- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Нуклеиновые кислоты презентация
Содержание
- 1. Нуклеиновые кислоты
- 2. Полимерные цепи нуклеиновых кислот построены из
- 3. Мономерное звено представляет собой трёхкомпонентное образование, включающее: гетероциклическое основание, углеводный остаток, фосфатную группу.
- 4. НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
- 5. Углеводы β-D-рибоза
- 6. Углеводы 2-дезокси-β-D-рибоза
- 7. Входящие в состав нуклеиновых кислот гетероциклические основания пиримидинового и пуринового рядов называют нуклеиновыми основаниями.
- 8. Заместители в гетероциклическом ядре нуклеиновых оснований: оксогруппа аминогруппа одновременно обе эти группы
- 9. Нуклеиновые пиримидиновые основания Цитозин (Ц) Урацил (У) Тимин (Т)
- 10. Нуклеиновые пуриновые основания Аденин (А) Гуанин (Г)
- 11. Азотистое основание и углевод связаны между
- 12. N-гликозиды нуклеиновых оснований с рибозой или
- 13. Общая формула нуклеозидов
- 15. Номенклатура нуклеозидов Цитозин + рибоза →
- 16. аденозин дезоксиаденозин
- 17. цитидин дезоксицитидин
- 18. Нуклеозиды достаточно устойчивы к гидролизу в
- 19. Нуклеотиды - фосфаты нуклеозидов Реакция этерификации между
- 20. аденозин-5’-фосфат аденозин-3’-фосфат
- 21. Номенклатура нуклеотидов
- 22. Аденозин-5'-монофосфат (АМФ) Аденозин-5'-дифосфат (АДФ) Аденозин-5'-трифосфат (АТФ)
- 23. циклический 3',5'-АМФ (цАМФ) является естественно встречающимся
- 25. ТЦА
- 26. Первичная структура нуклеиновых кислот определяется последовательностью нуклеотидных
- 27. ДНК в основном содержится в ядрах
- 28. Вторичная структура ДНК Под вторичной структурой понимают пространственную организацию полинуклеотидной цепи.
- 29. Дж. Уотсон, Ф. Крик 1953 г. Вторичная
- 33. Основания, располагающиеся внутри спирали, прочно упакованы и
- 34. Т.е. первичная структура (нуклеотидная последовательность) одной цепи предопределяет первичную структуру второй цепи.
- 35. Правила Чаргаффа Количество пуриновых оснований равно количеству
- 36. Роль комплементарных взаимодействий в осуществлении биологической функции
- 37. Однако нуклеотидная последовательность ДНК под действием различных
- 38. Мутагены – вещества, вызывающие мутации: мутагены
- 39. HNO2 Будет связывать цитозин
- 40. Третичная структура ДНК У всех живых
- 41. репликация
- 42. Синтез мРНК на матрице ДНК
- 43. тРНК
- 44. Аминокислоты, необходимые для синтеза белка, доставляются
- 46. Уровни структурной организации белковых макромолекул
- 47. Последовательность расположения аминокислотных остатков в одной
- 48. Кроме первичной, в белковых молекулах выделяют вторичную,
- 49. Ключевую роль в стабилизации этой структуры играют
- 50. Вторичная структура белка (α-спираль)
- 51. В отличие от α–спирали β–структура образована за
- 52. Особенности вторичной структуры белка во многом определяются аминокислотным составом
- 53. Под третичной структурой белка (субъединицей) подразумевают пространственную
- 54. Типы взаимодействий, стабилизирующие третичную структуру белка
- 55. а - электростатическое взаимодействие б -
- 57. Под четвертичной структурой белка подразумевают ассоциированные между
- 59. Пространственная структура белковой молекулы способна нарушаться под
- 60. Биологические функции белков 1. Строительная (структурная).
- 61. 4. Транспортная. Белки крови транспортируют кислород, жирные
- 62. 7. Опорная. Белки входят в состав
- 63. Классификация белков 1. По форме молекул
- 64. Молекулы фибриллярных белков вытянуты в длину, нитеобразны
- 65. Фибриллярные белки служат основным строительным материалом. К
- 66. Глобулярные белки выполняют ряд функций, связанных с
- 67. 2. По степени сложности белки разделяют
- 68. К простым белкам относят: - альбумины –
- 69. - глютелины – содержатся в семенах злаков
- 70. К сложным белкам относят: - липопротеины =
- 71. - металлопротеины = белок + металл

Слайд 2
Полимерные цепи нуклеиновых кислот построены из мономерных единиц – нуклеотидов, в
связи с чем нуклеиновые кислоты называют полинуклеотидами.

