- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Мембранний транспорт.Іонні канали.Електрична активність клітин різного типу. презентация
Содержание
- 1. Мембранний транспорт.Іонні канали.Електрична активність клітин різного типу.
- 2. 1.Мембранний транспорт 1.1 Пасивний транспорт. Проста дифузія.
- 3. 1. Транспорт речовин через мембрани Біологічні
- 6. Мембранні транспортні процеси, що відбуваються
- 7. Активний транспорт поділяють
- 8. 1.1 Пасивний транспорт. Проста дифузія Дифузійні процеси
- 9. 1.2 Пасивний транспорт.Полегшена дифузія Полегшена дифузія, порівняно
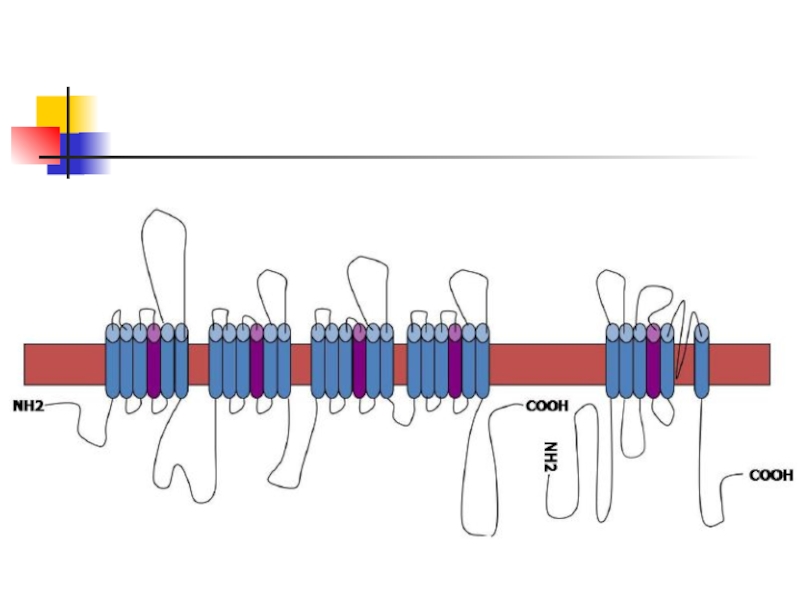
- 11. 2. Іонні канали За механізмом відкриття виділяють
- 12. 2.1 Ліганд-залежні іонні канали
- 13. Внутрішньоклітинними лігандами найчастіше є фосфатні залишки, що
- 14. 2.2 Потенціалзалежний Na+-канал.
- 15. Відома низка сполук, здатних специфічно блокувати Na+-канали.
- 16. Блокатори потенціалзалежних
- 17. Сакситоцин
- 18. Активатори
- 19. Батрахотоксин -
- 20. Токсини цигуатера продукуються
- 21. Продуцент бревітоксину – дінофлагелят
- 22. 2.3 Потенціал-залежні
- 24. Блокатори
- 25. Апамін (Apis mellifera)
- 26. Агітоксин-2 (отрута жовтого скорпіону Leiurus quinquestriatus herbraeus)
- 27. 2.4 Са2+ - канали За механізмом відкриття
- 28. Конотоксини як блокатори
- 29. κ- конотоксин інгібує калієві канали
- 30. 2.5 Хлорні канали Біологічні мембрани різних
- 31. Біоелектричні потенціали (біоструми) - електричні явища, що
- 32. Дякую за увагу!!!
- 33. (3.7) (3.8) k-коефіцієнт
- 34. Осмос – це дифузія молекул води через
- 35. Рівняння осмосу аналогічне рівнянню (3.4): (3.12)
- 37. 2.4 Дифузія іонів через мембрану На мембрані
- 40. АХ А А
- 42. Рис.3.6. Іонофори є двох видів: переносники і
- 43. 2.6 Активний транспорт. Тепер можна впевнено
- 44. Рис.3.7 К 3 +
- 45. L+
- 46. Згідно з однією з таких гіпотез Na+
- 47. Сучасні уявлення про механізм дії натрій-калієвої помпи

Слайд 21.Мембранний транспорт
1.1 Пасивний транспорт. Проста дифузія.
1.2 Пасивний транспорт.Полегшена дифузія.
2. Іонні канали
2.1
2.2 Потенціалзалежний Na+-канал.
2.3 Потенціал-залежні калієві канали.
2.4 Са2+ - канали.
2.5 Хлорні канали.
3. Електрична активність клітин різного типу

Слайд 31. Транспорт речовин через мембрани
Біологічні мембрани виконують багато різноманітних функцій.

