- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Углеводы. Энергетический обмен презентация
Содержание
- 1. Углеводы. Энергетический обмен
- 2. Углеводы На долю углеводов приходится примерно 75%
- 3. 1. Строение углеводов Термин "углеводы", предложенный в
- 4. Классификация Углеводы можно разделить на 3 основные
- 5. Моносахариды Моносахариды - производные многоатомных спиртов, содержащие
- 6. Строение представителей моносахаридов
- 7. Пиранозы — моносахариды, находящиеся в циклической форме и содержащие
- 8. Строение представителей моносахаридов
- 9. Реакции моносахаридов (мутаротация, образование гликозидной связи, этерификация,
- 11. Реакции моносахаридов Образование гликозидов. Гликозидная связь имеет важное
- 12. Образование гликозидов. Все линейные олигомеры (кроме дисахаридов)
- 13. Образование гликозидной связи A. Образование α-1,4- и α-1,6-гликозидных
- 14. Образование О- и N-гликозидных связей в гликопротеинах.
- 15. Реакции моносахаридов Этерификация. Это реакция образования эфирной связи
- 16. Реакции моносахаридов Окисление и восстановление. При окислении концевых
- 17. Реакции моносахаридов
- 18. Олигосахариды Олигосахариды содержат несколько (от двух до
- 19. Дисахариды пищи Сахароза - дисахарид, состоящий из α-D-глюкозы
- 20. Полисахариды (классификация) Структурные различия между
- 21. В зависимости от выполняемых ими
- 22. Полисахариды пищи (крахмал, целлюлоза, гликоген) В пище
- 23. Крахмал Крахмал состоит из амилозы и амилопектина.
- 25. Полисахариды Целлюлоза (клетчатка) - основной структурный полисахарид растений.
- 26. Полисахариды Гликоген - полисахарид животных и человека. Так
- 27. Переваривание углеводов Эпителиальные клетки кишечника способны всасывать
- 28. Переваривание углеводов
- 29. Всасывание моносахаридов в кишечнике Моносахариды, образовавшиеся в
- 30. Всасывание моносахаридов в кишечнике
- 31. Влияние инсулина на перемещение транспортёров глюкозы из
- 32. Метаболизм углеводов После всасывания в кишечнике моносахариды
- 34. Фосфорилирование глюкозы В дальнейших превращениях в
- 35. Фосфорилирование глюкозы Глюкокиназа. Фосфорилирование глюкозы в гепатоцитах в
- 36. Метаболизм глюкозо-6-фосфата Глюкозо-6-фосфат может использоваться в клетке
- 37. Дефосфорилирование глюкозо-6-фосфата Превращение глюкозо-6-фосфата в глюкозу возможно
- 38. Катаболизм глюкозы
- 39. Аэробный гликолиз Аэробным гликолизом
- 40. Реакции аэробного распада глюкозы
- 41. Превращение глюкозо-6-фосфата в триозофосфаты (подготовительный этап)
- 42. 2. Эта часть аэробного гликолиза включает реакции,
- 43. Превращение глицеральдегид-3-фосфата в 3-фосфоглицерат.
- 44. В следующих реакциях происходят внутримолекулярные перестройки, смысл
- 45. Превращение 3-фосфоглицерата в пируват.
- 46. Баланс АТФ при аэробном гликолизе и распаде
- 47. Выход АТФ при аэробном распаде глюкозы до
- 50. Анаэробный распад глюкозы (анаэробный гликолиз) Анаэробным гликолизом
- 51. Характеристика реакции анаэробного гликолиза При анаэробном гликолизе
- 53. Баланс АТФ при анаэробном гликолизе Анаэробный гликолиз
- 54. Значение катаболизма глюкозы Основное физиологическое назначение катаболизма
- 55. Регуляция катаболизма глюкозы Поскольку основное значение гликолиза
- 56. Фосфофруктокиназа активируется АМФ, но ингибируется АТФ. АМФ,
- 58. Значение гликолиза в печени для синтеза жиров
- 59. Синтез глюкозы (глюконеогенез) Некоторые ткани, например мозг,
- 60. Первичными субстратами глюконеогенеза являются лактат,
- 61. Синтез глюкозы из лактата (цикл Кори) Лактат,
- 63. Синтез глюкозы из аминокислот Аминокислоты, которые при
- 65. Синтез глюкозы из глицерола Глицерол могут использовать
- 67. Пентозофосфатный путь превращения глюкозы Пентозофосфатный путь (фосфоглюконатный)
- 68. Общий путь катаболизма. Энергетический обмен Живые организмы
- 69. Общая схема обмена веществ и энергии. 1
- 70. БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ Процессы катаболизма в клетках животных
- 71. В живых организмах существует целая группа органических
- 72. Типы фосфорилирования
- 74. Углеводы, белки и жиры — основные компоненты
- 75. Основные этапы общего пути катаболизма В процессе
- 77. Окислительное декарбоксилирование пировиноградной кислоты
- 79. Цитратный цикл Цитратный цикл (цикл Кребса, цикл
- 82. Дыхательная цепь (ЦПЭ) Энергия освобождается в процессе
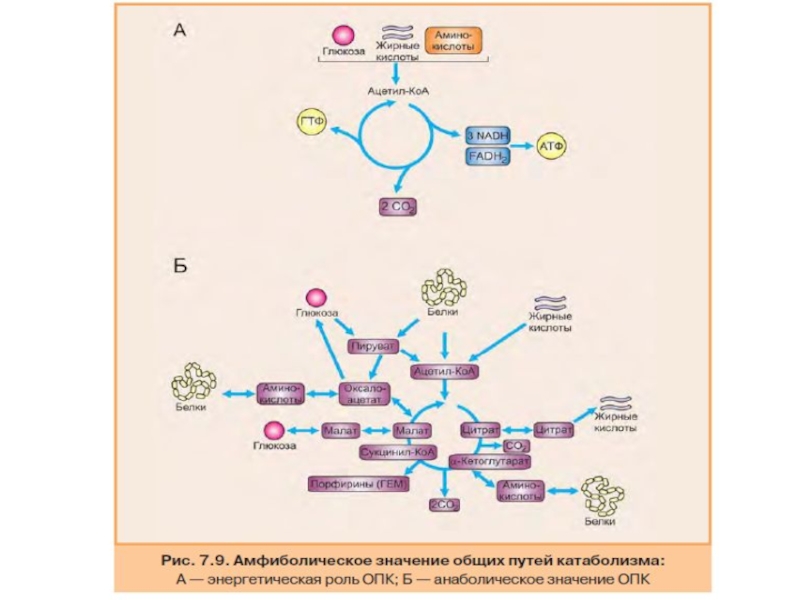
- 84. Амфиболическое значение общего пути катаболизма

