирование гена FLORICAULA
- 13. Рестрикционный анализ ДНК из выделенных клонов,
- 14. Создание геномной библиотеки из мутанта flo 613, выращиваемого при tº=25ºC (Тam3 не перемещается)
- 15. Скрининг библиотеки с меченой пробой из Tam3
- 16. Д.т. flo
- 17. Ген FLORICAULA – уникальный ген, присутствующий в
- 18. Изучение механизмов регуляции экспрессии
- 19. Слитые гены-репортеры К промотору исследуемого гена присоединяют репортерный ген
- 20. Green fluorescent protein (GFP) Luciferase (LUC) ß-glucuronidase
- 21. В трансформированных растениях проводят анализ экспрессии репротерного гена, отбирают трансформантов с искомым характером экспрессии
- 22. Анализ распределение ауксина в растениях с помощью генетической конструкции DR5:GUS DR5 – синтетический ауксин-регулируемый промотор
- 23. Анализ активности промоторов ключевых регуляторных генов в меристемах растений
- 24. Получение растений со сверхэкспрессией гена с целью изучения функции гена
- 25. Фенотип трансгенных растений 35S::LFY У растений 35S::LFY
- 26. Характер экспрессии генов-ортологов LFY/FLO может определять тип соцветия (открытое или закрытое)
- 27. Метод позволяет изучить регуляторную функцию любого участка
- 28. Мутант арабидопсиса по гену LFY
- 29. Основные семейства транскрипционных факторов растений
- 30. Гомеодомен-ДНК - комплекс вариабельный уч.
- 32. ТФ с гомеодоменами (семейство TALE) KNOX мишени:
- 33. Ген KNOTTED1 был идентифи-цирован на основе доминантных
- 34. Зная о важной роли гомеобоксных генов в
- 35. Сверхэкспрессия гена KNAT1 у арабидопсиса На листьях
- 36. Сверхэкспрессия гена KNAT в листьях томатов дикий тип 35S::KNAT
- 37. KNOTTED-подобные гены – возможные участники морфологической эволюции растений
- 38. Обнаружение связи между уровнем экспрессии KNOX-генов и
- 39. MCM1 – регулятор транскрипции дрожжей AG –
- 40. ТФ с MADS доменом вышележащие регуляторы: ТФ
- 41. Исследования трансгенных растений с конститутивной экспрессией гомеозисных
- 42. Цветки с двойным венчиком (лилейные) Цветки с
- 43. Использование методов «генетической хирургии» для изучения взаимодействия клеток и тканей в процессах развития
- 44. Day, Galgoci, Irish, 1995. Генетическое удаление
- 45. «Пространственная» модель Holder, 1979. Под влиянием
- 46. В основе «генетической хирургии» лежит метод трансформации
- 47. гены, кодирующие рибонуклеазы: Т1 рибонуклеазу Aspergillus oryzae
- 48. Целью работы Day, Galgoci, Irish (1995) была
- 49. Этапы работы Слитые гены включали в Т-ДНК
- 50. У резушки получено 5 трансгенных растений, в
- 51. Использование трансформации для изучения роли гормонов в развитиии растений
- 53. Корончатый галл на растении табака как результат трансформации агробактерией (штамм дикого типа)
- 54. Трансгенные рстения с геном tmr
- 55. Ген tmr в трансгенных растениях, экспрессирующийся в плодах (промотор 2А11)
- 56. Опухолеобразование у редиса – генетический признак
- 57. Фенокопии опухолеобразования у трансгенных растений с геном ipt
- 58. Прямая генетика: фенотип
- 59. - гомологичная рекомбинация/замещение гена -замещение гена: альтернативные
- 60. TALE: TAL (transcription activator-like) effectors Эффекторы TALE
- 61. Основные типы замолкания гена (сайленсинга) пост-транскрипционный сайленсинг
- 62. Белки, участвующие в метаболизме «малых» РНК у
- 63. Пример: выключение (сайленсинг) гена «икс» конститутивный (35S)
- 64. Регулируемое замолкание гена PDS (PHYTOENE
- 65. Регулируемое замолкание (индуцибельный сайленсинг) гена PDS (PHYTOENE DEASATURASE) No Dex Dex
- 66. Enhancer TATA En
- 67. Инсерционный мутагенез с использованием «ловушек»
- 68. Enhancer TATA
- 69. Принцип работы «ловушки для энхансеров»
- 70. Методы редактирования генома растений
- 71. Модификации генома: разновидности Изменение случайного места в геноме Редактирование генома
- 72. Редактирование генома Инактивация конкретного локуса Замена одного
- 73. Программируемые нуклеазы ZFN (Zink-Finger Nucleases) TALENs (Transcription
- 74. Нуклеазы с цинковыми пальцами В качестве ДНК-связывающего домена используется универсальный домен типа «Цинковые пальцы»
- 76. Как программируют ZFN? Wolfe et al., 1999
- 77. TALEN (Transcription Activator-Like Effectors Nucleases)
- 78. TALEN Kim, Kim, 2014 33-35 аминокислотные
- 79. Применение TALEN Потеря функции гена LOX3 (кодирует
- 80. Система CRISPR/Cas9 (Clustered regularly Interspaced Palindromic Repeats- CRISPR associated protein)
- 81. Система CRISPR/Cas9 В 2005 обнаружили, что
- 82. Работа системы CRISPR-Cas в бактериях Horvath, Barrangou, 2010
- 83. Čermák et al., 2015
- 84. Защита от вирусов с помощью CRISPR-Cas9 Ji et al., 2015
- 85. Рецептор ауксина Auxin binding protein 1 (ABP1) Chen et al., 2001
- 86. Gao et al., 2015
- 87. Рецептор ауксина Auxin binding protein 1 (ABP1) Chen et al., 2001

