- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Спинной мозг. Физиология ствола мозга презентация
Содержание
- 1. Спинной мозг. Физиология ствола мозга
- 2. Сегменты спинного мозга 8 шейных (C1
- 3. ПРИНЦИП МЕТАМЕРИИ
- 4. Принципы иннервации спинного мозга: Сегментарный принцип
- 5. Закон Белла - Мажанди Вентральные (передние) корешки
- 6. Нейроны спинного мозга (13 млн.): 1. Двигательные
- 7. Нейроны. В сером веществе спинного мозга находятся
- 8. По эффекторному действию: Возбуждающие (3%) Тормозные
- 9. ФУНКЦИИ СПИННОГО МОЗГА:
- 10. Основные рефлексы спинного мозга Рефлексы растяжения (миотатические)
- 11. Регуляция двигательной активности спинным мозгом Регуляция фазной
- 12. Мышцы содержат 3 типа рецепторов растяжения: Певичные и вторичные окончания Сухожильные рецепторы
- 13. Первичные афференты реагирует на степень и скорость
- 14. 1. Миотатические рефлексы (сухожильные, Т-рефлексы, от тендон)
- 16. Облегчение Т–рефлексов. Если коленный и другие Т–рефлексы
- 17. 2. Постуральные рефлексы (от английск. postur—поза, положение),
- 18. ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ
- 19. Участие спинного мозга в регуляции произвольных движений
- 20. 1. Рефлексы мышц-антагонистов При стимуляции мышечных волокон одной мышцы происходит одновременное торможение мышцы-антагониста.
- 21. 2. Сгибательные рефлексы (защитные) 1. Рефлекторная дуга
- 22. 3. Перекрестный сгибательно-разгибательный рефлекс
- 23. Функции сухожильных органов. Рефлекторная дуга сухожильных органов
- 24. Активация телец Гольджи приводит к торможению α-мотонейронов
- 25. Спинальная локомоция (автоматизмы) Основные характеристики локомоции, т.е.
- 26. Активность центров координируется проприоспинальными
- 27. Предполагают, что у
- 28. Шагательный автоматизм Поднимите малыша под мышки (
- 29. СОСАТЕЛЬНЫЙ РЕФЛЕКС
- 30. Автоматизм Бауэра Когда ребенок лежит
- 31. Рефлекс Бабинского Грудничок разводит пальчики на ногах
- 32. Рефлекс Моро Положите малыша на
- 33. РЕФЛЕКС ГАЛАНТА Проведите пальцем вдоль позвоночника
- 34. Хватательный автоматизм Робинсона. Угасает этот рефлекс к
- 35. Полная параплегия возникает при разрыве спинного
- 36. В последующие недели и месяцы двигательные рефлексы восстанавливаются.
- 38. СПИНАЛЬНЫЙ ШОК Обратимое угнетение двигательных и вегетативных
- 39. Патологические кистевые рефлексы Сгибательные: Рефлекс
- 40. Патологические стопные рефлексы Разгибательные: Рефлекс
- 41. Сухожильные (миотатические) рефлексы спинного мозга
- 42. Сгибательные: Нижний рефлекс Россолимо — сгибание
- 43. Частичная параплегия - синдром Броун-Секара
- 44. Зоны поражения у человека при повреждении спинного мозга на уровне грудных сегментов (синдром Броун-Секара)
- 45. Вегетативные (интероцептивные ) рефлексы спинного мозга
- 46. Висцеромоторные рефлексы Проявляются в двигательных реакциях
- 48. Зоны Захарьина-Геда
- 50. Афферентная функция Восприятие возбуждений, поступающих от афферентных
- 51. ПРОВОДЯЩИЕ СИСТЕМЫ СПИННОГО МОЗГА: ВОСХОДЯЩИЕ ПУТИ
- 52. Тонкий пучок Голля (fasciculus gracilis) - от
- 53. Латеральный кортикоспинальный пирамидный тракт - двигательные зоны
- 54. ПРОДОЛГОВАТЫЙ МОЗГ
- 55. Функции продолговатого мозга Рефлекторная Проводниковая Тоническая
- 56. Ядра продолговатого мозга 1. ЯДРА ЧЕРЕПНОМОЗГОВЫХ НЕРВОВ
- 57. IX пара - ЯЗЫКОГЛОТОЧНОГО НЕРВА:
- 58. ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ
- 59. Ядра продолговатого мозга - 2 ПЕРЕКЛЮЧАЮЩИЕ ЯДРА:
- 60. РЕФЛЕКСЫ ПРОДОЛГОВАТОГО МОЗГА Жизненно-важные рефлексы Защитные рефлексы Рефлексы поддержания позы Вегетативные рефлексы Вестибуло-вегетативные рефлексы
- 61. Центры продолговатого мозга ЖИЗНЕННОВАЖНЫЕ:
- 62. Постуральные рефлексы (поддержания позы) СТАТИЧЕСКИЕ - от
- 63. Постуральные (статические) рефлексы (Р. Магнус): 1. шейные
- 64. 2. вестибулярные тонические рефлексы связаны с возбуждением
- 65. Мост В мосту расположены ядра ЧМН:
- 66. Средний мозг
- 67. Основные ядра среднего мозга Ядра черепномозговых нервов:
- 68. Схема расположения двигательных центров в стволе мозга (продолговатом мозге, мосте и среднем мозге).
- 69. Двигательные рефлексы среднего мозга: СТАТИЧЕСКИЕ - от
- 70. СТАТОКИНЕТИЧЕСКИЕ – от рецепторов полукружных каналов - рефлексы прямолинейного ускорения - рефлексы углового ускорения
- 71. Красное ядро Участвует в поддержании мышечного тонуса.
- 72. децеребрационная ригидность В случае перерезки головного мозга
- 73. Механизм: ядро Дейтерса находится
- 74. Чёрная субстанция Тесно связана с четверохолмием, красным
- 75. медиальный продольный пучок В среднем мозге проходит
- 77. водопровод среднего мозга Водопровод среднего мозга окружает
- 78. четверохолмие Верхние бугры являются подкорковыми центрами зрительного
- 79. Ретикулярная формация среднего мозга
- 80. Ретикулярная формация Филогенетически древняя
- 81. Неспецифичность системы Ретикулярная формация — комплекс полисинаптических путей.
- 82. Проекции восходящей ретикулярной активирующей системы
- 83. Благодаря работам Г. Мегуна и Дж. Моруцци
- 84. Уровень активности ретикулярной формации и, следовательно, уровень
- 85. Стволовые центры, выделяющие нейрогормоны
- 86. Голубое пятно и норадреналиновая система. Нервные волокна
- 87. Ядра шва и серотонинергическая система.
- 88. Основные системы ретикулярной формации АФФЕРЕНТНЫЕ СИСТЕМЫ: от
- 89. Схема облегчающих (+) и тормозящих (-) зон
- 90. Афферентные и эфферентные связи ретикулярной формации стволовой части мозга
- 91. Таламус является коллектором практически всех афферентных путей