Слайд 4
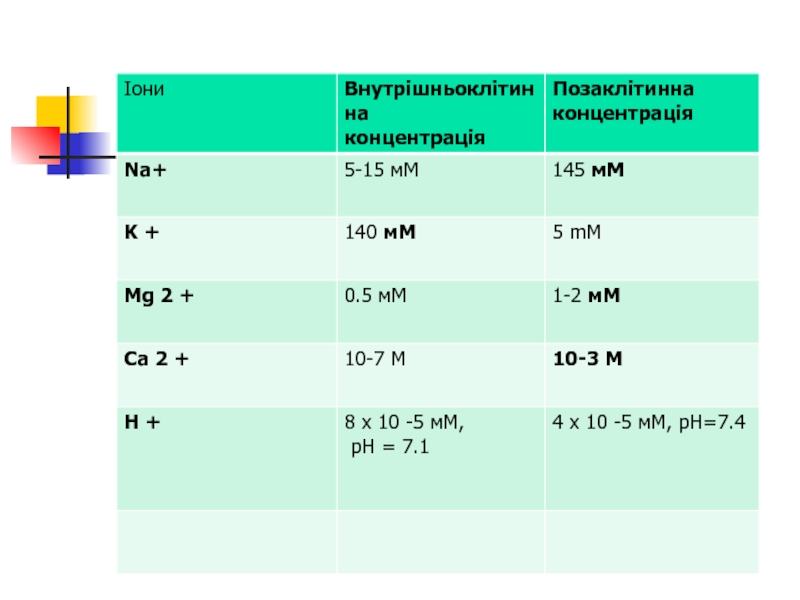
Плазматична мембрана, як і інші біологічні мембрани, є напівпроникною, тобто має неоднакову проникність для різних сполук. Гідрофобні сполуки, а також О2, СО2, N2 можуть вільно проходити крізь мембрану, у той час як заряджені іони взагалі не здатні її перетинати без використання спеціальних молекул-помічників. Малі і великі незаряджені полярні молекули (вода, сечовина, гліцерил та глюкоза і сахароза, відповідно), мають певні утруднення під час транспорту крізь мембрану, які більш виражені для великих молекул, і тому для більш ефективного переносу також потребують транспортних молекул.
Завдяки існуванню специфічних транспортних систем плазматична мембрана є основною структурою, що забезпечує створення і підтримку концентраційного (табл. 1.1) й електрохімічного градієнтів і залучає клітину до обміну речовин.

Слайд 6 Мембранні транспортні процеси, що відбуваються у клітині, можна віднести
Під час пасивного транспорту речовина переноситься за градієнтом концентрації без залучення енергії гідролізу АТФ. Пасивний транспорт, в свою чергу, може здійснюватися шляхом простої або полегшеної дифузії (в останньому випадку - за допомогою специфічного переносника, каналу чи іонофору).
Під час активного транспорту рух речовини здійснюється проти концентраційного градієнта за рахунок енергії, що звільнюється при гідролізі АТФ.

Слайд 7
Активний транспорт поділяють на первинно-активний (його опосередковують ферменти-АТФази, напр., Na+,

Слайд 81.1 Пасивний транспорт. Проста дифузія
Дифузійні процеси – це самовільне переміщення речовини

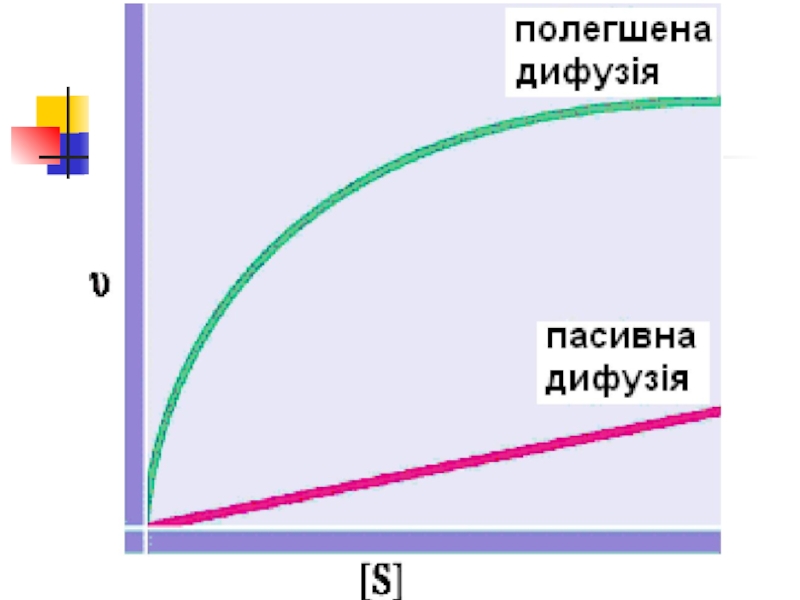
Слайд 91.2 Пасивний транспорт.Полегшена дифузія
Полегшена дифузія, порівняно із простою, є процесом більш