Слайд 3
Мономерное звено представляет собой трёхкомпонентное образование, включающее:
гетероциклическое основание,
углеводный остаток,
фосфатную группу.




Слайд 7
Входящие в состав нуклеиновых кислот гетероциклические основания пиримидинового и пуринового рядов
называют нуклеиновыми основаниями.

Слайд 8Заместители в гетероциклическом ядре нуклеиновых оснований:
оксогруппа
аминогруппа
одновременно обе эти группы

 Урацил (У) Тимин (Т)")
 Гуанин (Г)")
Слайд 11
Азотистое основание и углевод связаны между собой N-гликозидной связью. При этом
N-гликозидная связь осуществляется между атомом углерода С-1 рибозы (дезоксирибозы) и атомом азота N-1 пиримидинового и N-9 пуринового оснований.

Слайд 12
N-гликозиды нуклеиновых оснований с рибозой или дезоксирибозой – нуклеозиды.
В зависимости от
природы углеводного остатка различают рибонуклеозиды и дезоксирибонуклеозиды.
В составе нуклеиновых кислот обнаруживаются только β-нуклеозиды.
В составе нуклеиновых кислот обнаруживаются только β-нуклеозиды.


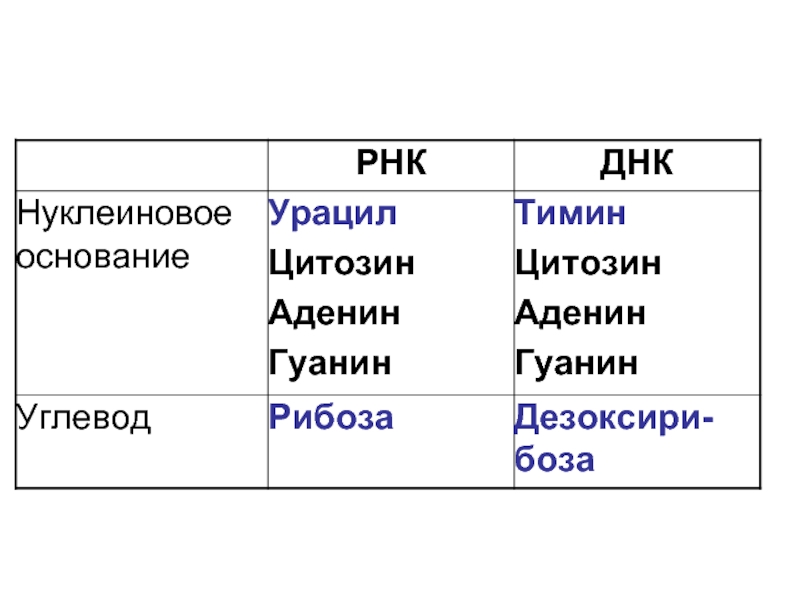
Слайд 15Номенклатура нуклеозидов
Цитозин + рибоза → цитидин
Цитозин + дезоксирибоза →
дезоксицитидин
Аденин + рибоза → аденозин
Аденин + дезоксирибоза → дезоксиаденозин
Аденин + рибоза → аденозин
Аденин + дезоксирибоза → дезоксиаденозин
-идин у пиримидиновых, -озин у пуриновых нуклеозидов



Слайд 18
Нуклеозиды достаточно устойчивы к гидролизу в слабощелочной среде. В кислой среде
они подвергаются гидролизу. При этом пуриновые нуклеозиды гидролизуются легче, чем пиримидиновые.