Слайд 2Сегменты спинного мозга
8 шейных (C1 - C8)
12 грудных (Th1 - Th12)
5
5 крестцовых (S1-S5)
1-3 копчиковых (Co1 - Co2)
12 грудных (Th1 - Th12)5 поясничных (L1 - L5)5")

Слайд 4Принципы иннервации спинного мозга:
Сегментарный принцип
Морфологических границ между сегментами нет, поэтому
Каждый сегмент спинного мозга иннервирует строго определённый отрезок тела: кожу и мышцы, называемый метамером или дерматомом.
Межсегментарный принцип
Каждый дерматом иннервируется одновременно тремя корешками.

Слайд 5Закон Белла - Мажанди
Вентральные (передние) корешки содержат эфферентные двигательные (выходящие) волокна,
 корешки содержат эфферентные двигательные (выходящие) волокна, а дорсальные (задние) корешки")
Слайд 6Нейроны спинного мозга
(13 млн.):
1. Двигательные или мотонейроны (3%):
- альфа-мотонейроны
- гамма-мотонейроны
2. Вставочные или интернейроны:
- собственные спинальные или проприоспинальные (нейроны собственного аппарата спинного мозга, устанавливают связи внутри и между сегментами)
- проекционные (реагируют на афферентные раздражители)
3. Вегетативные нейроны
:1. Двигательные или мотонейроны (3%):")
Слайд 7Нейроны. В сером веществе спинного мозга находятся тела двигательных, вставочных и
1) Мотонейроны. Различают крупные, или α‑мотонейроны, и мелкие, или γ‑мотонейроны.
α–Мотонейроны входят в состав медиальных и латеральных ядер. Это наиболее крупные клетки спинного мозга.
γ–Мотонейроны иннервируют интрафузальные волокна мышечных веретён.
2) Вставочные нейроны получают информацию от одних нейронов и передают её другим. Аксоны вставочных нейронов участвуют в образовании проводящих путей.
Клетки Реншоу регистрируют сигналы от возвратной ветви аксонов α‑мотонейронов. Аксоны клеток Реншоу образуют тормозные синапсы с перикарионами этих мотонейронов
 Мотонейроны. Различают")
Тормозные")

Слайд 10Основные рефлексы спинного мозга
Рефлексы растяжения (миотатические) - в основном разгибательные -
Сгибательные рывковые рефлексы (защитные)
Ритмические рефлексы (чесательный, шагательный) или автоматизмы.
Позные рефлексы (шейные тонические рефлексы наклонения и положения)
Вегетативные рефлексы
Висцеромоторные рефлексы
 - в основном разгибательные - рефлексы позы, толчковые (прыжок,")
Слайд 11Регуляция двигательной активности спинным мозгом
Регуляция фазной активности мышц:
Сгибательные рефлексы
Локомоции (автоматизмы)
Регуляция тонуса
мышц:
Миотатические
Позно-тонические рефлексы (постуральные)
Регуляция тонусамышц:Миотатические рефлексыПозно-тонические рефлексы (постуральные)")
Слайд 12Мышцы содержат 3 типа рецепторов растяжения:
Певичные и вторичные окончания
Сухожильные рецепторы

