- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Ретротранспозоны и ДНК транспозоны презентация
Содержание
- 1. Ретротранспозоны и ДНК транспозоны
- 5. Мобильные генетические элементы В 40-х годах
- 6. Первое описание элементов геномной ДНК, характеризующихся наследуемой
- 7. Все МГЭ по способу перемещения могут быть
- 8. Оба класса делятся последовательно на подклассы, суперсемейства
- 9. Мутации, создаваемые инсерцией ретротранспозонов, более стабильны, чем
- 10. Вырезание и встраивание МГЭ индуцирует мутагенез,
- 11. МГЭ и трансгенные технологии Накопление знаний о
- 12. Способность свободноживущих плоских червей к регенерации значительных
- 13. Помимо фундаментального значения, практический интерес может иметь
- 14. Как членов фракции повторов, МГЭ вначале считали
- 15. Геномная доместикация может приводить к существенной функциональной
- 16. МГЭ и горизонтальный перенос генов Мобильные элементы
- 17. В геноме F. hepatica среди ретротранспозонов обнаружен
- 18. Имеющийся объем знаний по изучению HGT позволяет



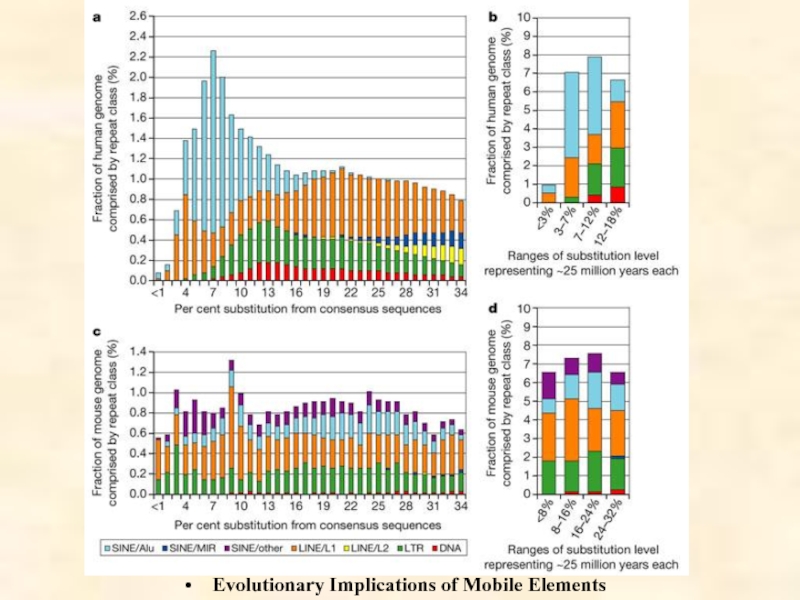
Слайд 5Мобильные генетические элементы
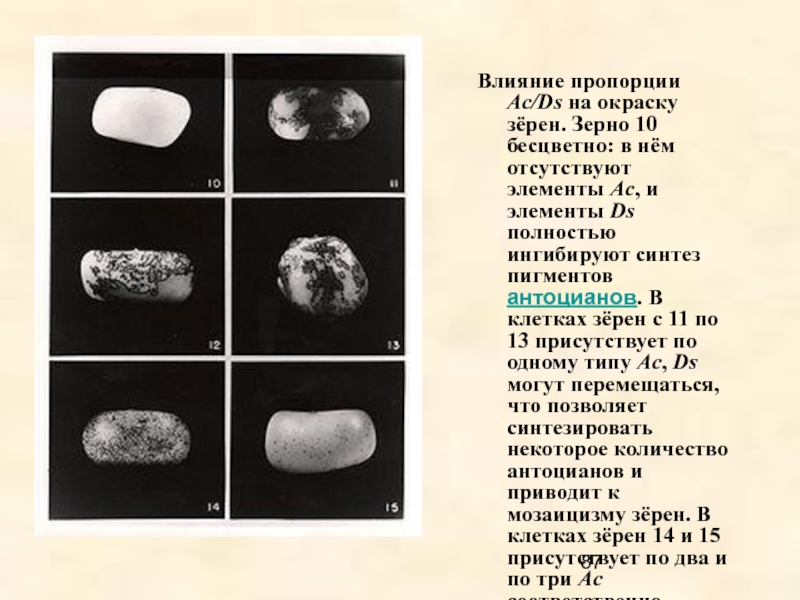
В 40-х годах Барбара Мак-Клинток, американский генетик, обнаружила
мозаичность окраски зерен у кукурузы, небъяснимую законами Менделя и мутационной теорией.
Она предположила, что некоторые гены могут менять свое место в геноме. В 70-х годах это предположение получило экспериментальное подтверждение.