Слайд 19Нуклеотиды - фосфаты нуклеозидов
Реакция этерификации между фосфорной кислотой и нуклеозидом обычно
осуществляется при С-5 или
С-3 атоме в остатке рибозы (рибонуклеотиды) или дезоксирибозы (дезоксирибонуклеотиды).



Аденозин-5'-дифосфат (АДФ)Аденозин-5'-трифосфат (АТФ)")
Слайд 23
циклический 3',5'-АМФ (цАМФ) является естественно встречающимся рибонуклеотидом (он образуется из АТФ
в процессе реакции, катализируемой ферментом аденилатциклазой). цАМФ наделен рядом уникальных функций и высокой биологической активностью в регуляции процессов обмена, выполняя роль медиатора внеклеточных сигналов в клетках животных.
 является естественно встречающимся рибонуклеотидом (он образуется из АТФ в процессе реакции, катализируемой")
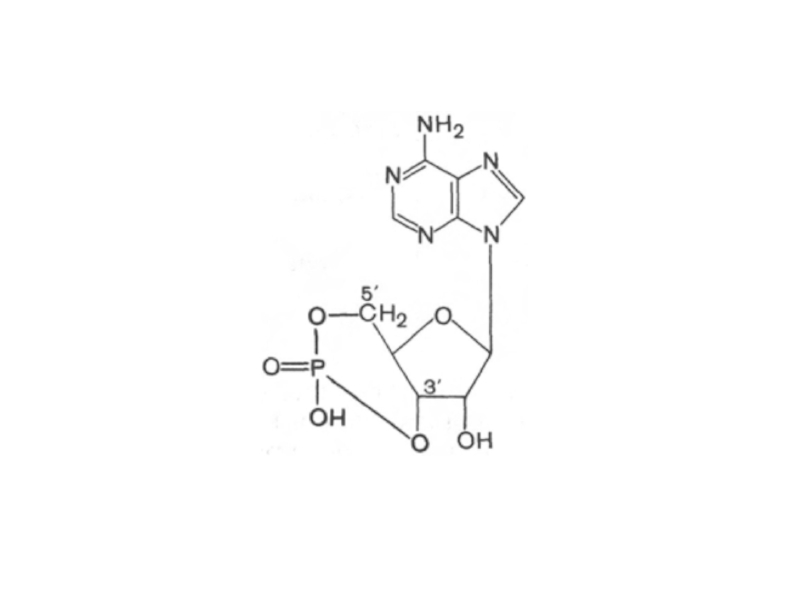

Слайд 26Первичная структура нуклеиновых кислот определяется последовательностью нуклеотидных звеньев, связанных ковалентными связями
в непрерывную цепь полинуклеотида.
Важной характеристикой нуклеиновых кислот служит нуклеотидный состав, т.е. набор и соотношение нуклеотидных компонентов.
Важной характеристикой нуклеиновых кислот служит нуклеотидный состав, т.е. набор и соотношение нуклеотидных компонентов.

Слайд 27
ДНК в основном содержится в ядрах клеток, а РНК находится в
рибосомах и в протоплазме клеток.
3 вида клеточных РНК (различаются по местоположению в клетке, составу и размерам, а также по функциям):
- транспортная (тРНК)
- матричная (мРНК)
- рибосомная (рРНК)
3 вида клеточных РНК (различаются по местоположению в клетке, составу и размерам, а также по функциям):
- транспортная (тРНК)
- матричная (мРНК)
- рибосомная (рРНК)

Слайд 28Вторичная структура ДНК
Под вторичной структурой понимают пространственную организацию полинуклеотидной цепи.

Слайд 29Дж. Уотсон, Ф. Крик
1953 г.
Вторичная структура ДНК
в виде двойной спирали
Молекула ДНК
состоит из двух
полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали, имеющей диаметр 1,8 – 2,0 нм.
Две нуклеотидные цепи антипараллельны друг другу (противоположные направления образования фосфодиэфирных связей 5’-3’ и 3’-5’). Пуриновые и пиримидиновые основания направлены внутрь спирали. Между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи возникают водородные связи. Эти основания составляют комплементарные пары.
полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали, имеющей диаметр 1,8 – 2,0 нм.
Две нуклеотидные цепи антипараллельны друг другу (противоположные направления образования фосфодиэфирных связей 5’-3’ и 3’-5’). Пуриновые и пиримидиновые основания направлены внутрь спирали. Между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи возникают водородные связи. Эти основания составляют комплементарные пары.

