клітини в культурі
б). Гени, що відповідають за туморогенез
1). Прото-онкогени та гени трансформуючих вірусів
функції прото-онкогенів
активація онкогена
2). Гени – супресори пухлинного росту
retinoblastoma
p53
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Онкогени та канцер презентация
Содержание
- 1. Онкогени та канцер
- 2. Взаємодія онкобілків ДНК-вірусів та клітинних білків
- 3. гени-супресори
- 4. Роль p53 в клітинному циклі G1
- 5. p53 “охоронець геному”
- 6. The p53 Signaling Pathway SIGMA-ALDRICH
- 7. Functional Domains of p53 MDM2 EMBO J
- 8. Ефект пошкодження ДНК та арешт клітинного росту p53 .
- 9. Механізми інактивації p53 p53 T ag
- 10. Клітинний цикл залежить від фосфорилювання Rb
- 11. роль pRB в регуляції клітинного поділу.
- 12. Гомологічні вірусні послідовності залучені у взаємодії з родиною білків Rb
- 13. Онкобілки взаємодіють з Rb Ad E1A HPV E7 SVL Tag Carcinogenesis, (2003) 24(2)159-169
- 14. Transformation via cell cycle control pathways
- 15. Апоптоз - (гр."falling"), притаманний багатоклітинному організму процес,
- 16. Біологічна суть апоптозу підтримка тканинного гомеостазу
- 18. Процес апоптозу умовно розділяють на: сигнальну
- 19. Шляхи апоптозу: ефектори та модулятори Існує два
- 21. Рецептори клітинної загибелі
- 22. Шляхи апоптозу: ефектори та модулятори Експресію Fas-R
- 23. Розвиток апоптозу може бути заблоковано активацією
- 24. Апоптоз, індукованний через TNF-α + TNF-R1,
- 25. Головний шлях апоптозу в клітинах ссавців «домен
- 26. Bcl-2 прото-онкоген вперше був описаний при
- 27. Регуляторна дія білків цієї родини здійснюється різними
- 28. Bcl2 та vBcl2 мають подібність
- 29. Bcl-2 Запуск апоптозу Через блок Bax Вірусні мімікріни Adenovirus EBV Herpesvirus CMV
- 30. Copyright ©2002 by the National Academy of
- 31. В результаті активації апоптичного сигналу, проапоптичні білки
- 32. Р53 та апоптоз При значних пошкодженнях ДНК
- 33. Сигнал смерті (радіація, мутація, хім.сполуки) P53 активується bax АПОПТОЗ Bcl2 (HHV8-vBcl2)
- 34. Inhibition of p53 functions (Fig. 18.21)
- 35. Hanahan та Weinberg запропонували, що нормальна клітина
- 37. Віруси та апоптоз Блокування вірусними білками апоптичного
Слайд 1 Онкогени та канцер
a). Багатоетапність канцерогенезу
1). Етапи еволюції пухлини
2). Ростові характеристики
. Багатоетапність канцерогенезу 1). Етапи еволюції пухлини 2). Ростові характеристики клітини в культурі б). Гени,")

Слайд 3 гени-супресори
наслідки при порушенні гені
гени
функція спадкові спорадичні
DCC (18q) взаємодія невідомі рак прямой
з кл поверхнею кишки
WT1 (11p) транскрипція Пухлина Вільямса Рак легенів
Rb1 (13q) транскрипція ретинобластома др-кл карцинома легенів
p53 (17p транскрипція Li-Fraumeni синдром рак мол.залози,
прямої кишки,легень
DCC (18q) взаємодія невідомі рак прямой
з кл поверхнею кишки
WT1 (11p) транскрипція Пухлина Вільямса Рак легенів
Rb1 (13q) транскрипція ретинобластома др-кл карцинома легенів
p53 (17p транскрипція Li-Fraumeni синдром рак мол.залози,
прямої кишки,легень

Слайд 4Роль p53 в клітинному циклі
G1
S
G2
M
G0
ДНК синтез
Ріст та
підготовка до
поділу
підготовка до
поділу
Стан спокою
фаза
фаза
фаза
фаза
мітоз
апоптоз
p53
пошкодження
Арешт кл.циклу