Слайд 2Лекция 6: Трансгенные растения-4
Получение инсерционных мутантов растений на основе агробактериальной трансформации
Клонирование
Изучение с помощью трансгенных растений экспрессии генов и структуры растительных промоторов
Получение растений со сверхэкспрессией гена с целью изучения функции гена
Использование методов «генетической хирургии» для изучения взаимодействия клеток и тканей в процессах развития
Использование трансформации для изучения роли гормонов в развитиии растений.

Слайд 3Т-ДНК инсерционный мутагенез
интеграция Т-ДНК
изменение структуры одного из генов за счет встраивания
….отбор по измененному фенотипу

Слайд 4Схемы используемых плазмид для получения:
рецессивных мутаций
доминантных мутаций
X - усилители транскрипции из


Слайд 6Клонирование гена agamous с помощью Т-ДНК инсерционного мутагенеза
ЭМС-мутагенез
10 лепестков,
4 чашелистика
агробактериальная
трансформация
мутанта ag-2
трансформированное растение с цветками дикого типа

Слайд 7Ген, маркированный транспозоном (tagged gene), можно клонировать с помощью «вытягивания за
, можно клонировать с помощью «вытягивания за транспозон»")
Слайд 8Транспозонный мутагенез
интеграция
Возможность клонировать
ген, «вытянув» его за транспозон

Слайд 9При встраивании транспозона в кодирующую последовательность гена, наблюдается мутантный фенотип.
При

 мутагенеза")
 мутагенеза")
Слайд 12Получение мутации flo613 и клонирование гена FLORICAULA львиного зева (Coen et
Выращивание 26 000 растений львиного зева с активным мобильным элементом Tam3 (Transposone of Antirrhinum majus) при tº=15ºC, вызывающей наибольшую частоту транспозиций (растения М1).
Самоопыление растений М1 с целью выявления рецессивных мутаций, возникших в М1 в результате перемещения Tam3.
Выявление 15 разных гомеозисных мутаций среди 80 000 растений М2, в том числе и flo613.
Укоренение и вегетативное размножение веточек мутанта flo613 при tº=15ºC, для выявления реверсий. Генетический анализ.
Выращивание 26 000")
Слайд 13 Рестрикционный анализ ДНК из выделенных клонов, необходимый для выявления участков
Субклонирование растительной ДНК с целью дальнейшего использования в качестве пробы
Создание геномной библиотеки из растений дикого типа
Скрининг библиотеки с меченой пробой растительной ДНК, фланкирующей транспозон, для выявления клона, содержащего ген FLO
Доказательство того, что выявленные клоны содержат ген FLO

