- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Гемопоэз. Органы кроветворения презентация
Содержание
- 1. Гемопоэз. Органы кроветворения
- 2. гемопоэз Более 30 млн клеток в минуту
- 3. гемопоэз Кроветворение (гемопоэз) - многостадийный процесс дифференцировки
- 5. Клетки, способные восстанавливать гемопоэз после облучения или
- 6. «Из стволовых клеток можно вырастить любой орган
- 7. способность к самообновлению способность к дифференцировке
- 8. Для лечения лейкозов и других заболеваний крови
- 9. Стволовые клетки Кроветорные СК Эмбриональные СК Стромальные (индуцированные) СК
- 10. Стволовые клетки взрослого организма мультипотентны или унипотентны.
- 11. Уникальное свойство эмбриональных стволовых клеток — тотипотентность
- 12. Костный мозг состоит из двух видов стволовых
- 13. Стромальные стволовые клетки костного мозга - универсальны.
- 14. Выделены уникальные вещества: особые белки, вызывающие перерождение
- 15. В 2012 г. в Стокгольме объявили лауреатов
- 16. Гемопоэз поддерживается в течение всей жизни за
- 18. Регуляция гемопоэза Позитивные: КСФ, ИЛ-6, ИЛ-11, ИЛ-12
- 19. Регуляция гемопоэза 1. Факторы, влияющие на ранние
- 20. Последовательность ультраструктурных изменений в процессе некроза и апоптоза
- 21. Молекулярные механизмы апоптоза. Выделяют два
- 22. Бурстстимулиру-ющая активность Витамин В12 , фолиевая кислота, железо, медь Эритропоэз
- 23. Иммунологический маркёр эритробластов – гликофорин А Дифференцировка
- 24. Синтез гемоглобина начинается на стадии проэритробласта В
- 25. Проэритробласт (эритробласт) Пронормобласт
- 26. Оксифильный эритробласт Базофильный (1) и полихрома- тофильный (2) эритробласты
- 27. Ретикулоциты
- 28. Распределение популяции ретикулоцитов по степени зрелости
- 29. Диаметр – 7,2-8,0 мкм Площадь поверхности –
- 30. Эритроциты в капилляре существенно меняют форму за
- 31. Схема мембраны эритроцита
- 32. Функции эритроцитов Участие в газообмене и поддержании
- 33. Гранулоцитопоэз В процессе созревания (в течение
- 34. Гранулогенез Первичные гранулы – азурофильные. Маркёр
- 35. Миелоидный росток Миелобласт Промиелоцит
- 36. Клетки нейтрофильного ростка
- 37. сегментоядерные нейтрофилы, Тельце Барра (1)
- 38. Клеточно-гуморальные контакты нейтрофилов
- 39. Основные этапы фагоцитоза – хемотаксис, адгезия антигена
- 40. Эозинофилы На мембране есть рецепторы к
- 41. Палочкоядерный эозинофил эозинофил
- 42. Базофил Тучная клетка
- 43. Базофилы В гранулах: гистамин, гистидин, гепарин,
- 44. Участие тучных клеток в реакции гиперчувствительности немедленного
- 45. Моноцитопоэз. Моноцитарный росток Монобласт Промоноцит
- 46. Моноцит Макрофаг
- 47. Цитохимические маркёры клеток моноцитарно-макрофагальной линии Неспецифическая эстераза,
- 48. Мононуклеарные фагоциты Обновление тканевых макрофагов происходит
- 49. Презентация антигена антигенпредставляющими клетками Макрофаги разделены
- 50. Фолликулярные дендритные клетки Локализация: В В-клеточных зонах
- 51. Интердигитальные дендритные клетки Локализация: В парокортикальных зонах
- 52. Биологическая роль СМФ в процессах обмена веществ
- 53. Секреторные факторы макрофагов Провоспалительные цитокины (ИЛ-1, ИЛ-6,
- 54. Роль цитокинов в воспалении. Активация эндотелиальных клеток,
- 55. Лимфоцитопоэз. Лимфоцитарный росток лимфобласт пролимфоцит лимфоцит
- 56. Большой гранулярный лимфоцит Плазмобласт
- 57. Проплазмоцит Плазмотическая клетка
- 58. Антигеннезависимые и антигензависимые этапы дифференцировки Т-лим- фоцитов
- 60. Дифференцировка В-лимфоцитов Основной характеристикой В-лимфоцитов является наличие
- 61. Механизм гуморального иммунитета: Интерферон Лизоцим +
- 63. Мегакариоцитарный росток мегакариобласт промегакариоцит мегакариоцит
- 64. Активированные тромбоциты на поврежденной сосудистой стенке.
- 65. Спасибо за внимание!