Она предположила, что некоторые гены могут менять свое место в геноме. В 70-х годах это предположение получило экспериментальное подтверждение.

Слайд 6Первое описание элементов геномной ДНК, характеризующихся наследуемой способностью передвигаться внутри и
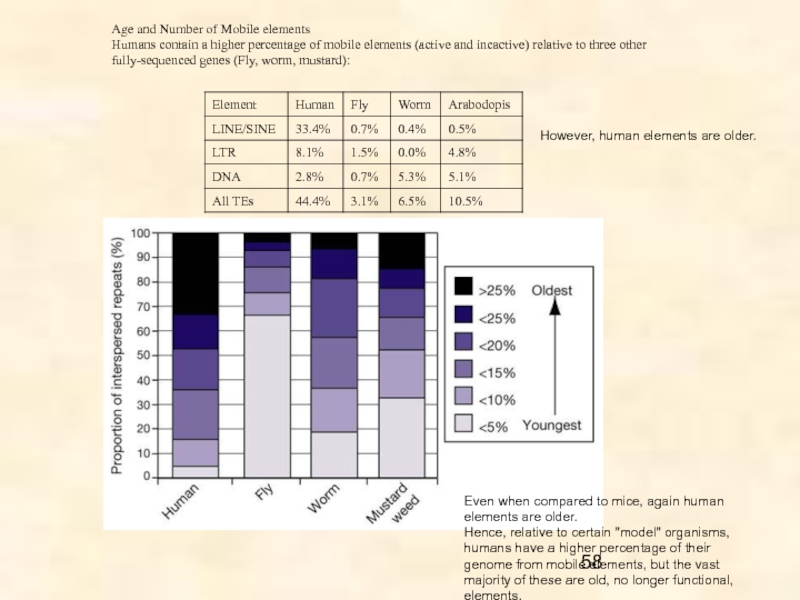
между геномами, было сделано в 1951 г. прошлого века Барбарой Мак-Клинток для кукурузы. Благодаря развитию технологий геномного секвенирования, к настоящему времени эти мобильные генетические элементы (МГЭ) обнаружены в геномах самых разных видов, часто получая названия, отражающие их способность к передвижению (hobo – бродяга, gypsy – цыган, flea – блоха и т.д.). Содержание МГЭ варьирует и может достигать 85% (Zea mays) генома, причем максимальное разнообразие МГЭ демонстрируют беспозвоночные. В малых геномах число копий МГЭ на семейство низкое, но в больших геномах оно значительно варьирует.

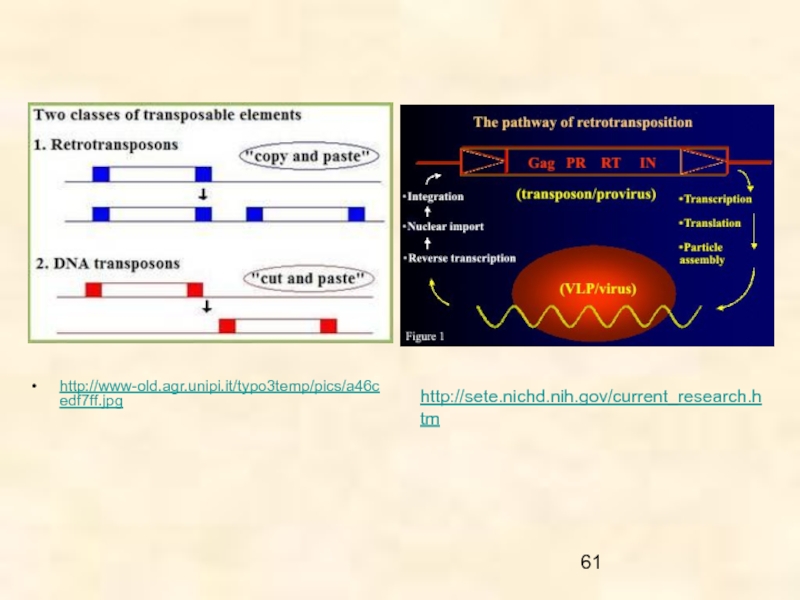
Слайд 7Все МГЭ по способу перемещения могут быть сгруппированы в два основных
класса:
1 - ретротранспозоны, перемещающиеся с помощью РНК-посредника («copy and paste»), и
2 - ДНК транспозоны (перемещается непосредственно ДНК).