Слайд 13Первичные афференты реагирует на степень и скорость растяжения мышц (динамику), а
, а вторичное — только на")
Слайд 141. Миотатические рефлексы (сухожильные, Т-рефлексы, от тендон) – рефлексы на растяжение
Вызываются ударом по сухожилию мышцы. При таком растяжении мышцы – активируются интрафузальные мышечные веретёна. Импульсация от них по Iа афферентам попадает на α-мотонейроны данной мышцы.
В результате происходит укорочение экстрафузальных волокон, тем самым мышца возвращается к исходной длине и восстанавливает базовый тонус.
 – рефлексы на растяжение мышцы. Вызываются")
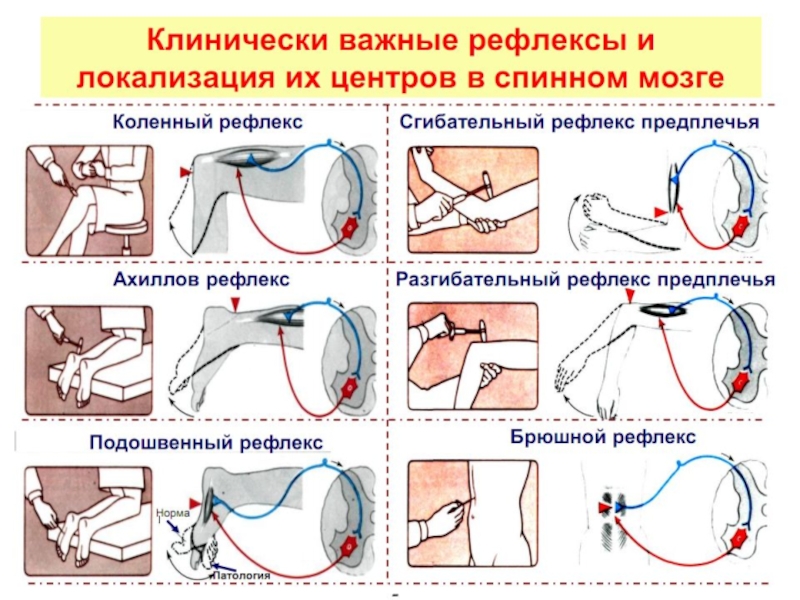
Слайд 16Облегчение Т–рефлексов.
Если коленный и другие Т–рефлексы нижней конечности ослаблены, их можно
Развиваемое при этом усилие облегчает активацию мотонейронов поясничного отдела спинного мозга.

Слайд 172. Постуральные рефлексы
(от английск. postur—поза, положение), термин, предложенный Шеррингтоном для обозначения
Постуральные (позотонические, статические) рефлексы обеспечивают поддержание в пространстве определённого положения всего тела или его части (например, конечности).
Познотонические рефлексы возникают с проприорецепторов мышц шеи, рецепторов фасции шеи животного. Переключение с этих рецепторов осуществляется на уровне шейного отдела спинного мозга и приводит к изменению тонуса мышц при изменении положения головы и шеи ( Магнус и Клейн).
, термин, предложенный Шеррингтоном для обозначения рефлексов, обеспечивающих сохранение определенного")

Слайд 19Участие спинного мозга в регуляции произвольных движений
От супраспинальных отделов управляющие сигналы
В результате при каждом мышечном сокращении происходит одновременное сокращение экстра– и интрафузальных МВ.
В этом случае даже в укороченной мышце – веретёна будут способны следить за длиной мышцы.

Слайд 201. Рефлексы мышц-антагонистов
При стимуляции мышечных волокон одной мышцы происходит одновременное торможение

Слайд 212. Сгибательные рефлексы
(защитные)
1. Рефлекторная дуга полисинаптическая.
2. Осуществляются при раздражении различных рецепторных
3. Этот рефлекс намного сложнее миототического – участвует не одна мышца.
Защитный сгибательный рефлекс.
1. Рефлекторная дуга полисинаптическая.2. Осуществляются при раздражении различных рецепторных полей: кожных рецепторов,")

Слайд 23Функции сухожильных органов.
Рефлекторная дуга сухожильных органов служит для поддержания постоянства напряжения мышцы.
У
Влияние системы регуляции длины в принципе ограничивается одной мышцей и ее антагонистом, тогда как регуляция напряжения с участием афферентов Ib относится к мышечному тонусу всей конечности.

Слайд 24Активация телец Гольджи приводит к торможению α-мотонейронов и обеспечивает расслабление сокращенной
4. Рефлексы ограничивающие напряжение мышц

Слайд 25Спинальная локомоция
(автоматизмы)
Основные характеристики локомоции, т.е. перемещения человека или животного в окружающей
Болевое раздражение какой–либо конечности спинального животного вызывает рефлекторные движения всех четырех; если же такая стимуляция продолжается достаточно долго, могут возникнуть ритмичные сгибательные и разгибательные движения не подвергающихся раздражению конечностей.
Если такое спинальное животное поставить на тредбан, то при некоторых условиях оно будет совершать координированные шагательные движения, весьма сходные с естественными.
Основные характеристики локомоции, т.е. перемещения человека или животного в окружающей среде при помощи")
Слайд 26 Активность центров координируется проприоспинальными системами и трактами, пересекающими
У более примитивных животных спинальные локомоции могут проявляться - в беге без головы.