Слайд 2гемопоэз
Более 30 млн клеток в минуту образуется в кроветворных органах, а
Образующиеся в костном мозге клетки по мере созревания равномерно поступают в кровеносное русло, время циркуляции их постоянно - эритроциты циркулируют 110-130 суток, тромбоциты - около 10 суток, нейтрофилы - менее 10 ч.
Каждый день теряется 1 ×1011 клеток крови, но эти потери постоянно восполняются клеточной фабрикой - костным мозгом - в течение всей жизни человека.

Слайд 3гемопоэз
Кроветворение (гемопоэз) - многостадийный процесс дифференцировки клеточных элементов, в результате которого
Развитие кроветворения происходит со сменой преимущественной локализации его в различные периоды жизни человека, а каждый из кроветворных органов играет особую роль в размножении и созревании гемопоэтических клеток.
 - многостадийный процесс дифференцировки клеточных элементов, в результате которого образуются эритроциты, лейкоциты, тромбоциты,")
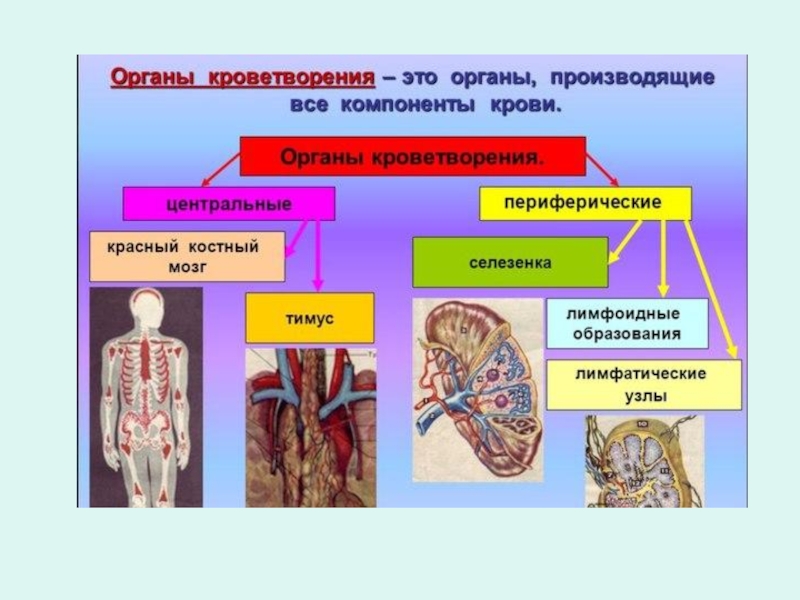
Слайд 5Клетки, способные восстанавливать гемопоэз после облучения или токсических воздействий, носят название
Впервые термин «стволовые
клетки» был введен в 1908 году
Максимов Александр Александрович
(04.02.1874 – 04.12.1928)

Слайд 6«Из стволовых клеток можно вырастить любой орган ?»
Орган вырастить нельзя
Мифы и реальность использования «стволовых клеток»
«Стволовые клетки – источник вечной молодости?»
Вечной молодости не будет, но можно улучшить состояние кожи (на время)
?

Слайд 7способность к самообновлению
способность к дифференцировке
количество стволовых клеток ограничено
Источники:
Костный мозг
Пуповинная
Периферическая кровь
Cтволовые клетки. Свойства.

Слайд 8Для лечения лейкозов и других заболеваний крови
При лечении рассеянного склероза
Стволовые
Стволовые клетки активно используются в тканевой инженерии.
Спектр применения гемопоэтических стволовых клеток расширяется с каждым днем, проводятся исследования в области ревматологии, (аутоиммунное заболевание ревматоидный артрит), кардиологии (ИБС), гепатологии, (токсические циррозы), онкологии, при ишемии конечностей. Даже разрабатывается подход к лечению СПИДа с применением стволовых клеток
Медицинское применение стволовых клеток

 СК")
Слайд 10Стволовые клетки взрослого организма мультипотентны или унипотентны.
А эмбриональные стволовые клетки плюрипотентны.
Стволовые
Эмбриональные стволовые клетки