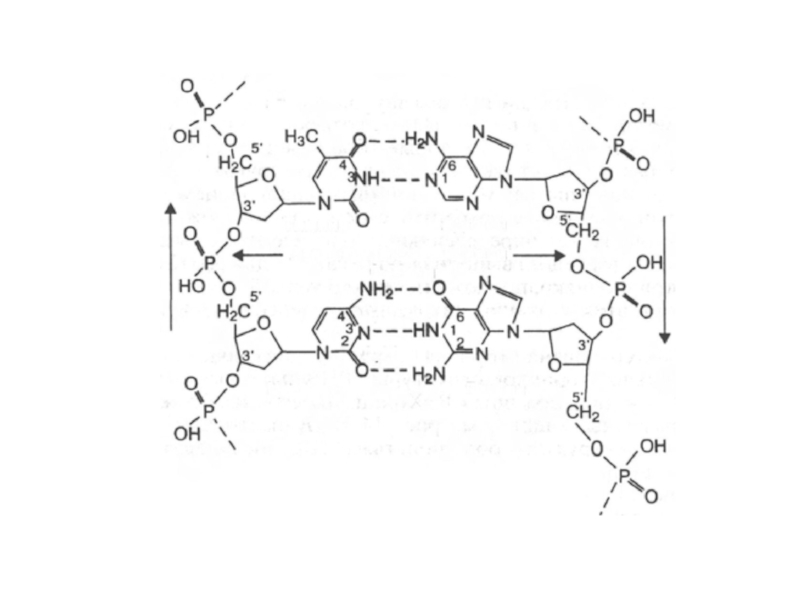
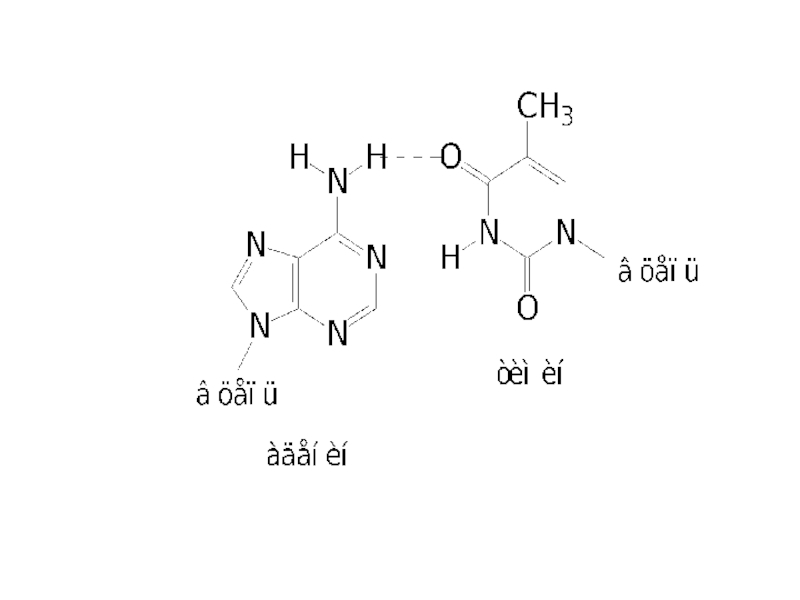
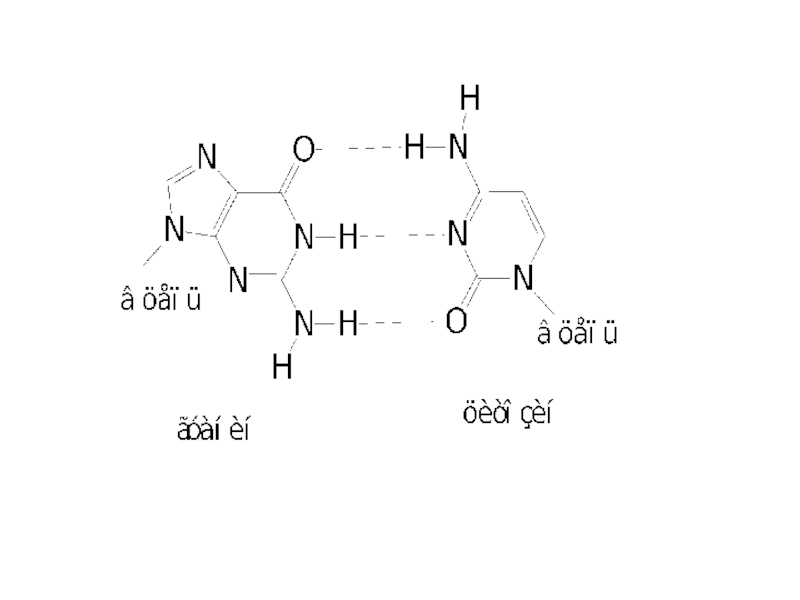
Слайд 33Основания, располагающиеся внутри спирали, прочно упакованы и не контактируют с водой.
Вода контактирует лишь с ОН – группами углевода и фосфатными группами.
Водородные связи между комплементарными основаниями – один из видов взаимодействий, стабилизирующих двойную спираль. Две цепи ДНК, образующие двойную спираль, не идентичны, но комплементарны между собой.
Водородные связи между комплементарными основаниями – один из видов взаимодействий, стабилизирующих двойную спираль. Две цепи ДНК, образующие двойную спираль, не идентичны, но комплементарны между собой.