Слайд 5 p53 “охоронець геному”
в зародкових клітинах мутації р53 призводять
до
синдрому Li-Fraumeni
p53 мутації часто утворюються в пухлинах людей
Функція білку p53 транскрипційний фактор, що регулює клітинний цикл та репарацію ДНК
порушення цілісності ДНК викликає арешт клітинного циклу G1, що залежить від p53;
Клітини з мутантою формою p53 не можуть “зупинитись”
та переходять в S фазу та реплікують пошкоджену ДНК
синдрому Li-Fraumeni
p53 мутації часто утворюються в пухлинах людей
Функція білку p53 транскрипційний фактор, що регулює клітинний цикл та репарацію ДНК
порушення цілісності ДНК викликає арешт клітинного циклу G1, що залежить від p53;
Клітини з мутантою формою p53 не можуть “зупинитись”
та переходять в S фазу та реплікують пошкоджену ДНК


Слайд 7Functional Domains of p53
MDM2
EMBO J (99)18:1661
http://www.novocastra.co.uk/oapdgs.htm
HPV E6
Ad E1b p55
18:1661http://www.novocastra.co.uk/oapdgs.htmHPV E6Ad E1b p55")

Слайд 9Механізми інактивації p53
p53
T ag
p53
Tag
Стабілізує p53 в неактивній формі
p53
E6
p53
E6
E6AP
Ub
Ub
Ub
E6AP:
E3 Ub лігаза
p53
p53
E4
p53
p53
E1B
Перетворює р53 з активатора в репресор транскрипції
E1B

Слайд 10Клітинний цикл залежить від фосфорилювання Rb
G1
S
G2
M
G0
спокій
phase
phase
phase
phase
Rb
p
p
p
p
Rb
p
p
p
p
Rb
p
p
p
p
Rb
p
p
p
p
Rb
p
p
Точка рестрикції
Rb
p
p
фосфорилювання Rb
дозволяє клітинам
пройти точку
рестрикції та уввійти
у S фазу
гіперфосфорильований Rb
гіпофосфорильований Rb



 24(2)159-169")
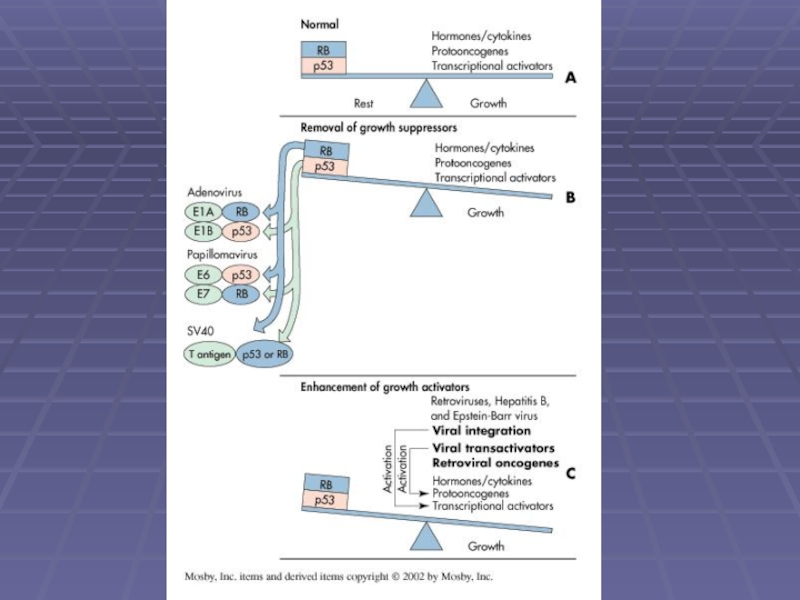
Слайд 14Transformation via cell cycle control pathways
Inhibition of Rb function by
viral proteins Many viruses actively inhibit Rb function
Result: bypass of restriction point control
Passage from G1 ? S phase
SV40 LT, adenovirus E1A, HPV E7 proteins
Result: bypass of restriction point control
Passage from G1 ? S phase
SV40 LT, adenovirus E1A, HPV E7 proteins

Слайд 15Апоптоз - (гр."falling"), притаманний багатоклітинному організму процес, за допомогою якого специфічні
клітини знищують та очищають організм від змінених клітин.
Kerr, J.F.R., Wyllie, A.H. and Currie, A.R. 1972. Br. J. Cancer 26:239.
We Thank Dave Cantrell from our Biomedical Communications, Arizona Health Sciences Center, for the graphic design and animation.
Apoptosis Society For Free Radical Biology and Medicine Tome & Briehl 2