Слайд 11Уникальное свойство эмбриональных стволовых клеток — тотипотентность
Проблема №1: как направить их
Проблема №2: иммунологическая несовместимость тканей трансплантата и реципиента
Проблема №3: онкогенность!
Проблема №4: этический барьер
«Уникальность и проблемы» эмбриональных стволовых клеток

Слайд 12Костный мозг состоит из двух видов стволовых клеток:
гемопоэтические стволовые клетки
стромальные
У новорожденного – на 10 тыс.КСК-1 стромальная СК
У подростков – на 100 тыс.КСК-1 стромальная СК
К 50 годам – на 500 тыс.КСК -1 стромальная СК
К 70 годам – на 1 млн КСК – 1 стромальная СК
Индуцированные (стромальные) стволовые клетки

Слайд 13Стромальные стволовые клетки костного мозга - универсальны. Они поступают с кровотоком
Собственные стромальные стволовые клетки не отторгаются, кроме того, при их введении низка вероятность их злокачественного перерождения.
Стромальные стволовые клетки костного мозга

Слайд 14Выделены уникальные вещества: особые белки, вызывающие перерождение стромальных клеток в клетки
Американские исследователи вырастили стволовые клетки мышечной ткани (миобласты) из бедренных мышц 72-летнего пациента-инфарктника. Затем эти клетки ввели ему непосредственно в зону инфаркта, после чего у больного было отмечено значительное улучшение сократительной способности сердца.
Стромальные клетки в клинической практике - это уже реальность.
. В")
Слайд 15В 2012 г. в Стокгольме объявили лауреатов Нобелевской премии по физиологии

Слайд 16Гемопоэз поддерживается в течение всей жизни за счёт «стволовых клеток»
Нормальное кроветворение
Клетки гемопоэза условно подразделены на 5-6 отделов, границы между которыми весьма размыты, а между отделами содержится много переходных, промежуточным форм.
В процессе дифференцировки происходит постепенное снижение пролиферативной активности клеток и способности развиваться сначала во все кроветворные линии, а затем во все более ограниченное количество линий.
Характеристики нормального гемопоэза

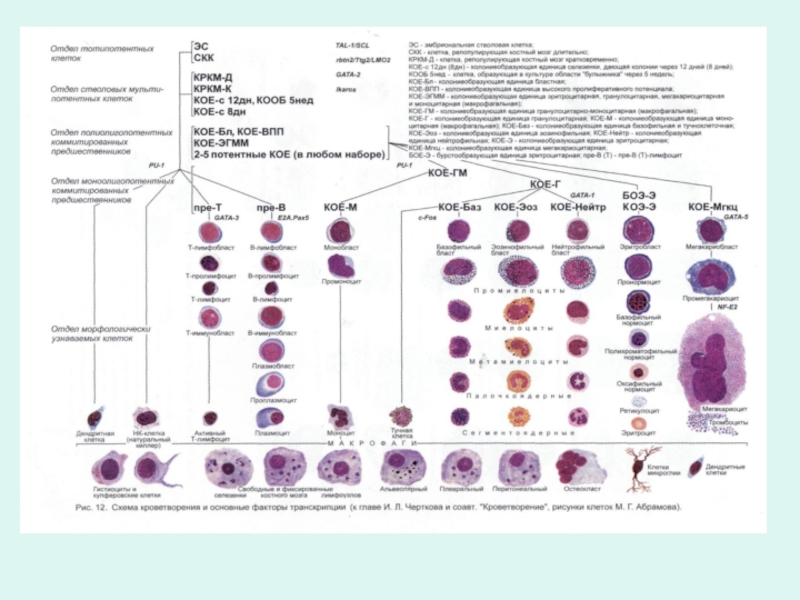
Слайд 18Регуляция гемопоэза
Позитивные: КСФ, ИЛ-6, ИЛ-11, ИЛ-12
Негативные: TGF-β, MIP-1α, ФНО-α, интерферон-α, интерферон-γ,

Слайд 19Регуляция гемопоэза
1. Факторы, влияющие на ранние СКК: фактор стволовых клеток (ФСК),
2. Линейно-неспецифические факторы - ИЛ-3, ИЛ-4, ГМ-КСФ (для гранулоцитомонопоэза).
3. Позднедействующие линейно-специфические факторы, которые поддерживают пролиферацию и созревание коммитированных предшественников и их потомков. К ним относятся эритропоэтин, тромбопоэтин, колониестимулирующие факторы (Г-КСФ, М-КСФ, ГМ-КСФ), ИЛ-5
, гранулоцитарный колониестимулирующий фактор (Г-КСФ),")

Слайд 21Молекулярные механизмы апоптоза.
Выделяют два типа сигнальных путей, вызывающих апоптоз, -
повреждение ДНК, радиация и другие факторы, вызывающие активацию каспазы-9
сигналы, возникающие при активации Fas-рецептора с
последующей активацией каспазы-8.
Каспазы9 и 8 активируют каспазу-3 и вместе с другими каспазами, протеазами, ДНК-азами вызывают апоптоз.