Слайд 27
Предполагают, что у человека тоже есть спинальные локомоторные
По–видимому, их активация при раздражении кожи проявляется в виде шагательного рефлекса новорожденного ( сохраняется до 2-х месяцев).
Однако, по мере созревания центральной нервной системы супраспинальные отделы, подчиняют себе такие центры и у взрослого человека они утрачивают способность к самостоятельной активности.

Слайд 28Шагательный автоматизм
Поднимите малыша под мышки ( стопы должны лишь прикасаться к
. Затем слегка")

Слайд 30Автоматизм Бауэра
Когда ребенок лежит на животе, подставьте свою ладонь ему

Слайд 31Рефлекс Бабинского
Грудничок разводит пальчики на ногах веером лишь тогда, когда вы

Слайд 32Рефлекс Моро
Положите малыша на спинку и хлопните ладонями по кровати

Слайд 33РЕФЛЕКС ГАЛАНТА
Проведите пальцем вдоль позвоночника с правой стороны ( на
.")
Слайд 34Хватательный автоматизм Робинсона.
Угасает этот рефлекс к 4-му месяцу. А на смену ему

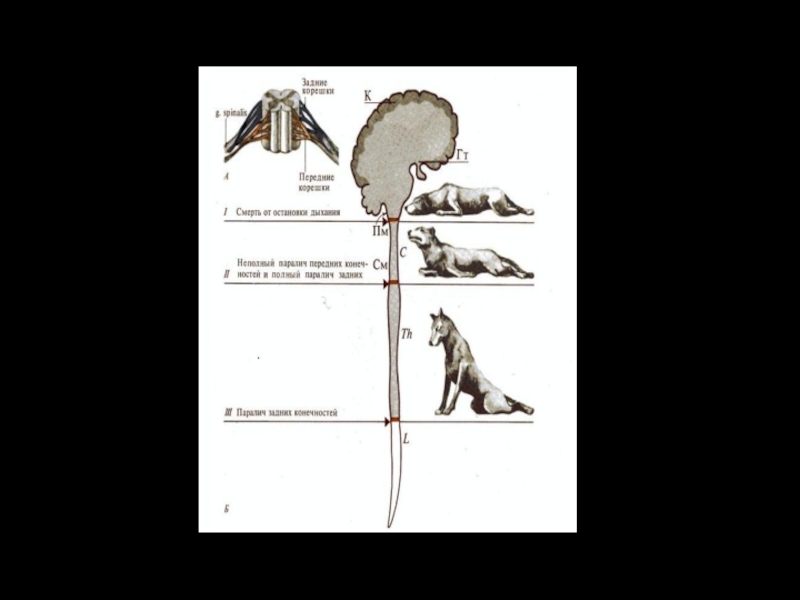
Слайд 35Полная параплегия возникает при разрыве спинного мозга в грудном отделе– от Т2
1) мгновенным и окончательным параличом всех произвольных движений мышц, каудальнее места повреждения;
2) полной и окончательной потерей осознаваемой чувствительности областей тела, соответствующих этим сегментам;
3) временной полной арефлексией, т.е. временным исчезновением всех двигательных и вегетативных рефлексов в тех же областях тела.

Слайд 36 В последующие недели и месяцы двигательные рефлексы восстанавливаются.
полная арефлексия (обычно 4–6 недель);
небольших рефлекторных движений пальцев ног, в первую очередь–большого (от двух недель до нескольких месяцев);
постепенное усиление сгибательных рефлексов–сначала большого пальца ноги (рефлекс Бабинского) и голеностопного сустава, затем колена и бедра.
хроническая стадия (через шесть и более месяцев)
Сгибательные рефлексы преобладают. Но появляется, также, генерализация разгибательных - до спазмов (спинальное стояние).
Если разгибательные движения появляются раньше, то есть надежда на неполное прерывание спинного мозга.
Последовательность включения рефлексов: сгибательные, сухожильные, вегетативные.

Слайд 38СПИНАЛЬНЫЙ ШОК
Обратимое угнетение двигательных и вегетативных рефлексов после разрыва спинного мозга
Главный фактор его возникновения – утрата связей с остальными отделами центральной нервной системы.
Перерезка нисходящих путей отключает множество возбуждающих входов спинальных нейронов; при этом, тормозные спинальные интернейроны растормаживаются.
Оба эти фактора приведут к сильному подавлению активности рефлекторных дуг, клиническим проявлением которого будет арефлексия.

Слайд 39
Патологические кистевые рефлексы
Сгибательные:
Рефлекс Жуковского — сгибание пальцев кисти при ударе молоточком по
Рефлекс Бехтерева запястно-пальцевой — сгибание пальцев руки при перкуссии молоточком тыла кисти.
Верхний рефлекс Россолимо — сгибание пальцев кисти при быстром касательном ударе по их подушечкам.