Слайд 14Создание геномной библиотеки из мутанта flo 613, выращиваемого при tº=25ºC (Тam3
")
Слайд 15Скрининг библиотеки с меченой пробой из Tam3 для выявления клонов, содержащих
Получение отпечатков клонов библиотеки на нитроцеллюлозных фильтрах
Гибридизация фильтров с меченой пробой (чаще – кДНК меченая Р32)
Авторадиография

Слайд 16 Д.т. flo 613
9.0
5.5
1.5
Сравнение рестрикционных карт клонов,
Блот-гибридизация по Саузерну (гибридизация ДНК мутанта и дикого типа с пробой растительной ДНК фланкирующей транспозон)

Слайд 17Ген FLORICAULA – уникальный ген, присутствующий в геноме покрытосеменных в 1
Наиболее активная экспрессия гена FLORICAULA наблюдается в закладыва-ющихся примордиях цветка.
Клонирование гена открывает возможность для изучения роли кодируемого им белка и мРНК во взаимодействии клеток

Слайд 18 Изучение механизмов регуляции экспрессии генов растений с использованием
Возможности метода:
Выявление факторов (внешних и внутренних), определяющих особенности экспрессии исследуемого гена
Выявление цис-регуляторных элементов гена, отвечающих на действие внешних и внутренних факторов
 генов")

Слайд 20Green fluorescent protein (GFP)
Luciferase (LUC)
ß-glucuronidase (GUS)
Promoter-reporter fusion:
Использование гибридных конструкций, содержащих
репортерный
- флуоресцентный белок - стабильный,
- позволяет определить локализацию продукта на клеточном, тканевом уровне
- не дает точной количественной оценки экспрессии (полуколичественный метод)
широко используется для изучения активности генов, т.к. позволяет выявить слабый уровень экспрессии
позволяет определить локализацию продукта на клеточном, тканевом уровне,
белок стабильный, накапливается в тканях
- не дает точной количественной оценки экспрессии (полуколичественный метод)
позволяет определить локализацию продукта на тканевом уровне,
дает наиболее точную количественную оценку
может использоваться для изучения динамических процессов (белок короткоживущий)
pCO::GUS
pSCR::GFP
Luciferase (LUC)ß-glucuronidase (GUS)Promoter-reporter fusion:Использование гибридных конструкций, содержащих репортерный ген, «слитый» с промотором")
Слайд 21В трансформированных растениях проводят анализ экспрессии репротерного гена, отбирают трансформантов с

Слайд 22Анализ распределение ауксина в растениях с помощью генетической конструкции DR5:GUS
DR5 –



Слайд 25Фенотип трансгенных растений 35S::LFY
У растений 35S::LFY ген LFY экспрессируется и в

Слайд 26Характер экспрессии генов-ортологов LFY/FLO может определять тип соцветия (открытое или закрытое)
")
Слайд 27Метод позволяет изучить регуляторную функцию любого участка гена
Направленный мутагенез промоторной области


Слайд 29Основные семейства транскрипционных факторов растений
Консервативные для эукариот:
Уникальные для растений:
ТФ с MADS-доменом
ТФ
ТФ MYB
ТФ bZIP
etc.
ТФ GRAS
ТФ AP2-like
ТФ с В3-доменом
ТФ с GARP/ARRM-доменом
etc.
MYB

Слайд 30
Гомеодомен-ДНК - комплекс
вариабельный уч.
гомеодомен
N
С
Структура транскрипционного фактора с ДНК-связывающим гомеодоменом