Слайд 34Т.е. первичная структура (нуклеотидная последовательность) одной цепи предопределяет первичную структуру второй
цепи.
 одной цепи предопределяет первичную структуру второй цепи.")
Слайд 35Правила Чаргаффа
Количество пуриновых оснований равно количеству пиримидиновых оснований
Количество аденина равно количеству
тимина; количество гуанина равно количеству цитозина
Сумма аденина и цитозина равна сумме гуанина и тимина
Сумма аденина и цитозина равна сумме гуанина и тимина

Слайд 36Роль комплементарных взаимодействий в осуществлении биологической функции ДНК
Комплементарность цепей составляет химическую
основу важнейшей функции ДНК – хранения и передачи наследственных признаков.
Сохранность нуклеотидной последовательности – залог безошибочной передачи генетической информации.
Сохранность нуклеотидной последовательности – залог безошибочной передачи генетической информации.

Слайд 37Однако нуклеотидная последовательность ДНК под действием различных факторов может подвергаться изменениям
– мутациям.
Мутация – изменение наследственности.
Наиболее распространённый вид мутации – замена какой-либо пары оснований на другую. Одной из причин может быть сдвиг таутомерного равновесия. Другие причины – воздействие химических факторов или излучений.
Мутация – изменение наследственности.
Наиболее распространённый вид мутации – замена какой-либо пары оснований на другую. Одной из причин может быть сдвиг таутомерного равновесия. Другие причины – воздействие химических факторов или излучений.

Слайд 38
Мутагены – вещества, вызывающие мутации:
мутагены прямого действия,
промутагены, которые сами по себе
неактивны, но в организме под действием ферментов превращаются в мутагенные продукты.
Типичные мутагены – нитриты и азотистая кислота, которые могут образовываться в организме из нитратов.
Типичные мутагены – нитриты и азотистая кислота, которые могут образовываться в организме из нитратов.


Слайд 40Третичная структура ДНК
У всех живых организмов двухспиральные молекулы ДНК плотно
упакованы с образованием сложных трехмерных структур. Двухцепочечные ДНК прокариот и эукариот суперспирализированы. Суперспирализация необходима для компактной упаковки молекулы в небольшом объеме пространства, а также немаловажно для начала процессов репликации (“снятия копии”), а также для процесса биосинтеза белка (транскрипция). Третичная структура ДНК эукариот в отличие от прокариот функционирует только в комплексе с белками хромосом.