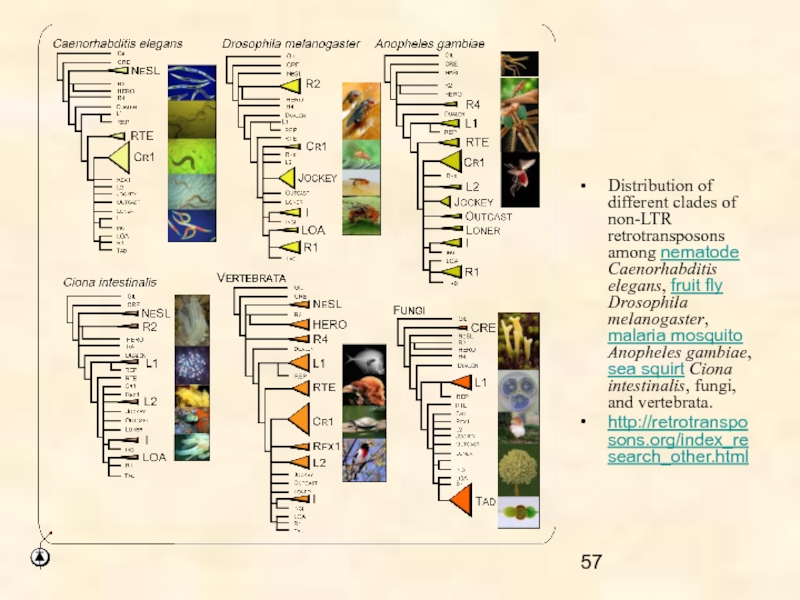
Ретротранспозоны обнаружены у всех эукариот, от грибов до млекопитающих и, по-видимому, эти МГЭ связаны с половым размножением.
ДНК транспозоны – самая древняя группа МГЭ, распространенная среди про- и эукариот.
Интересно, что МГЭ первого класса были обнаружены как у плоских, так и круглых червей, а МГЭ второго класса оказались типичными для паразитических нематод, но не для трематод и ленточных червей. К настоящему времени только у гельминтов млекопитающих описано более 30 видов МГЭ.
1 - ретротранспозоны, перемещающиеся с помощью РНК-посредника («copy and paste»), и
2 - ДНК транспозоны (перемещается непосредственно ДНК).
Ретротранспозоны обнаружены у всех эукариот, от грибов до млекопитающих и, по-видимому, эти МГЭ связаны с половым размножением.
ДНК транспозоны – самая древняя группа МГЭ, распространенная среди про- и эукариот.
Интересно, что МГЭ первого класса были обнаружены как у плоских, так и круглых червей, а МГЭ второго класса оказались типичными для паразитических нематод, но не для трематод и ленточных червей. К настоящему времени только у гельминтов млекопитающих описано более 30 видов МГЭ.

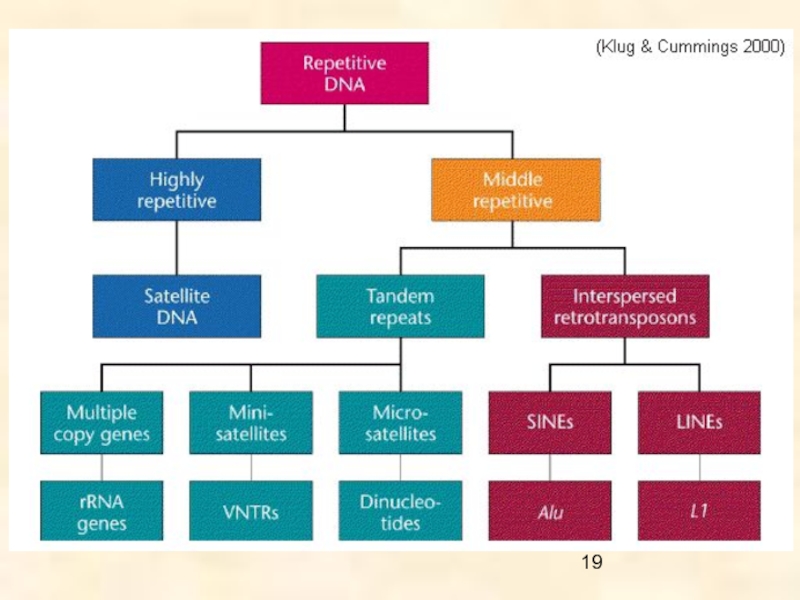
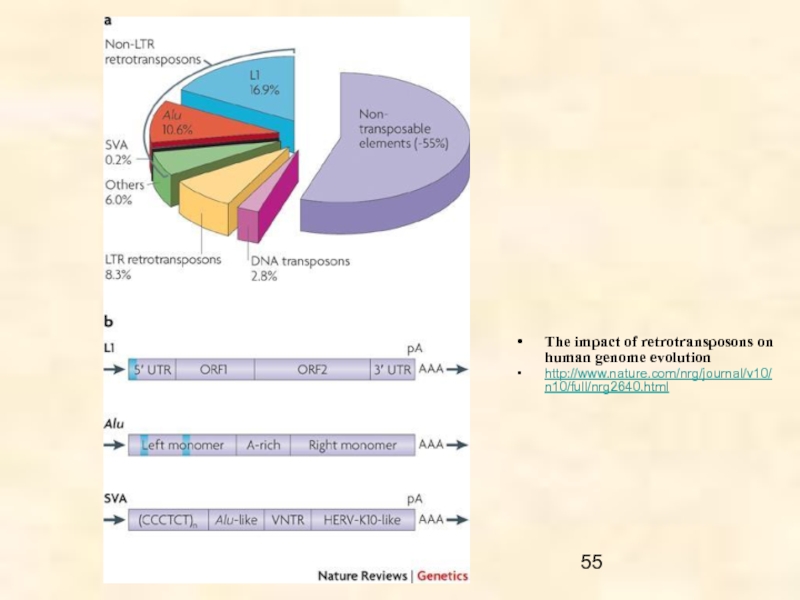
Слайд 8Оба класса делятся последовательно на подклассы, суперсемейства и семейства, согласно механизму
их интеграции.