Слайд 31
Гены семейств KNOX (STM) и WOX
Гены семейства KNOX арабидопсиса
Гены WOX арабидопсиса и других растений
 и WOX (WUS) кодируют гомеодомен-содержащие транскрипционные факторыГены семейства")
Слайд 32ТФ с гомеодоменами
(семейство TALE)
KNOX
мишени: гены биосинтеза ЦК и ГК
программы развития:
1).
2). Развитие сложного
листа
stm
35S::
KNAT1
HD-ZIPIII
WOX
мишени: 1). гены ТФ KANADY (антагонисты HD-ZIPIII) 2). гены PIN
(регулируют транспорт ауксинов)
программы развития:
Адаксиально-абаксиальная симметрия листа
phb
WT
WT
вышележащие регуляторы: система CLAVATA
мишени: гены ARR-A -репрессоры ответа на ЦК (для WUS)
программы развития:
1). Идентичность зародыша и суспензора (WOX2 и WOX8)
2). Идентичность ПАМ и КАМ
(WUS и WOX5)
WUS
WOX5
WOX8
WOX2
KNOXмишени: гены биосинтеза ЦК и ГК программы развития:1). Развитие ПАМ2). Развитие сложного")
Слайд 33Ген KNOTTED1 был идентифи-цирован на основе доминантных мутаций kn1, вызывающих отсутствие

Слайд 34Зная о важной роли гомеобоксных генов в развитии животных, приступили к
По гомологии с KN1 клонированы другие гомеобоксные гены кукурузы (более 13), A.thaliana (7), табака, томатов, сои, риса, ячменя и др.

Слайд 35Сверхэкспрессия гена KNAT1 у арабидопсиса
На листьях возникали дополнительные меристематические очаги, из
Листья A. thaliana 35S::KNAT1 превращаются из простых в пальчаторассеченные, хотя фенотип сильно варьировал (Chuck, Linkoln, Hake, 1996)



Слайд 38Обнаружение связи между уровнем экспрессии KNOX-генов и усложнением структуры листа позволило
у видов с простым листом (арабидопсис, табак, кукуруза, рис) KNOX-гены экспрессируются только в АМ, но не в примордиях листьев;
у томата, имеющего сложный лист, эти гены экспрессируются и в примордиях листьев

Слайд 39MCM1 – регулятор транскрипции дрожжей
AG – гомеозисный ген арабидопсиса (класс С)
DEF
SRF – регулятор транскрипции человека
Структура белков, содержащих MADS-домены
DEF –гомеозисный ген львиного зева")
Слайд 40ТФ с MADS доменом
вышележащие регуляторы: ТФ LFY
мишени: ???
программы развития: развитие
AP1, AP3, PI, AG, SEP1,2,3
SOC1, FLC , CAL
вышележащие регуляторы:
1). ТФ FT 2). Белки FCA и FY
2). Polycomb-комплекс VRN
мишени: ген ТФ LFY
программы развития: развитие флоральной меристемы
PHERES
вышележащие регуляторы:
Polycomb-комплекс FIS
мишени: ???
программы развития: развитие зародыша и эндосперма

Слайд 41Исследования трансгенных растений с конститутивной экспрессией гомеозисных генов также доказывают верность
35S::AP3
35S::PI
35S::AP3
35S::PI
35S::AG

Слайд 42Цветки с двойным венчиком (лилейные)
Цветки с двойной чашечкой
Можно предполагать, что именно изменения характера экспрессии консервативных генов ортологов АВС лежат в основе всего разнообразия форм цветка покрытосеменных
Цветки с двойной чашечкой Мутации в АВС- генах приводят")
Слайд 43Использование методов «генетической хирургии» для изучения взаимодействия клеток и тканей в

Слайд 44Day, Galgoci, Irish, 1995. Генетическое удаление лепестков и тычинок для выяснения
Модели, описывающие процесс возникновения органов цветка:
«Последовательная» модель
Wardlow, 1957.
Порядок возникновения органов цветка отражает последовательность индуктивных сигналов (развитие органов каждого последующего круга зависит от сигналов, поступающих из предыдущих кругов

Слайд 45«Пространственная» модель Holder, 1979. Под влиянием позиционных сигналов происходит разметка ФМ,

Слайд 46В основе «генетической хирургии» лежит метод трансформации растений химерными генами следующей
Кодирующая часть гена – последовательность, кодирующая продукт, вызывающий гибель растительных клеток и не способный перемещаться в другие клетки:
а) ген Corynebacterium diphtheriae, кодирующий дифтерийный токсин DT-A-белок, который катализирует АДФ-рибозилирование фактора элонгации. Это ведет к ингибированию белкового синтеза и гибели клеток. В состав DT-A-белка не входит сигнальный пептид, белок не является секреторным, поэтому вызывает гибель только тех клеток, где ген экспрессируется;

Слайд 47гены, кодирующие рибонуклеазы: Т1 рибонуклеазу Aspergillus oryzae и барназу Bacillus amyloliquefaciens,
ген pehA из Burkholderia caryophilla PG2982 – бактерии, метаболизирующей глицерол-глифосат до глифосата
Глифосат – гербицид, ингибирующий синтез ароматических аминокислот путем связывания с EPSP. Токсичен для растений, E. сoli и др.