Слайд 44
Аминокислоты, необходимые для синтеза белка, доставляются в рибосомы с помощью т-РНК. Каждая молекула
т-РНК переносит только одну аминокислоту.
Антикодон — это последовательность из трех нуклеотидов, комплементарных нуклеотидам кодона в и-РНК.
Информация о первичной структуре молекулы белка зашифрована в молекуле ДНК.
Каждая аминокислота зашифрована последовательностью из трех нуклеотидов. Эта последовательность называется триплетом или кодоном (см. таблицы генетического кода).
Антикодон — это последовательность из трех нуклеотидов, комплементарных нуклеотидам кодона в и-РНК.
Информация о первичной структуре молекулы белка зашифрована в молекуле ДНК.
Каждая аминокислота зашифрована последовательностью из трех нуклеотидов. Эта последовательность называется триплетом или кодоном (см. таблицы генетического кода).



Слайд 47
Последовательность расположения аминокислотных остатков в одной или нескольких полипептидных цепях, составляющих
молекулу белка, – это первичная структура белка.

Слайд 48Кроме первичной, в белковых молекулах выделяют вторичную, третичную и четвертичную структуры.
Под
вторичной структурой белка подразумевают конформацию полипептидной цепи, т. е. способ её скручивания или складывания в соответствии с программой, заложенной в первичной структуре,
в α–спираль или β–структуру.

Слайд 49Ключевую роль в стабилизации этой структуры играют водородные связи, которые в
α–спирали образуются между карбонильным атомом кислорода каждого первого и атомом водорода NH–группы каждого пятого
α–аминокислотных остатков

")
Слайд 51В отличие от α–спирали β–структура образована за счёт межцепочечных водородных связей
между соседними участками полипептидной цепи


Слайд 53Под третичной структурой белка (субъединицей) подразумевают пространственную ориентацию или способ укладки
полипептидной цепи в определенном объеме, которая включает элементы вторичной структуры. Она стабилизируется за счет различных взаимодействий, в которых участвуют боковые радикалы α–аминокислотных остатков, находящихся в линейной полипептидной цепи на значительном удалении друг от друга, но сближенные в пространстве за счет изгибов цепи.
 подразумевают пространственную ориентацию или способ укладки полипептидной цепи в определенном")

Слайд 55
а - электростатическое взаимодействие
б - водородная связь
в - гидрофобные взаимодействия неполярных
групп
г - диполь-дипольные взаимодействия
д - дисульфидная (ковалентная)связь.
г - диполь-дипольные взаимодействия
д - дисульфидная (ковалентная)связь.