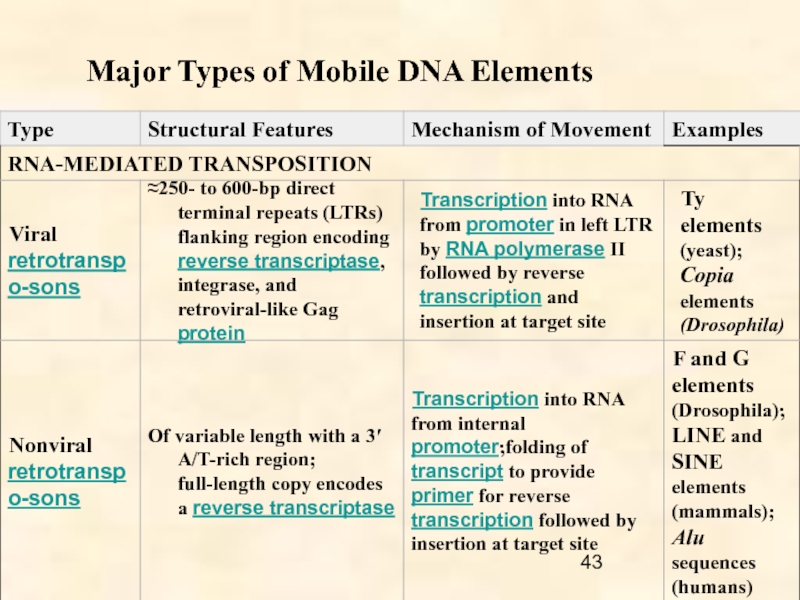
Ретротранспозоны обнаружены у всех эукариот от грибов до млекопитающих; они включают
- LTR-ретротранспозоны,
- non-LTR ретротранспозоны и
- тирозин-рекомбинантные ретротранспозоны.
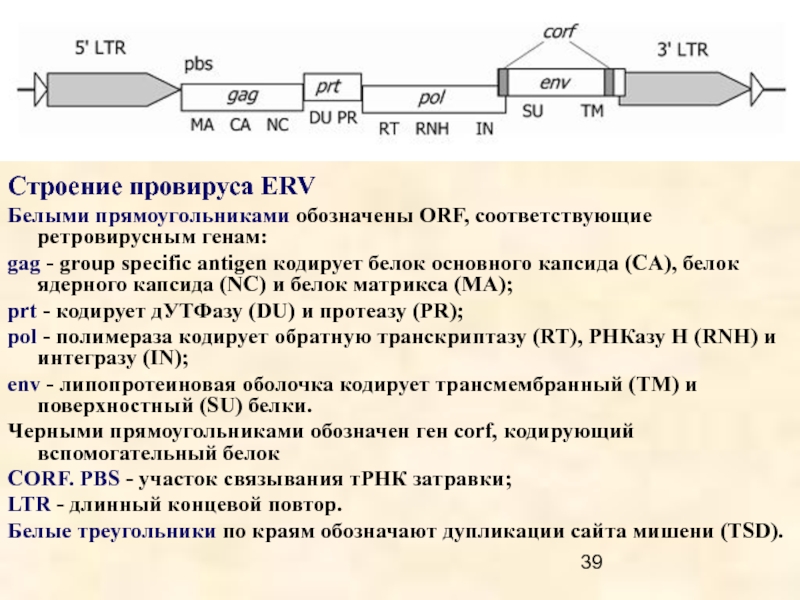
LTR ретротранспозоны объединяют пять групп: Copia, BEL, Gypsy, эндогенные ретровирусы (ERV) и DIRS.
non-LTR ретротранспозоны включают автономные LINES (перемещающиеся самостоятельно) и неавтономные SINES (требующие для своего перемещения наличия автономных МГЭ) элементы.