Слайд 112. Іонні канали
За механізмом відкриття виділяють кілька типів мембранних каналів:
•
• потенціал-залежні – регулюються при деполяризації ПМ у клітинах, здатних до збудження (нейрони, м’язові клітини);
• механочутливі - напр., відкриття механічними впливами звукових хвиль
іонних каналів клітин внутрішнього вуха, що спричиняє виникнення нервового імпульсу, який надалі в мозку інтерпретується як звук
• світлочутливі (містяться на ПМ клітин сітківки ока).


Слайд 13Внутрішньоклітинними лігандами найчастіше є фосфатні залишки, що приєднуються до ОН-груп серину,


Слайд 15Відома низка сполук, здатних специфічно блокувати Na+-канали. Найвідомішими з таких речовин



Слайд 18Активатори
потенціалзалежних
Вератридин – алкалоїд стероїдної природи, міститься в рослинах родини лілейних (Veratrum album – чемериця біла)


Слайд 20Токсини цигуатера продукуються
джгутиковими Gamberdiscus,
коралових риб, з м’ясом яких потрапляють до
шлунково-кишкового тракту людини.

Слайд 21Продуцент бревітоксину – дінофлагелят
Gymnodinium brevis. Ця
накопичуватися в тілі коралових риб.



")
")
Слайд 272.4 Са2+ - канали
За механізмом відкриття розрізняють
потенціалзалежні й ліганд-залежні Са2+-канали.
Потенціалзалежні Са2+-канали залежно від субодиничного складу, провідності, швидкості активації та інактивації, специфічних блокаторів, локалізації поділяються на кілька типів. Так, є два великі класи цих структур - HVA (high voltage activated – ті, що активуються за умов сильної деполяризації) та LVA (low voltage activated – ті, що активуються за умов слабкої деполяризації) Ca2+-канали. HVA Ca2+-канали, у свою чергу, поділяються на канали L, N, P/Q і R- типи, тоді як до LVA Ca2+-каналів належать лише канали T-типу

Слайд 28Конотоксини як блокатори
α-конотоксин інгібує нікотиновий ацетилхоліновий рецептор у нервовій і м’язовій системі
δ- конотоксин інгібує
інактивацію
напругозалежних
натрієвих
каналів

Слайд 29
κ- конотоксин інгібує калієві канали
μ- конотоксин інгібує напругозалежні
ω- конотоксин інгібує N-тип напругозалежних кальцієвих каналів

Слайд 302.5 Хлорні канали
Біологічні мембрани різних типів клітин містять кілька видів каналів

Слайд 31Біоелектричні потенціали (біоструми) - електричні явища, що спостерігаються у живих клітинах
3. Електрична активність клітин різного типу
 - електричні явища, що спостерігаються у живих клітинах у спокої і при")

Слайд 33
(3.7)
(3.8)
k-коефіцієнт розподілу, показує, яку частину концентрація всередині складає від
(3.9)
(3.10)
 (3.8) k-коефіцієнт розподілу, показує, яку частину концентрація всередині складає від концентрації ззовні. Тоді рівняння")
Слайд 34Осмос – це дифузія молекул води через напівпроникну мембрану з області
2.2 Осмос.
Сила, яка викликає рух молекул розчинника (води), називається осмотичним тиском. За законом Вант-Гоффа:
(3.11)
де і – ізотопічний коефіцієнт, який показує, у скільки разів збільшується кількість розчинених часток при дисоціації молекул в електролітах; для неелектролітів і = 1;
С – концентрація розчиненої речовини; R – газова стала;
T – абсолютна температура розчину.

Слайд 35Рівняння осмосу аналогічне рівнянню (3.4):
(3.12)
де
Осмос відіграє значну роль в біологічних явищах. Наприклад, осмос зумовлює гемоліз еритроцитів у гіпотонічних розчинах. Вода доти проникає всередину клітини, доки гідростатичний тиск всередині клітини не зрівноважить осмотичний тиск. Внаслідок цього клітина набрякає, а мембрана розтягується.
Дія деяких послаблюючи лікарських препаратів грунтується на осмосі води в область підвищеної концентрації цього препарату в шлунку
:(3.12) де - швидкість переносу води через площу S;")
Слайд 36
Явище фільтрації відіграє важливу роль у переносі води через стінки кровоносних судин, тобто в процесах обміну водою між кров'ю і тканиною. Фільтрація плазми крові в ниркових нефронах приводить до утворення первинної сечі. При порушеннях обміну фільтрація веде до набряків в тканинах.
2.3 Фільтрація.
Фільтрація – це рух молекул води через пори у мембрані під дією градієнта статичного тиску. Швидкість фільтрації підлягає закону Пуазейля:
;
,
(3.13)
де u – об’єм фільтрованої води; t - час; x – гідравлічний опір пори радіусом R; h - в’язкість води;