Слайд 48Целью работы Day, Galgoci, Irish (1995) была проверка существующих моделей с
 была проверка существующих моделей с использованием метода генетической хирургии")
Слайд 49Этапы работы
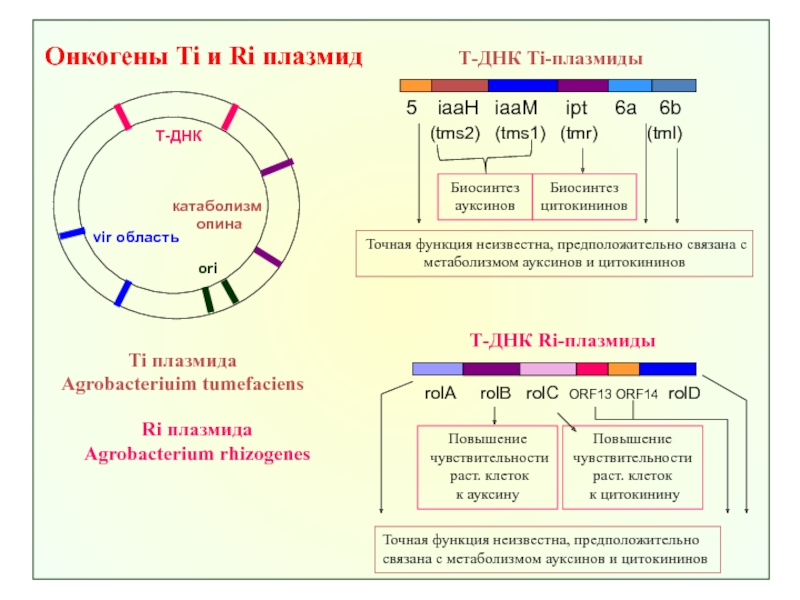
Слитые гены включали в Т-ДНК область (фрагмент Ti-плазмиды,который передается растению)
 бинарного вектора pBIADN и")
Слайд 50У резушки получено 5 трансгенных растений, в геноме которых присутствовали от
Все трансгенные растения имели одинаковый фенотип:органы 2 и 3 кругов (лепестки и тычинки) отсутствовали, а чашелистики и пестики имели нормальную морфологию. Следовательно, разметка кругов начинается на ранних стадиях до экспрессии трансгена во ФМ, т.е. верна пространственная модель
РЕЗУЛЬТАТЫ:


Слайд 53Корончатый галл на растении табака как результат трансформации агробактерией (штамм дикого
")

")


Слайд 58Прямая генетика: фенотип
Прямая и обратная генетика в изучении функции гена