Ретротранспозоны, по-видимому, связаны с половым размножением, поскольку среди представителей 24 типов животных они не были обнаружены только у размножающихся бесполым путем коловраток.
Для своей экспансии ретротранспозоны вначале транскрибируются РНК полимеразой хозяина, а затем копируются собственной обратной транскриптазой. Поскольку оба фермента не обладают редактирующей способностью, эти копии имеют тенденцию приобретения случайных замен оснований. Такие замены часто инактивируют последовательности-потомки и приводят к утрате их мобильности (“dead-on-arrival”).
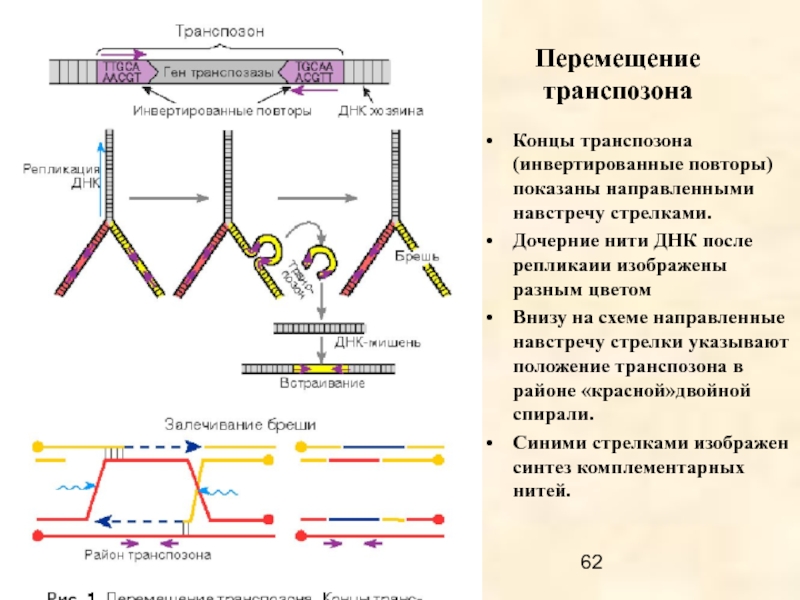
ДНК транспозоны – самая древняя группа МГЭ, распространенная среди про- и эукариот; по механизмам транспозиции подразделяется на «cut-and-paste» транспозоны, самореплицирующиеся транспозоны и транспозоны «катящегося кольца» (полурепликативная транспозиция).
Ретротранспозоны обнаружены у всех эукариот от грибов до млекопитающих; они включают
- LTR-ретротранспозоны,
- non-LTR ретротранспозоны и
- тирозин-рекомбинантные ретротранспозоны.
LTR ретротранспозоны объединяют пять групп: Copia, BEL, Gypsy, эндогенные ретровирусы (ERV) и DIRS.
non-LTR ретротранспозоны включают автономные LINES (перемещающиеся самостоятельно) и неавтономные SINES (требующие для своего перемещения наличия автономных МГЭ) элементы.
Ретротранспозоны, по-видимому, связаны с половым размножением, поскольку среди представителей 24 типов животных они не были обнаружены только у размножающихся бесполым путем коловраток.
Для своей экспансии ретротранспозоны вначале транскрибируются РНК полимеразой хозяина, а затем копируются собственной обратной транскриптазой. Поскольку оба фермента не обладают редактирующей способностью, эти копии имеют тенденцию приобретения случайных замен оснований. Такие замены часто инактивируют последовательности-потомки и приводят к утрате их мобильности (“dead-on-arrival”).
ДНК транспозоны – самая древняя группа МГЭ, распространенная среди про- и эукариот; по механизмам транспозиции подразделяется на «cut-and-paste» транспозоны, самореплицирующиеся транспозоны и транспозоны «катящегося кольца» (полурепликативная транспозиция).

Слайд 9Мутации, создаваемые инсерцией ретротранспозонов, более стабильны, чем мутации, вызванные ДНК транспозонами,
поскольку в отличие от последних, ретротранспозоны, встроившись в геном уже никогда не исчезают.