Слайд 40
Патологические стопные рефлексы
Разгибательные:
Рефлекс Оппенгейма — разгибание I пальца стопы при проведении
Рефлекс Гордона — медленное разгибание I пальца стопы и веерообразное расхождением других пальцев при сдавлении икроножных мышц.
Рефлекс Шефера — разгибание I пальца стопы при сдавлении ахиллова сухожилия.
Рефлекс Бабинского — разгибание I пальца стопы при штриховом раздражении кожи наружного края подошвы.

 рефлексы спинного мозга")
Слайд 42Сгибательные:
Нижний рефлекс Россолимо — сгибание пальцев стопы при быстром касательном ударе
Рефлекс Бехтерева –Менделя — сгибание пальцев стопы при ударе неврологическим молоточком по её тыльной поверхности.
Рефлекс Жуковвского-Корнилова — сгибание пальцев стопы при ударе неврологическим молоточком по её подошвенной поверхности непосредственно под пальцами.
Рефлекс Бехтерева— сгибание пальцев стопы при ударе неврологическим молоточком по подошвенной поверхности пятки.


Слайд 44Зоны поражения у человека при повреждении спинного мозга на уровне грудных
")
 рефлексы спинного мозга")
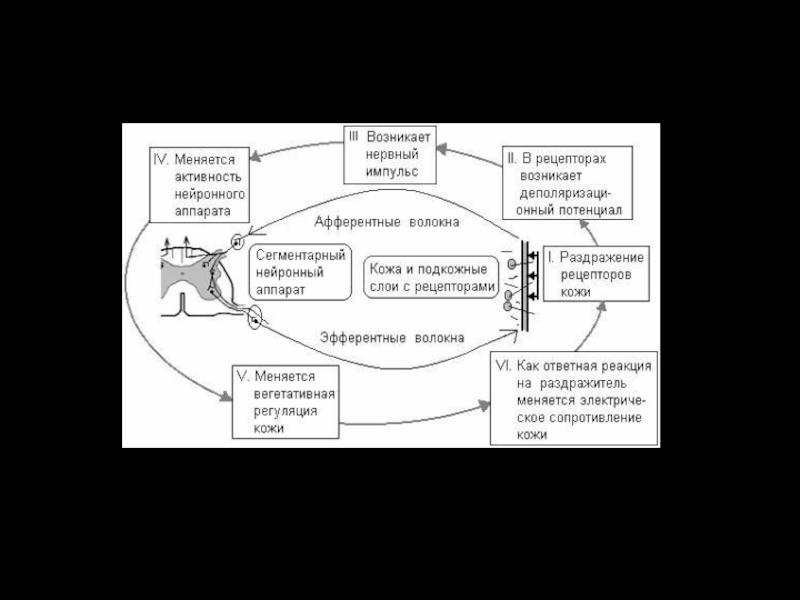
Слайд 46Висцеромоторные рефлексы Проявляются в двигательных реакциях мышц грудной клетки и брюшной
Рефлексы вегетативной нервной системы
Обеспечивают реакцию внутренних органов, сосудистой системы на раздражение висцеральных, мышечных кожных рецепторов
(Зоны Геда, рефлекс Гольца и др.)

Слайд 47
рефлекс Гольца
раздражение - удар в эпигастральную область, реакция— рефлекторная остановка сердца


Слайд 50Афферентная функция
Восприятие возбуждений, поступающих от афферентных клеток в ответ на раздражение

Слайд 51ПРОВОДЯЩИЕ СИСТЕМЫ
СПИННОГО МОЗГА:
ВОСХОДЯЩИЕ ПУТИ (ЭКСТЕРО- ПРОПРИО-ИНТЕРОЦЕПТИВНАЯ ЧУВСТВИТЕЛЬНОСТЬ)
НИСХОДЯЩИЕ ПУТИ (ЭФФЕКТОРНЫЕ, ДВИГАТЕЛЬНЫЕ)
СОБСТВЕННЫЕ
НИСХОДЯЩИЕ ПУТИ (ЭФФЕКТОРНЫЕ, ДВИГАТЕЛЬНЫЕ)СОБСТВЕННЫЕ (ПРОПРИОСПИНАЛЬНЫЕ) ПУТИ (АССОЦИАТИВНЫЕ И")
Слайд 52Тонкий пучок Голля (fasciculus gracilis) - от нижней части тела -
Клиновидный пучок Бурдаха (fasciculus cuneatus) - от верхней части тела - те же рецепторы
Латеральный спиноталамический тракт - болевая и температурная чувствительность
Вентральный спиноталамический тракт - тактильная чувствительность
Дорсальный спинно-мозжечковый тракт Флексига - (дважды перекрещенный) - проприоцепция
Вентральный спинно-мозжечковый тракт Говерса - (неперекрещенный) - проприоцепция
ВОСХОДЯЩИЕ ПУТИ СПИННОГО МОЗГА:
 - от нижней части тела - проприоцепторы сухожилий и мышц,")
Слайд 53Латеральный кортикоспинальный пирамидный тракт - двигательные зоны коры - перекрест в
Прямой передний кортикоспинальный пирамидный тракт - перекрест на уровне сегментов - команды те же, что и у латерального тракта
Руброспинальный тракт Монакова - красные ядра - перекрест-интернейроны спинного мозга - тонус мышц-сгибателей
Вестибулоспинальный тракт - вестибулярные ядра Дейтерса - перекрест - мотонейроны спинного мозга - тонус мышц-разгибателей
Ретикулоспинальный тракт - ядра ретикулярной формации - интернейроны спинного мозга - регуляция тонуса мышц
Тектоспинальный тракт - ядра покрышки среднего мозга - интернейроны спинного мозга - регуляция тонуса мышц
Нисходящие пути спинного мозга