Слайд 23Иммунологический маркёр эритробластов – гликофорин А
Дифференцировка и созревание эритроидных клеток от
Способность к пролиферации сохраняется до ранних полихроматофильных эритробластов
В норме эритрокариоциты проходят 5 митозов, в результате чего из 1 эритробласта получается 32 эритроцита

Слайд 24Синтез гемоглобина начинается на стадии проэритробласта
В норме ранний полихроматофильный эритробласт подходит
Если количество гемоглобина более 27 пг на стадии базофильного эритробласта – завершение дифференцировки и апоптоз. Это – неэффективный эритропоэз. В норме 3-8% таких эритрокариоцитов (PAS-положительные эритрокариоциты).
При нехватке железа эритрокариоцит не достигает критической массы гемоглобина для деления и делится преждевременно, образуя микроциты.

Пронормобласт")
 и полихрома- тофильный (2) эритробласты")


Слайд 29Диаметр – 7,2-8,0 мкм
Площадь поверхности – 140мкм2
Объём – 90мкм3
Нормохромные
Нормоцит-нормальный эритроцит

Слайд 30Эритроциты в капилляре существенно
меняют форму за счет деформируемости
мембраны, что позволяет им
проходить
способность к газообмену с окружающей
тканью


Слайд 32Функции эритроцитов
Участие в газообмене и поддержании буферного кислотно-основного равновесия крови
Определяют реологию
Участвуют в гемостазе
Участвуют в иммунных процессах, взаимодействуя с циркулирующими иммунными комплексами
Адсорбируют токсины, липиды, аминокислоты
Источник оксидантной и антиоксидантной систем

Слайд 33Гранулоцитопоэз
В процессе созревания (в течение 10-13 дней) происходит:
Уменьшение ядра
Конденсация хроматина
Исчезновение
Сегментация ядра
Появление специфической зернистости (на стадии поздних миелобластов и промиелоцитов)
Утрата базофилии и увеличение объёма цитоплазмы
Цитохимические маркёры: МП, ЩФ, липиды, хлорацетатэстераза. PAS-положит. субстанция
 происходит:Уменьшение ядраКонденсация хроматинаИсчезновение ядрышекСегментация ядраПоявление специфической зернистости")
Слайд 34Гранулогенез
Первичные гранулы – азурофильные. Маркёр – миелопероксидаза. Содержат: лизоцим, катепсин,
Вторичные гранулы – базофильные. Маркёры: лактоферрин, катионный белок кателицидин, В12 – связывающий белок и др.
Третичные гранулы – желатиназа
Четвертичные гранулы – эндоплазматические органеллы: ЩФ, тетранектин, β2-микроглобулин.



")

Слайд 39Основные этапы фагоцитоза – хемотаксис, адгезия антигена к рецепторам мембраны фагоцита,
,")
Слайд 40Эозинофилы
На мембране есть рецепторы к Fc-фрагменту иммуноглобулинов, рецепторы для компонентов
В специф. гранулах – катионный белок и перекиси, кислая фосфатаза, коллагеназа, эластаза, катепсин, арилсульфатаза, простагландины и др.
Инактивируют БАВ из базофилов и тучных клеток
Обуславливают внеклеточный цитолиз, участвуя в противогельминтном иммунитете
Секретируют ИЛ-2, ИЛ-3, ФНО-α, ИЛ-4, ИЛ-5



Слайд 43Базофилы
В гранулах: гистамин, гистидин, гепарин, серотонин, ферменты, лейкотриены, тромбоксаны, простогландины,
На мембране есть рецепторы для комплемента, высокой плотности рецепторы к IgЕ.