Передвигаясь по геному и интегрируя в новые сайты, МГЭ могут обеспечить потенциально вредную генетическую изменчивость, от полиморфизма простой последовательности, до кардинальных изменений в структуре хромосом, их целостности и инактивации соседних генов. Показано, например, что все non-LTR ретротранспозоны или ретротранспозон-подобные последовательности трипаносом T. brucei и T. cruzi, а также лейшмании Leishmania major имеют тенденцию локализации в местах хромосомных инверсий, «strand-switch» регионов и хромосомных концов.
Передвигаясь по геному и интегрируя в новые сайты, МГЭ могут обеспечить потенциально вредную генетическую изменчивость, от полиморфизма простой последовательности, до кардинальных изменений в структуре хромосом, их целостности и инактивации соседних генов. Показано, например, что все non-LTR ретротранспозоны или ретротранспозон-подобные последовательности трипаносом T. brucei и T. cruzi, а также лейшмании Leishmania major имеют тенденцию локализации в местах хромосомных инверсий, «strand-switch» регионов и хромосомных концов.

Слайд 10
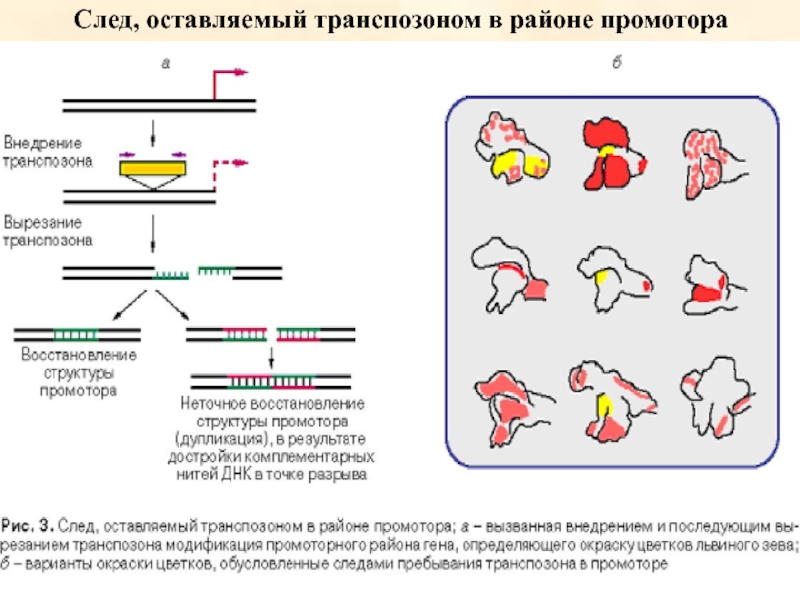
Вырезание и встраивание МГЭ индуцирует мутагенез, уровень которого может в тысячу
раз превышать скорость спонтанных мутаций. Поэтому для подавления неконтролируемой пролиферации МГЭ сформировались специальные защитные механизмы против их транспозиции, включающие эпигенетические процессы, такие как система транскрипционного и посттранскрипционного сайленсинга генов. Однако различные виды физиологического стресса, например, температурные изменения или колонизация новых мест обитания хозяевами, могут активировать МГЭ и привести к их пролиферации, разрушив эпигенетический сайленсинг.
С другой стороны, есть данные, предполагающие коэволюцию МГЭ и геномов хозяев, положительное влияние и важную роль этих элементов в формировании и поддержании хромосом хозяина, модификации генной экспрессии и участии в разделении генома на хромосомные домены с эпигенетическими метками.
С другой стороны, есть данные, предполагающие коэволюцию МГЭ и геномов хозяев, положительное влияние и важную роль этих элементов в формировании и поддержании хромосом хозяина, модификации генной экспрессии и участии в разделении генома на хромосомные домены с эпигенетическими метками.

Слайд 11МГЭ и трансгенные технологии
Накопление знаний о структуре и активности МГЭ может
пролить свет на эволюционную историю и филогенетические отношения самих элементов и генома хозяина (в частности, дать информацию об эволюции генов в сайтах интеграции МГЭ), а также будет способствовать развитию трансгенных технологий, полезных для изучения структуры, функции и контроля генов паразитических видов.
Виды Platyhelminthes рассматриваются в настоящее время как объекты трансфекции для изучения тканевой регенерации, дизайна новых и более эффективных антигельминтных препаратов и исследования молекулярных связей между паразитом и хозяином. Недавно, при изучении регенеративных процессов и МГЭ у саламандры Ambyostoma mexicanum и голотурии Holothuria glaberrima, была высказана гипотеза, что транскрипционная активность ретротранспозонов может специфически контролироваться хозяином и играть определенную роль в регенерации органов животных.