Слайд 16Біологічна суть апоптозу
підтримка тканинного гомеостазу шляхом знищення надлишкових\ функціонально аномальних
клітин, а його інгібування – один з найважливіших механізмів онкогенезу.
Апоптоз інфікованих клітин індукується в результаті безпосередньої дії вірусних компонентів або внаслідок розпізнаванням їх клітинами імунної системи.
Апоптоз інфікованих клітин індукується в результаті безпосередньої дії вірусних компонентів або внаслідок розпізнаванням їх клітинами імунної системи.


Слайд 18Процес апоптозу умовно розділяють на:
сигнальну фазу, під час якої клітина
отримує сигнал, що ініціює апоптоз;
ефекторну фазу, коли активуються ефекторні внутрішньоклітинні механізми загибелі,
фазу деградації, при якій проходить деградація ДНК та інші зміни
ефекторну фазу, коли активуються ефекторні внутрішньоклітинні механізми загибелі,
фазу деградації, при якій проходить деградація ДНК та інші зміни

Слайд 19Шляхи апоптозу: ефектори та модулятори
Існує два основних шляху апоптозу в клітинах
ссавців
“Зовнішній” через «домен смерті»
(DD- death domain)
“Внутрішній” - мітохондріальний шлях
“Зовнішній” через «домен смерті»
(DD- death domain)
“Внутрішній” - мітохондріальний шлях

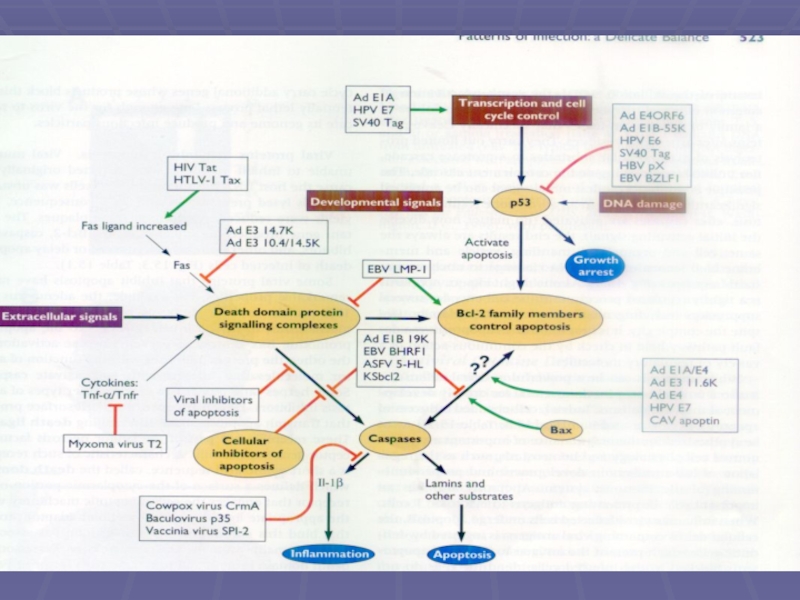
Слайд 21Рецептори клітинної загибелі
Fas-peцептор (Fas-R), TNF-R1, TNF-R2, "рецептор смерті-3" (DR-3
та 4 )
Найкраще вивчені Fas-R та TNF-R1.
Fas-R (APO-1/CD95) експресується на всіх ядерних клітинах
Існує 2 форми: звязаний з мембраною та в розчинній формі.
Розчинна форма Fas-R - для звязування Fas-ліганда (Fas-L) не тільки на СD8+-цитотоксичних лімфоцитах та NK-клітинах, але й
СD4+-Т-лімфоцитах-хелперах 1-го типу, які проявляють цитотоксичні властивасті
Найкраще вивчені Fas-R та TNF-R1.
Fas-R (APO-1/CD95) експресується на всіх ядерних клітинах
Існує 2 форми: звязаний з мембраною та в розчинній формі.
Розчинна форма Fas-R - для звязування Fas-ліганда (Fas-L) не тільки на СD8+-цитотоксичних лімфоцитах та NK-клітинах, але й
СD4+-Т-лімфоцитах-хелперах 1-го типу, які проявляють цитотоксичні властивасті
, TNF-R1, TNF-R2,")
Слайд 22Шляхи апоптозу: ефектори та модулятори
Експресію Fas-R на мембрані клітин індукують прозапальні
цитокіни ІЛ-1, -2, -6, ИФН-γ, фактори некрозу пухлин (TNF-α)
Запалення любої природи сприяє Fas-R-залежному пошкодженню органу (печінка).
Крім того, цитокіни стимулюють збільшення кількості молекул Fas-L на Т- та NK-лімфоцитах.
Запалення любої природи сприяє Fas-R-залежному пошкодженню органу (печінка).
Крім того, цитокіни стимулюють збільшення кількості молекул Fas-L на Т- та NK-лімфоцитах.