Слайд 56Ядра продолговатого мозга
1. ЯДРА ЧЕРЕПНОМОЗГОВЫХ НЕРВОВ
XII пара - ПОДЪЯЗЫЧНОГО НЕРВА -
XI пара - ДОБАВОЧНОГО НЕРВА - n. аccessorius - двигательные ядра
Х пара - БЛУЖДАЮЩЕГО НЕРВА - n. vagus:
1) вегетативное ядро
2) чувствительное ядро одиночного пучка
3) обоюдное ядро - двигательное глотки и гортани

Слайд 57IX пара - ЯЗЫКОГЛОТОЧНОГО НЕРВА:
1) двигательное ядро - рот
2) чувствительное ядро - вкус задней трети языка
3) вегетативное ядро - слюнные железы
На границе с мостом:
YIII пара - ВЕСТИБУЛОКОХЛЕАРНОГО НЕРВА
1) кохлеарные ядра
2) вестибулярные ядра - медиальное Швальбе,
латеральное Дейтерса, верхнее Бехтерева
 двигательное ядро - рот и глотка 2) чувствительное ядро")

Слайд 59Ядра продолговатого мозга - 2
ПЕРЕКЛЮЧАЮЩИЕ ЯДРА:
- Голля и Бурдаха -
- Ретикулярной формации- от коры и подкорко-вых ганглиев к спинному мозгу
- Оливарные ядра - от коры, подкорковых ядер и мозжечка к спинному мозгу и от спинного мозга к мозжечку, таламусу и коре; от слуховых ядер в средний мозг и четверохолмие

Слайд 60РЕФЛЕКСЫ ПРОДОЛГОВАТОГО МОЗГА
Жизненно-важные рефлексы
Защитные рефлексы
Рефлексы поддержания позы
Вегетативные рефлексы
Вестибуло-вегетативные рефлексы

Слайд 61Центры продолговатого мозга
ЖИЗНЕННОВАЖНЫЕ:
Слезоотделения
1) Дыхательный Чиха тельный
Кашлевой
2) Сердечно-сосудистый Мигательный
Рвотный
3) Пищеварения
Слюноотделения
Сосания
Жевания
Глотания

Слайд 62Постуральные рефлексы (поддержания позы)
СТАТИЧЕСКИЕ - от рецепторов преддверия
- рефлексы выпрямления (установочные)
СТАТОКИНЕТИЧЕСКИЕ - от рецепторов полу-кружных каналов
- рефлексы прямолинейного ускорения
- рефлексы углового ускорения
СТАТИЧЕСКИЕ - от рецепторов преддверия - рефлексы положения - рефлексы")
Слайд 63Постуральные (статические) рефлексы (Р. Магнус):
1. шейные тонические - запускаются при возбуждении
голова вниз – гипертонус разгибателей задних конечностей,
голова назад – гипертонус разгибателей передних конечностей,
голова вправо – гипертонус разгибателей правых конечностей,
голова влево – гипертонус разгибателей левых конечностей,
Любое отклонение головы вызывает движение глазных яблок в противоположном направлении.
(в чистом виде при разрушении вестибулярного аппарата, дающего дополнительную информацию о положении головы)
 рефлексы (Р. Магнус):1. шейные тонические - запускаются при возбуждении проприоцепторов мышц шеи:голова вниз")
Слайд 642. вестибулярные тонические рефлексы связаны с возбуждением рецепторов преддверия перепончатого лабиринта,
Они не зависят от положения головы относительно туловища, а зависят от положения головы в пространстве (без сгибания в шее).
Подразделяются на:
- вестибулошейные рефлексы отвечают за вертикальное положение головы.
- вестибулоспинальные рефлексы подстраивают положение конечностей под положение головы.
(в чистом виде при фиксации головы по отношению к туловищу или при выключении проприоцепторов шейных мышц новокаиновой блокадой).

Слайд 65Мост
В мосту расположены ядра ЧМН:
V пара - тройничный нерв,
VI
VII пара - лицевой нерв,
VIII пара – вестибулокохлеарный нерв.