Слайд 59- гомологичная рекомбинация/замещение гена -замещение гена: альтернативные подходы (Zn-fingers nucleases, TALEN) - РНК-интерференция/
Подходы, используемые обратной генетикой:
 - РНК-интерференция/ генный сайленсинг")
Слайд 60TALE: TAL (transcription activator-like) effectors
Эффекторы TALE – регуляторные белки, секретируются бактериями
TALE – оказывают влияние на экспрессию генов растения-хозяина, тем самым обуславливая восприимчивость или устойчивость к заболеванию
Kay et al. 2009
AvrBs3 – связывается с UPA-box, активируют гены-мишени – в т.ч. Bs3, запускающий реакцию СВЧ
 effectorsЭффекторы TALE – регуляторные белки, секретируются бактериями Xanthomonas (патогены растений) с")
Слайд 61Основные типы замолкания гена (сайленсинга)
пост-транскрипционный сайленсинг
(регуляция экспресии гена на уровне трансляции)
PTGS
транскрипционный сайленсинг (регуляция экспрессии гена на уровне транскрипции)
TGS - Transcriptional gene silencing
Специфическое
разрезание мРНК (А)
Репрессия трансляции (В)
Метилирование гистонов, гетерохроматинизация (С)
пост-транскрипционный сайленсинг(регуляция экспресии гена на уровне трансляции)PTGS - Post-transcriptional gene silencingтранскрипционный")
Слайд 62Белки, участвующие в метаболизме «малых» РНК у растений
DCL1 (DICER-like) –
HYL1 (HYPONASTIC LEAVES) ядерный dsRNA-связывающий белок
HEN1 (HUA ENHANCER 1)
HEN1 и HYL1 участвуют в ядерном процессинге «малых» РНК
HST (HASTY) – гомолог экспортина, участвует в транспорте miRNA из ядра в цитоплазму
AGO (ARGONAUTE)- рибонуклеаза, компонент RISC (RNA-induced silencing complex)
 – рибонулеказа, участвующая в процессинге")
Слайд 63Пример: выключение (сайленсинг) гена «икс»
конститутивный
(35S)
регулируемый
(индуцибельный)
Интроны
промотор
икс
ски
dsRNA
RISC –
RNA-induced
silencing
DICER-подобная рибонуклеаза
mRNA
деградация транскрипта, репрессия трансляции
Конструкция, используемая для трансформации
 гена «икс»конститутивный(35S)регулируемый (индуцибельный)Интроныпромоторикс скиdsRNA RISC – RNA-induced silencing complexDICER-подобная рибонуклеазаmRNAдеградация транскрипта, репрессия")
Слайд 64
Регулируемое замолкание гена PDS (PHYTOENE DEASATURASE)
LB
RB
RB
LB
35S
LhGR
GUS
op
PDS
PDS
intron
pHELLSTOP
PDS-
PHYTOENE
DEASATURASE
оператор
b-глюкоронидаза
Нет ИНДУКТОРА
Добавление ИНДУКТОРА: + DEX дексаметазон
LhGR – транскрипционный фактор (ТФ)
Неактивный ТФ
HSP90
LB
RB
RB
LB
35S
LhGR
GUS
op
PDS
PDS
intron
Экспрессия
b-глюкоронидазы (репортерного гена)
LhGR
Активный ТФ
Экспрессия
PDS- ингибирующей конструкции
DEX
Сайленсинг гена PDS
LBRBRBLB35SLhGRGUSopPDSPDSintronpHELLSTOPPDS-PHYTOENE DEASATURASEоператорb-глюкоронидазаНет ИНДУКТОРАДобавление ИНДУКТОРА: + DEX дексаметазон (стероидный лиганд)LhGR –")
Слайд 65Регулируемое замолкание (индуцибельный сайленсинг)
гена PDS (PHYTOENE DEASATURASE)
No Dex
Dex
 гена PDS (PHYTOENE DEASATURASE) No Dex Dex")
Слайд 66 Enhancer TATA
En En En En
Enhancer
Enhancer TATA
3. Enhancer trap (ловушка для энхансера)
2. Activation tag (активирующая последовательность)
1. Геномная ДНК дикого типа
Примеры использования Т-ДНК в качестве «зондов» для выявления кодирующих и регуляторных областей (Т-DNA tagging)
TATA reporter gene
Scholte M. T-DNA tagging in Medicago truncatula. 2002

Слайд 67 Инсерционный мутагенез с использованием «ловушек» промоторов
Репортерный ген без промотора

Слайд 68 Enhancer TATA
4. Ловушка для промотора (promoter trap) -1
reporter
гибридный белок
Инсерция в экзон
Enhancer TATA
5. Ловушка для промотора (promoter trap) -2
reporter gene
гибридный белок
Инсерция в интрон
SD SA
Splice aceptor sequence
сплайсинг
Scholte M. T-DNA tagging in Medicago truncatula. 2002
Примеры использования Т-ДНК в качестве «зондов» для выявления кодирующих и регуляторных областей (Т-DNA tagging)
 -1reporter geneгибридный белок Инсерция в экзон")
Слайд 69Принцип работы
«ловушки для энхансеров»
GAL4:GFP
Примеры использования Т-ДНК в качестве «зондов»