Слайд 23
Розвиток апоптозу може бути заблоковано активацією ряду факторів
I-FLICE (ендогенна домінантно-негативна
форма каспази 8), bcl-2 та Х-звязані інгібітори апоптозу
, bcl-2")
Слайд 24
Апоптоз, індукованний через TNF-α + TNF-R1, подібний Fas-R - Fas-L, та
потребує олігомерізації рецептору та здійснюється через шлях FADD - каспаза 8, а також подібний з ним білок TRADD.
Гіперекспресія TRADD призводить до запуску апоптозу та активації ядерного фактору кВ (NFKB), який запобігає TNF-індукованій загибелі клітин
Гіперекспресія TRADD призводить до запуску апоптозу та активації ядерного фактору кВ (NFKB), який запобігає TNF-індукованій загибелі клітин

Слайд 25Головний шлях апоптозу в клітинах ссавців
«домен смерті» (DD- death domain)
комлекс DISC
(death-inducing signaling complex)
.
адаптерний білок FADD
(Fas-associated DD)
білок FADD містить “ефекторний домен смерті” (DED, death effector domain)
аутоактивація – каспази 8
утворюються агрегати FasL-Fas-FADD-прокаспаза 8
.
адаптерний білок FADD
(Fas-associated DD)
білок FADD містить “ефекторний домен смерті” (DED, death effector domain)
аутоактивація – каспази 8
утворюються агрегати FasL-Fas-FADD-прокаспаза 8
комлекс DISC (death-inducing signaling complex). адаптерний")
Слайд 26Bcl-2 прото-онкоген вперше був описаний при B-клітинній лімфомі
Bcl 2
родина білків (17 представників) – модулятори апоптозу:
Bcl-xL, Mcl-1, Bag, A1, Bcl-2 - інгібітори апоптозу
Bcl-xS, Bax, Bad, Bid - промотори апоптозу
.
Bcl-xL, Mcl-1, Bag, A1, Bcl-2 - інгібітори апоптозу
Bcl-xS, Bax, Bad, Bid - промотори апоптозу
.
“Внутрішній” - мітохондріальний шлях
 –")
Слайд 27Регуляторна дія білків цієї родини здійснюється різними механізмами
-
утворення гомо- та гетеро-димерів
за допомогою наявності спільних доменів ВН1 та ВН2 з Вах і пригнічують апоптоз,
а домени ВН3 задіяні в комплексуванні з білками Bcl-2 та Bcl-xL, що сприяють розвитку апоптозу.
- білки цієї родини мають трансмембранний домен, який дозволяє їм локалізуватись на внутрішньоклітинних мембранах
Це сприяє зміні пронинкненості мембран та виходу цитохрому С та інших проапоптичних медіаторів з мітохондрій.
за допомогою наявності спільних доменів ВН1 та ВН2 з Вах і пригнічують апоптоз,
а домени ВН3 задіяні в комплексуванні з білками Bcl-2 та Bcl-xL, що сприяють розвитку апоптозу.
- білки цієї родини мають трансмембранний домен, який дозволяє їм локалізуватись на внутрішньоклітинних мембранах
Це сприяє зміні пронинкненості мембран та виходу цитохрому С та інших проапоптичних медіаторів з мітохондрій.