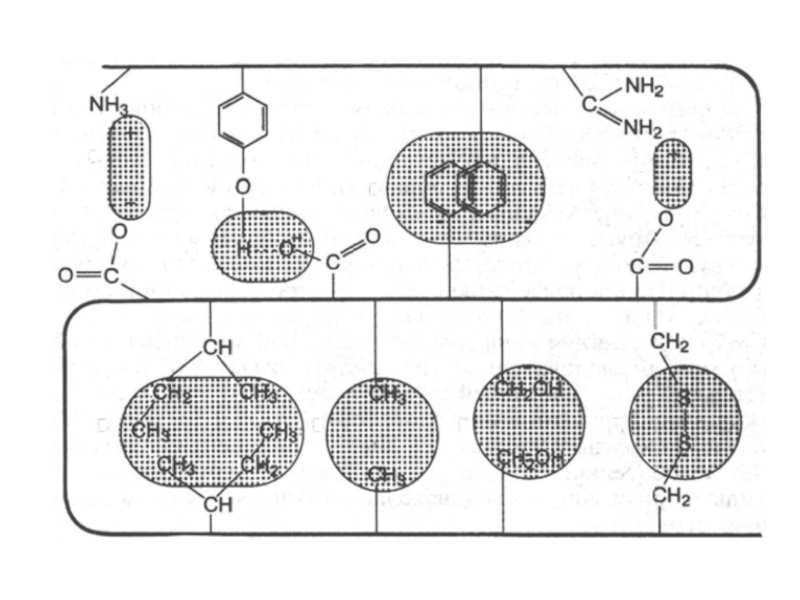
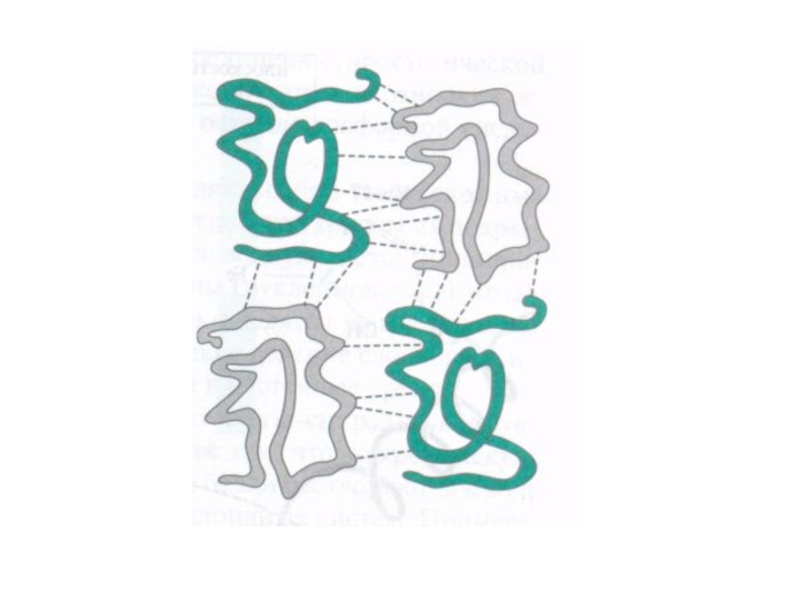
Слайд 57Под четвертичной структурой белка подразумевают ассоциированные между собой две или более
субъединиц, ориентированных в пространстве. Четвертичная структура поддерживается за счет водородных связей и гидрофобных взаимодействий. Она характерна для некоторых белков (гемоглобин).

Слайд 59Пространственная структура белковой молекулы способна нарушаться под влиянием изменения pH–среды, повышенной
температуры, облучения УФ–светом и т.д. Разрушение природной (нативной) макроструктуры белка называется денатурацией. В результате денатурации исчезает биологическая активность и снижается растворимость белков. Первичная структура белка при денатурации сохраняется.

Слайд 60Биологические функции белков
1. Строительная (структурная). Белки – основа протоплазмы любой
клетки, основной структурный материал всех клеточных мембран.
2. Каталитическая. Все ферменты являются белками.
3. Двигательная. Все формы движения в живой природе осуществляются белковыми структурами клеток.
2. Каталитическая. Все ферменты являются белками.
3. Двигательная. Все формы движения в живой природе осуществляются белковыми структурами клеток.
. Белки – основа протоплазмы любой клетки, основной структурный материал")
Слайд 614. Транспортная. Белки крови транспортируют кислород, жирные кислоты, липиды, гормоны. Специальные
белки переносят различные вещества через биомембраны.
5. Гормональная. Ряд гормонов относятся к белкам.
6. Запасная. Белки способны образовывать запасные отложения.
5. Гормональная. Ряд гормонов относятся к белкам.
6. Запасная. Белки способны образовывать запасные отложения.

Слайд 62
7. Опорная. Белки входят в состав костей скелета, сухожилий, суставов и
т. д.
8. Рецепторная. Рецепторные белки играют важную роль в передаче нервного или гормонального сигнала в клетку – мишень.
8. Рецепторная. Рецепторные белки играют важную роль в передаче нервного или гормонального сигнала в клетку – мишень.

Слайд 63Классификация белков
1. По форме молекул различают фибриллярные (волокнистые) и глобулярные
(корпускулярные) белки. Фибриллярные белки нерастворимы в воде. Глобулярные белки растворимы в воде или водных растворах кислот, оснований или солей. Из-за большого размера молекул образующиеся растворы являются коллоидными.
 и глобулярные (корпускулярные) белки. Фибриллярные белки")
Слайд 64Молекулы фибриллярных белков вытянуты в длину, нитеобразны и склонны группироваться одна
около другой с образованием волокон. В некоторых случаях они удерживаются рядом благодаря многочисленным водородным мостикам. Молекулы глобулярных белков сложены в компактные клубочки. Водородные связи в этом случае внутримолекулярные, и площадь соприкосновения между отдельными молекулами невелика. В этом случае межмолекулярные силы относительно слабы.