Слайд 67Основные ядра среднего мозга
Ядра черепномозговых нервов:
--- III пара - глазодвигательного нерва --- IV пара - блокового нерва --- Ядро Даркшевича - продольный пучок среднего мозга, связывающий ядра глазодвигательного, блокового и отводящего нерва в единую систему
Непарное вегетативное ядро Якубовича-Эдингера - через цилиарный ганглий к мышцам радужки и ресничного тела
Ядра тектальной области: верхнее или переднее двухолмие - зрительные рефлексы; нижнее или заднее двухолмие - слуховые рефлексы - четверохолмие
Черная субстанция
Красные ядра

Слайд 68Схема расположения двигательных центров в стволе мозга (продолговатом мозге, мосте и
.")
Слайд 69Двигательные рефлексы среднего мозга:
СТАТИЧЕСКИЕ - от рецепторов преддверия - рефлексы выпрямления
переход животного из неестественной позы в обычное для него положение.
При падении –
- сначала за счет вестибулярного выпрямительного рефлекса восстанавливается нормальное положение головы - мордой вниз.
- затем изменение положения головы возбуждает проприоцепторы шейных мышц и они запускают шейный выпрямительный рефлекс, в результате которого вслед за головой туловище также возвращается в нормальное положение.
 переход животного из")
Слайд 70СТАТОКИНЕТИЧЕСКИЕ –
от рецепторов полукружных каналов
- рефлексы прямолинейного ускорения
- рефлексы углового

Слайд 71Красное ядро
Участвует в поддержании мышечного тонуса. Тесно связано с мозжечком, вестибулярными
Руброспинальный тракт активирует α-мотонейроны сгибателей и тормозит активность α-мотонейронов разгибателей.

Слайд 72децеребрационная ригидность
В случае перерезки головного мозга ниже красного ядра возникает децеребрационная

Слайд 73
Механизм:
ядро Дейтерса находится под тормозным влиянием красного ядра.
После перерезки ниже
Тормозное влияние на ядро Дейтерса оказывает и мозжечок, поэтому удаление мозжечка ведет к усилению децеребрационной ригидности.

Слайд 74Чёрная субстанция
Тесно связана с четверохолмием, красным ядром, ретикулярной формацией ствола мозга,
Участвует в поддержании пластического тонуса. Отвечает за эмоциональное поведение, точные движения, особенно, пальцев рук; регулируют акты жевания и глотания (патология – паркинсонизм)
Чёрный пигмент появляется в клетках к 2-3 годам, по химическому строению является дофамином, который по аксонам транспортируется в базальные ганглии.

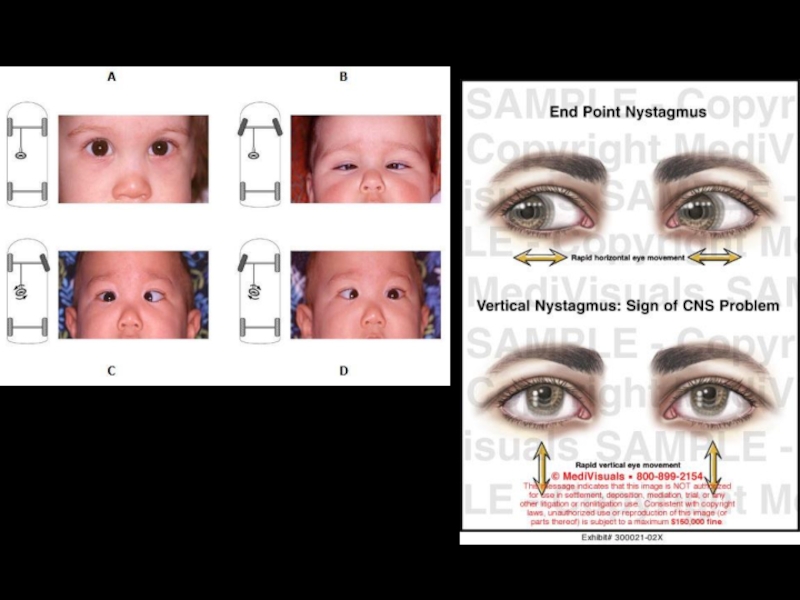
Слайд 75медиальный продольный пучок
В среднем мозге проходит медиальный продольный пучок , который
Этот пучок координирует движение глазных яблок, согласуя их с вестибулярными (вестибулярные ядра продолговатого мозга) и другими двигательными реакциями, благодаря чему взор удерживается на точке фиксации, несмотря на отклонение головы от обычного положения – явление нистагма.
При травмах среднего мозга или патологии вестибулярного аппарата возникает спонтанный нистагм – непроизвольное маятникообразное движение глазных яблок.

Слайд 77водопровод среднего мозга
Водопровод среднего мозга окружает центральное серое вещество, относящееся к

Слайд 78четверохолмие
Верхние бугры являются подкорковыми центрами зрительного анализатора.
Нижние бугры – слухового анализатора.
Связаны соответственно с латеральными и медиальными коленчатыми телами.
Четверохолмия являются центром безусловно-рефлекторных двигательных актов в ответ на зрительные и слуховые раздражения. При его участии формируются ориентировочные рефлексы на звук и свет.
Вегетативное обеспечение ориентировочных реакций получило название - старт-рефлексов: учащение сердечных сокращений, дыхания, повышение АД, тонуса мышц – всё это является предпосылкой для бегства или оборонительной реакции (осуществляется через гипоталамус).