Виды Platyhelminthes рассматриваются в настоящее время как объекты трансфекции для изучения тканевой регенерации, дизайна новых и более эффективных антигельминтных препаратов и исследования молекулярных связей между паразитом и хозяином. Недавно, при изучении регенеративных процессов и МГЭ у саламандры Ambyostoma mexicanum и голотурии Holothuria glaberrima, была высказана гипотеза, что транскрипционная активность ретротранспозонов может специфически контролироваться хозяином и играть определенную роль в регенерации органов животных.

Слайд 12Способность свободноживущих плоских червей к регенерации значительных участков тела (подразумевающая присутствие
зародышевых клеток) привлекает внимание исследователей на протяжении нескольких последних десятилетий. Понимание основ регенирации у животных, в том числе плоских червей, может внести важный вклад в регенеративную медицину человека. Планарии, свободноживущие филогенетические родственники трематод, способны регенирировать целых червей из небольшого фрагмента их тела. Клетки, ответственные за эти регенеративные свойства, были названы «необластами».
Однако для изучения данного феномена необходимы новые более эффективные методы для интродукции чужеродных генов и векторы для стабильной трансфекции. Методом электропарации в геном необластов планарии Dugesia japonica были эффективно интродуцированы три транспозона: mariner, Herms и Piggybac, два из которых хорошо экспрессировались через восемь месяцев после трансфекции. Содержание паразитических видов червей в лабораторных условиях, а также генетические манипуляции с ними осуществлять сложнее. Тем не менее, уже известны успешные эксперименты по получению генетически модифицированных гельминтов - трематод, цестод и нематод. Практическое применение данных исследований лежит в сфере сайленсинга генов, вовлеченных в жизненно важные физиологические процессы, посредством РНК интерференции.
Однако для изучения данного феномена необходимы новые более эффективные методы для интродукции чужеродных генов и векторы для стабильной трансфекции. Методом электропарации в геном необластов планарии Dugesia japonica были эффективно интродуцированы три транспозона: mariner, Herms и Piggybac, два из которых хорошо экспрессировались через восемь месяцев после трансфекции. Содержание паразитических видов червей в лабораторных условиях, а также генетические манипуляции с ними осуществлять сложнее. Тем не менее, уже известны успешные эксперименты по получению генетически модифицированных гельминтов - трематод, цестод и нематод. Практическое применение данных исследований лежит в сфере сайленсинга генов, вовлеченных в жизненно важные физиологические процессы, посредством РНК интерференции.
 привлекает внимание")
Слайд 13Помимо фундаментального значения, практический интерес может иметь способность ретроэлементов выполнять важные
функции в эмбриогенезе, в частности, влиять на плюрипотентность клеток.
Ретротранспозоны часто видо- или родоспецифичны, что открывает перспективу разработки чувствительных и специфичных молекулярных диагностикумов, особенно необходимых для паразитических видов.
Кроме того, изучение МГЭ представляет определенный интерес для эпидемиологических и медицинских исследований, поскольку они могут генерировать специфические генетические паттерны, ассоциированные с клиническими проявлениями заболеваний.
Ретротранспозоны часто видо- или родоспецифичны, что открывает перспективу разработки чувствительных и специфичных молекулярных диагностикумов, особенно необходимых для паразитических видов.
Кроме того, изучение МГЭ представляет определенный интерес для эпидемиологических и медицинских исследований, поскольку они могут генерировать специфические генетические паттерны, ассоциированные с клиническими проявлениями заболеваний.

Слайд 14Как членов фракции повторов, МГЭ вначале считали «мусорной», «эгоистичной» ДНК, или
«геномными паразитами». Однако накопление весомых свидетельств о важной роли МГЭ в эволюции генов и геномов приводит к пониманию того обстоятельства, что помимо вредных эффектов эти геномные элементы вносят позитивный вклад в эволюцию генома их хозяев и поэтому могут быть переименованы в «потенциально полезные доместицированные элементы».

Слайд 15Геномная доместикация может приводить к существенной функциональной эволюции МГЭ.
- Яркий
пример – влияние этих элементов на эволюцию адаптивной иммунной системы челюстных позвоночных. Домен Rag1, участвующий в обеспечении образования огромного разнообразия антител, высоко гомологичен транспозазе ДНК транспозонов transib, которые были обнаружены в геномах беспозвоночных, но не идентифицированы у позвоночных.
- Предполагают, что ретротранспозон LITc генома трипаносомы Trypanosoma cruzi может участвовать в репарации ДНК, а
- ген SETMAR, участвующий в каскаде репараций двойных разрывов ДНК, произошел от транспозазы mariner.
- Есть основания полагать, что теломераза, по-видимому, произошла от обратной транскриптазы ретротранспозонов.