Слайд 65Фибриллярные белки служат основным строительным материалом. К их числу относят следующие
белки: кератин – в коже, волосах, ногтях, рогах и перьях; коллаген – в сухожилиях; миозин – в мускулах; фиброин – в шёлке.

Слайд 66Глобулярные белки выполняют ряд функций, связанных с поддержанием и регуляцией жизненных
процессов, - функций, требующих подвижности и, следовательно, растворимости. К их числу относят следующие белки: все ферменты, многие гормоны, например инсулин (из поджелудочной железы), тироглобулин (из щитовидной железы), адренокортикотропныйгормон (АКТГ) (из гипофиза); антитела, ответственные за аллергические реакции и обеспечивающие защиту от чужеродных организмов; альбумин яиц; гемоглобин, являющийся переносчиком кислорода из лёгких в ткани; фибриноген, который превращается в нерастворимый фибриллярный белок фибрин, что вызывает свёртывание крови.

Слайд 67
2. По степени сложности белки разделяют на простые и сложные. При
гидролизе простых белков получаются только аминокислоты. Сложные белки (протеиды) помимо собственно белковой части содержат небелковые остатки, называемые коферментами и простетическими группами.

Слайд 68К простым белкам относят:
- альбумины – водорастворимые белки, составляют 50% всех
белков плазмы крови человека, содержатся в белке яиц, молоке, растениях;
- глобулины – нерастворимые в воде белки, составляющие большую часть белков семян растений, особенно бобовых и масличных;
- проламины – характерны исключительно для семян злаков. Они играют роль запасных белков. В их составе много пролина и глутаминовой кислоты;
- глобулины – нерастворимые в воде белки, составляющие большую часть белков семян растений, особенно бобовых и масличных;
- проламины – характерны исключительно для семян злаков. Они играют роль запасных белков. В их составе много пролина и глутаминовой кислоты;

Слайд 69- глютелины – содержатся в семенах злаков и бобовых растений;
- гистоны
– присутствуют в ядрах клеток животных и растений, преобладают в белках хромосом;
- протамины – содержатся в половых клетках человека, животных и растений;
- протеиноиды – трудно растворимые белки с высоким содержанием серы – фибриллярные белки (фиброин – белок шёлка, кератины - белки волос, рогов, копыт, коллагены – белки соединительной ткани).
- протамины – содержатся в половых клетках человека, животных и растений;
- протеиноиды – трудно растворимые белки с высоким содержанием серы – фибриллярные белки (фиброин – белок шёлка, кератины - белки волос, рогов, копыт, коллагены – белки соединительной ткани).

Слайд 70К сложным белкам относят:
- липопротеины = белок + липид. Образуются за
счёт водородных связей и гидрофобного взаимодействия. Обязательные компоненты клеточных мембран, крови, мозга;
- фосфопротеины = белок + PO43- (остаток фосфорной кислоты связан с серином и треонином). Играют важную роль в питании молодых организмов (казеин молока, вителлин и фосвитин яичного желтка, ихтулин икры рыб);
- фосфопротеины = белок + PO43- (остаток фосфорной кислоты связан с серином и треонином). Играют важную роль в питании молодых организмов (казеин молока, вителлин и фосвитин яичного желтка, ихтулин икры рыб);

Слайд 71
- металлопротеины = белок + металл (Cu, Ca, Fe, Mn, Zn,
Ni, Mo, Se);
- гликопротеины = белок + углевод. К ним относятся фибриноген, протромбин (факторы свёртывания крови), гепарин (антисвёртывающее вещество), гормоны, интерферон (ингибитор размножения вирусов животных).
- гликопротеины = белок + углевод. К ним относятся фибриноген, протромбин (факторы свёртывания крови), гепарин (антисвёртывающее вещество), гормоны, интерферон (ингибитор размножения вирусов животных).
;- гликопротеины")