Слайд 80Ретикулярная формация
Филогенетически древняя сердцевина мозга — занимает медиовентральную область
Она состоит из множества образующих комплексы сетевых структур (например, адренергических, серотонинергических и норадренергических нейронных систем).
В ретикулярной формации находятся области, связанные с регуляцией ритма сердца, АД, дыхания и других функций.
Некоторые её нисходящие тракты блокируют передачу импульсов в сенсорных проводниках спинного мозга и влияют на систему рефлексов растяжения.

Слайд 81Неспецифичность системы
Ретикулярная формация — комплекс полисинаптических путей. Аксоны входят в неё не
Поэтому большинство нейронов ретикулярной системы активируется с одинаковой степенью лёгкости различными сенсорными стимулами, в этом отношении ретикулярная формация является неспецифической.

Слайд 82Проекции восходящей ретикулярной активирующей системы
На рисунке приведена общая
Центральный компонент этой системы расположен в ретикулярной формации моста и среднего мозга, это бульборетикулярная облегчающая область.
Возбуждающие сигналы из этой области поступают прежде всего в таламус, где они возбуждают новую систему нейронов.

Слайд 83Благодаря работам Г. Мегуна и Дж. Моруцци наряду с неспецифическими нисходящими
Если через вживленные электроды раздражать центральные части ретикулярной формации ствола, то кошка, находящаяся в сонном состоянии, пробуждается. Эта поведенческая реакция пробуждения сопровождается характерными изменениями частотного спектра электроэнцефалограммы, переходом от регулярных, высоковольтных колебаний α—ритма к низковольтным колебаниям (β—ритма).
Данная электроэнцефалографическая реакция получила название реакции десинхронизации.

Слайд 84Уровень активности ретикулярной формации и, следовательно, уровень активности большого мозга в
Так, сигналы боли исключительно сильно возбуждают ретикулярную формацию мозгового ствола, повышая реакции внимания и тревоги.
Важность сенсорных сигналов для поддержания деятельного состояния ретикулярной формации ствола мозга демонстрируется пересечением мозгового ствола над местом вхождения в мозг V пары черепных нервов. Эти нервы входят в верхние отделы ствола мозга и передают значительное количество соматосенсорных сигналов в мозг.
Прекращение поступления сигналов в ретикулярную формацию ствола резко уменьшает её активность, что приводит организм в состояние комы (кома — тяжёлое бессознательное состояние). Когда ствол пересекают ниже места вхождения V пары черепных нервов, кома не возникает.


Слайд 86Голубое пятно и норадреналиновая система. Нервные волокна из этой области распространяются
Чёрное вещество и дофаминергическая система. Нервные волокна идут к хвостатому ядру и скорлупе и выделяют дофамин. Часть волокон направляется к гипоталамусу и лимбической системе. Чёрный пигмент (ДОФА) появляется в клетках к 2-3 годам. Разрушение дофаминергических нейронов чёрного вещества приводит к развитию болезни Паркинсона.

Слайд 87Ядра шва и серотонинергическая система.
Серотонин принимает участие в функционировании антиноцицептивной системы, механизмах сна и развитии ряда нейропсихиатрических расстройств.
Гигантоклеточные нейроны ретикулярной формации ствола и холинергическая система.
Нервные волокна идут вверх (к промежуточному мозгу и коре) и вниз (в спинной мозг).
В терминальных разветвлениях аксонов выделяется ацетилхолин, оказывающий возбуждающие эффекты.

Слайд 88Основные системы ретикулярной формации
АФФЕРЕНТНЫЕ СИСТЕМЫ:
от спинного мозга, от мозжечка, от четверохолмия,
от чувствительных ядер черепномозговых нервов
ЭФФЕРЕНТНЫЕ СИСТЕМЫ:
Восходящая активирующая система - неспецифическое тонизирование через синапсы на дентритах нейронов I и II слоев коры
Нисходящие ретикулоспинальные системы:
- облегчающая
- тормозящая

Слайд 89Схема облегчающих (+) и тормозящих (-) зон РФ и ее связи
ТОРМОЗЯЩИЕ ПУТИ:
1 – кортикоретикулярный,
2- каудатоспинальный,
3 –мозжечковретикулярный,
4 -ретикулоспинальный
ОБЛЕГЧАЮЩИЕ ПУТИ:
5 -спиноретикулярный,
6- вестибулоспинальный
 и тормозящих (-) зон РФ и ее связи с корой и подкоркойТОРМОЗЯЩИЕ")

Слайд 91Таламус является коллектором практически всех афферентных путей (исключение – обонятельный тракт).
Утратил эту функцию - с развитием коры головного мозга. Однако остался высшим центром болевой чувствительности.
Принимает участие в активации процессов внимания и в формировании эмоций (смеха, плача и др.).
Участвует в организации двигательных реакций: сосания, жевания, глотания; а также в обеспечении их вегетативным компонентом.
. На определённом этапе эволюции")