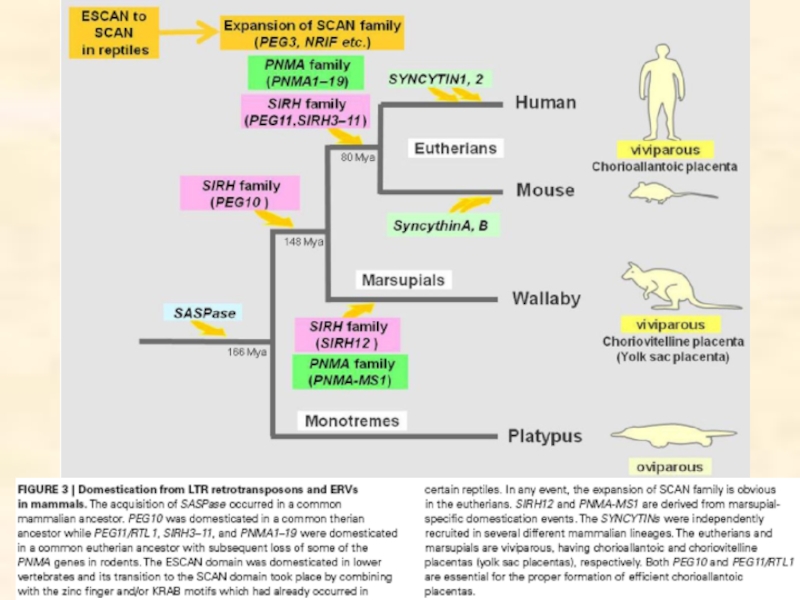
- Считается, что доместицированные гены ретроэлементов PNMA, SIRH и SYNCYTIN, необходимые для нормального развития плаценты, значительно повлияли на радиацию млекопитающих.
- Наконец, описано большое число генов, произошедших от транспозонов, которые приобрели функцию регуляторов транскрипции.
- Предполагают, что ретротранспозон LITc генома трипаносомы Trypanosoma cruzi может участвовать в репарации ДНК, а
- ген SETMAR, участвующий в каскаде репараций двойных разрывов ДНК, произошел от транспозазы mariner.
- Есть основания полагать, что теломераза, по-видимому, произошла от обратной транскриптазы ретротранспозонов.
- Считается, что доместицированные гены ретроэлементов PNMA, SIRH и SYNCYTIN, необходимые для нормального развития плаценты, значительно повлияли на радиацию млекопитающих.
- Наконец, описано большое число генов, произошедших от транспозонов, которые приобрели функцию регуляторов транскрипции.

Слайд 16МГЭ и горизонтальный перенос генов
Мобильные элементы генома являются основными кандидатами на
роль переносчиков в рамках гипотезы горизонтального переноса ДНК (Horizontal Gene Transfer, HGT) между репродуктивно изолированными видами. События HGT рассматриваются сейчас, как важная движущая сила эволюции эукариотического генома. Причем считается, что система паразит-хозяин является оптимальной моделью для изучения данного феномена. Приобретение МГЭ de novo считается доказанным, если гомология первичных последовательностей в геномах неродственных организмов составляет более 90%.

Слайд 17В геноме F. hepatica среди ретротранспозонов обнаружен повтор BovB (67 тыс.
полных или частичных копий). Ранее для него был описан горизонтальный перенос между змеями и жвачными, а наличие данного элемента у F. hepatica предполагает, что перенос BovB между различными таксонами позвоночных может усиливаться трематодной инфекцией.
В геноме анкилостомы Ancylostoma caninum содержится ДНК транспозон bandit. Его ближайшим филогенетическим родственником является ДНК транспозон человека Hsmar, что предполагает возможность горизонтального переноса между анкилостомами и их хозяевами-млекопитающими.
В геноме анкилостомы Ancylostoma caninum содержится ДНК транспозон bandit. Его ближайшим филогенетическим родственником является ДНК транспозон человека Hsmar, что предполагает возможность горизонтального переноса между анкилостомами и их хозяевами-млекопитающими.
.")
Слайд 18Имеющийся объем знаний по изучению HGT позволяет сделать ряд важных заключений:
(1) для HGT не требуется долговременной физической ассоциации между видами, но физический контакт может увеличить вероятность HGT;
(2) смена хозяев может затруднить идентификацию HGT, ассоциированную с паразитизмом;
(3) обнаружение МГЭ часто идентифицируется как HGT, однако следует проверять, нет ли первичного механизма, обусловившего его распространение;
(4) HGT часто подвергается сомнению из-за потенциально возможной контаминации, и эта проблема не должна оставаться без внимания.
Дальнейшие исследования HGT могут в значительной мере пополнить наши представления о геномной коэволюции и дать эволюционое понимание эпидемиологии паразитарных инфекций.
 для HGT не")