Слайд 28Bcl2 та vBcl2 мають подібність
Bcl2
E1B
vBcl2
(HHV8)
ASFV A179L
(African Swine
Fever virus)
BH4
BH3
BH1
BH2
TM
Bcl-2 запобігає апоптозу, блокуючи
вивільнення цитохрому С з мітохондрій
ASFV A179L(African SwineFever virus)BH4BH3BH1BH2TMBcl-2 запобігає апоптозу, блокуючи вивільнення цитохрому С з мітохондрій")

Слайд 30Copyright ©2002 by the National Academy of Sciences
Huang, Qiulong et al.
(2002) Proc. Natl. Acad. Sci. USA 99, 3428-3433
HHV-8 BcL-X Bcl-2
Подібність із BcL-2
 Proc. Natl. Acad.")
Слайд 31В результаті активації апоптичного сигналу, проапоптичні білки сприяють відкриттю мітохондріальних каналів
і виходу цитохрому С (вихід цитохрому С перешкоджає перетворенню каспази-9 в активну форму – відміняють апоптичний сигнал),
антиапоптичні білки–їх закривають.
Таким чином, апоптоз до певної міри залежить від співвідношення білків Вах та Bcl в мітохондріях.)
антиапоптичні білки–їх закривають.
Таким чином, апоптоз до певної міри залежить від співвідношення білків Вах та Bcl в мітохондріях.)

Слайд 32Р53 та апоптоз
При значних пошкодженнях ДНК активується експресія про - апоптичних
генів Вах, Fas, DR5 та інших,
р53 здатен пригнічувати експресію анти-апоптичних генів (наприклад, Bcl-2 та рецептору IGF-1).
При недостатності р53 та надлишку Всl-2 відбувається накопичення порушень в клітинах: подібні порушення спостерігаються при розвитку різноманітних пухлин.
р53 здатен пригнічувати експресію анти-апоптичних генів (наприклад, Bcl-2 та рецептору IGF-1).
При недостатності р53 та надлишку Всl-2 відбувається накопичення порушень в клітинах: подібні порушення спостерігаються при розвитку різноманітних пухлин.

 P53 активуєтьсяbaxАПОПТОЗBcl2 (HHV8-vBcl2)")
")
Слайд 35Hanahan та Weinberg запропонували, що нормальна клітина повинна отримати 6
фенотипових ознак для становлення злоякісності. Одна з них – резистентність до апоптозу. В цій моделі в хронологічному порядку викладені механізми, завдяки яким отримуються ці ознаки. Вони можуть варіювати для кожної пухлини. Генна нестабільність посилює набдання різних фенотипів
Апоптоз та рак
Hanahan, D. and Weinberg, R.A. 2000. Cell. 100:57.

Слайд 37Віруси та апоптоз
Блокування вірусними білками апоптичного сигналу і передачу його всередину
клітини ( аденовірусні E1B та Е3-10,4К, Е3-14,5 К, білок вірусу герпесу vFLIP)
Інгібуваня каспазної активності (Е3-14,7К)
Вірусні аналоги
Bcl-2 з антиапоптичною активністю (Е1В, білок герпес-вірус саймірі, SKHV- SKbcl-2, EBV- LMP1 , BHRF1)
Інгібування проапоптичної дії р53
(LТ-антиген SV-40, Е1А та Е1В-аденовірусів, Е6 та Е7 папіломавірусів, білки надранніх генів СМV- ІЕ1 та ІЕ2).
Інгібуваня каспазної активності (Е3-14,7К)
Вірусні аналоги
Bcl-2 з антиапоптичною активністю (Е1В, білок герпес-вірус саймірі, SKHV- SKbcl-2, EBV- LMP1 , BHRF1)
Інгібування проапоптичної дії р53
(LТ-антиген SV-40, Е1А та Е1В-аденовірусів, Е6 та Е7 папіломавірусів, білки надранніх генів СМV- ІЕ1 та ІЕ2).


Слайд 39
Fas L
DISC
FADD
DED
DED
DED
Caspase 8/10
3
3
3
Bid
3
3
Truncated Bid
Bid, Bad, Bax, Bag, Bik, Bok,
Bcl-xs and other
Active Caspase 8/10
Proapoptotic Bcl-2 Family Members
Heterodimerization of Bcl-2 Members

Слайд 40
Fas L
DISC
FADD
DED
DED
DED
Active Caspase 8/10
Caspase 8/10
Flip
IAPs
Effector-caspases 3.6.7
IAP
Crm A
Proteolysis of Death Substrates
-PARP
ICAD
CAD
ICAD
Active CAD
Active
P 53
89
24
DNA repair