Слайд 372.4 Дифузія іонів через мембрану
На мембрані існує різниця потенціалів, так звані
В загальному випадку перерос іонів через мембрану визначається двома факторами: нерівномірністю їх розпаду і дією електричного поля мембрани, тобто наявністю двох градієнтів - концентраційного
і електричного
.
Підставимо (3.2) в (3.1) і маємо рівняння Нернста-Планка:
(3.14)
Іншу формулу рівняння Нернста-Планка можна одержати, якщо згадати співвідношення між коефіцієнтом дифузії D і абсолютного температурою:
Оскільки можна вважати електричне поле в мембрані постійним
, де φм - різниця потенціалів на мембрані,
- її товщина.
Тоді:
(3.15)

Слайд 38
ми тут, не приводимо, можна рівнянню (12) надати вигляду:
Позначимо
(3.16)
Вибіркова проникність мембрани залежить від того, яка густина фіксованих зарядів одного знаку в порі. Якщо вона велика, то добре проникають іони протилежного знаку і погано однойменні.
Іони H+ і OH- практично в клітину не проникають, оскільки вони адсорбуються поверхнею мембрани, заряджають її і відштовхують інші іони. Проникнення іонів з розчинів підлягає правилу: чим вищий ступень дисоціації, тим менша проникність іонів. Однобічна проникність електролітів та їх нагромадження в клітині зумовлені різницею між вмістом клітини і зовнішнім середовищем.

Слайд 39
2.5 Полегшена дифузія.
Дифузія речовини чераз пори (канали) також описується дифузійними рівняннями наявність каналів суттєво збільшує проникність мембрани, ефективний коефіцієнт проникності мембрани в цьому випадку задежить від числа каналів на 1см2 площі мембрани n, радіуса канала r , його довжини
(3.17)
Ще одне "полегшення" дифузії зв'язане з молекулами-переносниками (Р). Механізм такої дифузії визначається дифузією через мембрану комплексу переносника з речовиною (Ax) в одному напрямі і дифузією вільного переносника в протилежному напрямі.
Аx - речовина зовнішнього середовища. При хімічній спорідненості А і Х утворюється комплекс АХ. Схема полегшеної дифузії показана на рис.3.5.



Слайд 42Рис.3.6.
Іонофори є двох видів: переносники і каналоформери. Валіноміцин - переносник. Молекули

Слайд 432.6 Активний транспорт.
Тепер можна впевнено сказати, що механізми активного транспорту іонів
Безсумнівно, що в транспорті іонів беруть участь компоненти мембрани і насамперед фермент Na+, K+ АТФ-аза є основною структурою транспортного механізму, який дістав назву натрій-калієвої помпи і здійснює нагромадження в клітині калію і виведення з клітини натрію.

Слайд 44Рис.3.7
К 3
+
Na+, K+- АТФ-аза - це великий
Її молекула складається з двох субодиниць - великої, a-ліпопротеїда і меншої, b - глікопротеїда.
a - поліпептид виявляє каталітичну активність і може фосфоризуватися в
b - глікопротеїд не здатний каталізувати гідроліз АТФ, але виявляє Na -специфічну іонофоретичну активність. Являючись інтегральним мембранним білком Na+, K+ АТФ-аза пронизує мембрану наскрізь і тому частини молекули виступають як в середину клітини ( a одиниця), так і за межі клітини ( b - од.) Активний центр молекули, відповідальний за гідроліз АТФ, повернутий всередину клітини і локалізований на a - поліпептидові(рис.3.7.).
присутності АТФ.

Слайд 45L+ Na+
цитоплазма
Рис.3.8.
Кількісні взаємовідношення в АТФ-азній
АТФ: Na+: K+ = 1:2:3
Отже клітину покидає більше позитивно заряджених частинок, аніж акумулюється в ній. Це створює надлишковий від'ємний заряд всередині клітини і мембранний потенціал на її поверхні.
Міжклітинне середовище
Хоча конкретні механізми активного транспорту ще точно невідомі, запропоновано багато гіпотез , які пояснюють його. Частина гіпотез грунтується на уявленнях про участь в транспорті спеціальних переносників.

Слайд 46Згідно з однією з таких гіпотез Na+ переноситься з переносником Y.
).

Слайд 47Сучасні уявлення про механізм дії натрій-калієвої помпи будуються на уявленнях про