Слайд 72Редактирование генома
Инактивация конкретного локуса
Замена одного локуса на другой с помощью гомологичной
Этот путь затруднён у растений.
Введение DSB также существенно увеличивает вероятность гомологичной рекомбинации
Чтобы инактивировать конкретный локус, надо осуществить в нём разрыв двунитевой ДНК (DSB)
Этот путь затруднён")
Слайд 73Программируемые нуклеазы
ZFN (Zink-Finger Nucleases)
TALENs (Transcription Activator-Like Effector Nucleases)
CRISPR-Cas система (Clustered regularly
TALENs (Transcription Activator-Like Effector Nucleases)CRISPR-Cas система (Clustered regularly Interspaced Palindromic Repeats- CRISPR")
Слайд 74Нуклеазы с цинковыми пальцами
В качестве ДНК-связывающего домена используется универсальный домен типа

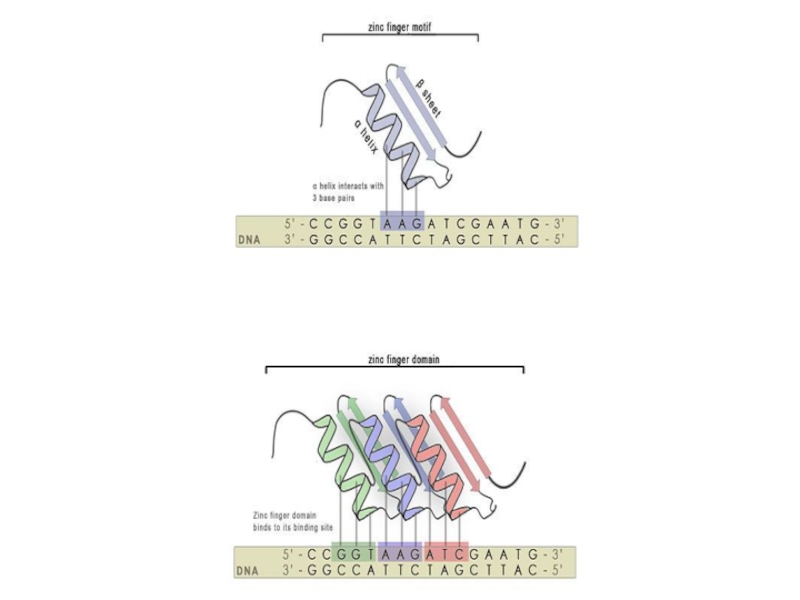
Слайд 76Как программируют ZFN?
Wolfe et al., 1999
Каждый цинковый палец распознаёт 3 (4)
Можно подбирать цинковые пальцы под последовательности
Обычно хорошо режутся не все последовательности, а только GNN. (т.е., например, 9-нуклеотидная последовательность должна быть GNNGNNGNN)
В среднем хорошая специфичная последовательность для разрезания попадается один раз на 100 нп.
 нуклеотида. Обычно используют 3-6")
")
Слайд 78TALEN
Kim, Kim, 2014
33-35 аминокислотные повторы
12ый -13ый аминокислотные остатки (repeat variable
 связываются с определённым нуклеотидом")
Слайд 79Применение TALEN
Потеря функции гена LOX3 (кодирует липоксигеназу, создающую перекись водорода) увеличивает
Этот ген инактивировали с помощью TALEN
Ma et al., 2015
Рис дикого типа
Рис lox3
После обработки для ускоренного старения
 увеличивает сроки хранения семянЭтот ген")
Слайд 80Система CRISPR/Cas9 (Clustered regularly Interspaced Palindromic Repeats- CRISPR associated protein)
")
Слайд 81Система CRISPR/Cas9
В 2005 обнаружили, что спейсеры гомологичны последовательностям фагов и конъюгативных
Streptococcus thermophilus: Устойчивость к фагам коррелировала с составом спейсеров (Barrangou et al., 2007)
Гены cas тоже необходимы для устойчивости (Barrangou et al., 2007)




Chen et al., 2001")

Chen et al., 2001")