Слайд 44Участие тучных клеток в реакции гиперчувствительности немедленного типа. ПГ - простагландины, ЛТ



Слайд 47Цитохимические маркёры клеток моноцитарно-макрофагальной линии
Неспецифическая эстераза, подавляемая фторидом натрия
Кислая фосфатаза
В процессе
Незначительное содержание липидов и гликогена

Слайд 48Мононуклеарные фагоциты
Обновление тканевых макрофагов происходит за счёт притока моноцитов крови
56%
15% - макрофаги в лёгких
15% - макрофаги в селезёнке
8% - в перитонеальной полости
6% - в других тканях

Слайд 49Презентация антигена антигенпредставляющими клетками
Макрофаги разделены на 2 касса:
Антигенперерабатывающие (профессиональные фагоциты)
Антигенпредставляющие
Антигенпредставляющие (дендритные клетки)")
Слайд 50Фолликулярные дендритные клетки
Локализация:
В В-клеточных зонах лимфоузлов
В селезёнке
В мукозо-ассоциированной лимфоидной ткани (MALT)
Представляют
ФДК – хранилище антигенов, обеспечивающие длительное поддержание иммунологической памяти
Представляют антиген В-лимфоцитамФДК – хранилище")
Слайд 51Интердигитальные дендритные клетки
Локализация:
В парокортикальных зонах лимфоузлов
В Т-зависимых зонах селезёнки, миндалин
В мозговом
Представляют антиген Т-лимфоцитам, вызывая антигенспецифический иммунный ответ

Слайд 52Биологическая роль СМФ в процессах обмена веществ
Участие в углеводном обмене (рецепторы
Активная роль в патогенезе атеросклероза (рецепторы для ЛПНП →образование «пенистых» клеток)
Метаболизм металлов: железа, цинка, меди

Слайд 53Секреторные факторы макрофагов
Провоспалительные цитокины (ИЛ-1, ИЛ-6, ИЛ-8, ИЛ-12, ИЛ-18, ФНО-α, интерферон-α,
Противовоспалительные цитокины (ИЛ-10, трансформирующий ростовой фактор)-иммуносупрессия, фиброз

Слайд 54Роль цитокинов в воспалении. Активация эндотелиальных клеток, повышение экспрессии молекул адгезии





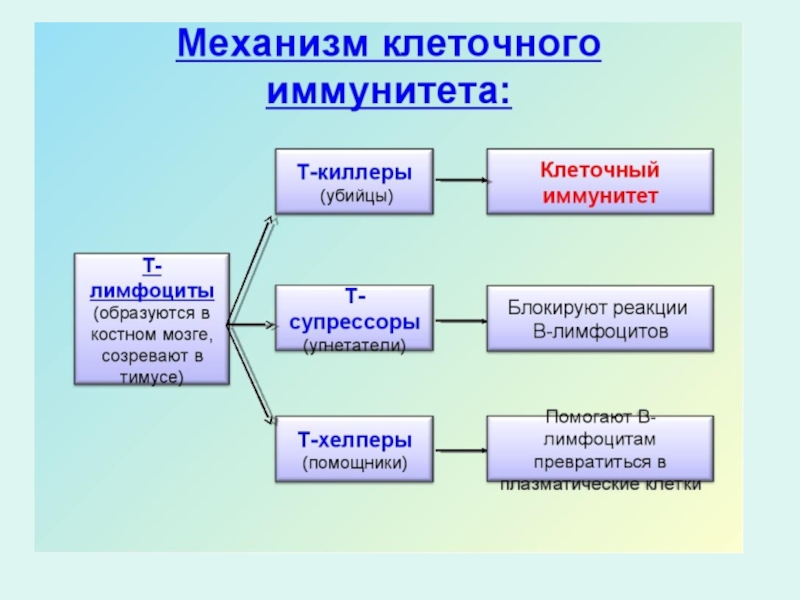
Слайд 60Дифференцировка В-лимфоцитов
Основной характеристикой В-лимфоцитов является наличие на их мембране рецепторов для
В-лимфоциты в костном мозге проходят этап антигеннезависимой дифференцировки-формирование зрелой (наивной) В-клетки. С момента завершения формирования рецепторного комплекса В-клетка приобретает способность взаимодействовать с антигеном
Зрелые В-лимфоциты покидают костный мозг, попадают в циркуляцию и поступают в периферические лимфоидные органы, где при встрече с антигеном они проходят этап антигензависимой дифференцировки.




Слайд 64Активированные тромбоциты на
поврежденной сосудистой стенке.
При активации происходит округление тромбоцитов
и образование
которых они способны агрегировать
между собой или адгезироваться на
поверхности