Слайд 2Углеводы
На долю углеводов приходится примерно 75% массы пищевого суточного рациона и

Слайд 31. Строение углеводов
Термин "углеводы", предложенный в XIX столетии, был основан на

Слайд 4Классификация
Углеводы можно разделить на 3 основные группы в зависимости от количества
Моносахариды
Олигосахариды
Полисахариды.

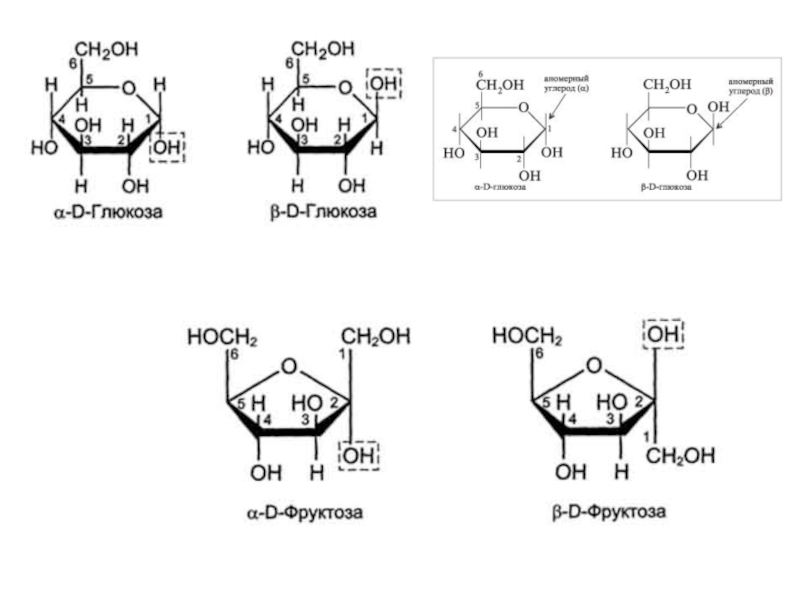
Слайд 5Моносахариды
Моносахариды - производные многоатомных спиртов, содержащие карбонильную группу. В зависимости от
Альдозы содержат функциональную альдегидную группу -НС=О, тогда как кетозы содержат кетонную группу >С=О. Название моносахарида зависит от числа составляющих его углеродных атомов, например альдотриозы, кетотриозы, альдогексозы, кетогексозы и т.д.
Моносахариды по строению можно отнести к простым углеводам, так как они не гидролизуются при переваривании, в отличие от сложных, которые при гидролизе распадаются с образованием простых углеводов
В пище человека (фрукты, мёд, соки) содержится небольшое количество моносахаридов, в основном глюкоза и фруктоза (знать циклические и линейные формулы фруктозы, глюкозы, галактозы!).


Слайд 7Пиранозы — моносахариды, находящиеся в циклической форме и содержащие шестичленное (пирановое) кольцо. Пиранозы
Фуранозы — моносахариды, находящиеся в циклической форме и содержащие пятичленное (фурановое) кольцо. Фуранозы образуются в результате внутримолекулярной ацетализации гидроксигруппы и карбонильной группы. Наиболее часто кольцо фуранозы является циклическим полуацеталем (альдопентозы) или циклическим полукеталем (кетогексозы). Фуранозы могут существовать в виде α- и β- аномеров.
 кольцо. Пиранозы образуются в результате внутримолекулярной")

Слайд 9Реакции моносахаридов (мутаротация, образование гликозидной связи, этерификация, окисление – восстановление)
Мутаротация, или
Мутаротация, или аномеризация -взаимопревращение аномерных форм")
Слайд 11Реакции моносахаридов
Образование гликозидов. Гликозидная связь имеет важное биологическое значение, потому что именно

Слайд 12Образование гликозидов.
Все линейные олигомеры (кроме дисахаридов) или полимеры содержат мономерные остатки,
Аномерная ОН-группа моносахарида может взаимодействовать с NН2-группой других соединений, что приводит к образованию N-гликозидной связи. Подобная связь присутствует в нуклеотидах и гликопротеинах
 или полимеры содержат мономерные остатки, участвующие в образовании двух")
Слайд 13Образование гликозидной связи
A. Образование α-1,4- и α-1,6-гликозидных связей. Б. Строение линейного полисахарида: 1 -

Слайд 14Образование О- и N-гликозидных связей в гликопротеинах.
1 - N-гликозидная связь между

Слайд 15Реакции моносахаридов
Этерификация. Это реакция образования эфирной связи между ОН-группами моносахаридов и различными

Слайд 16Реакции моносахаридов
Окисление и восстановление. При окислении концевых групп глюкозы -СНО и -СН2ОН


Слайд 18Олигосахариды
Олигосахариды содержат несколько (от двух до десяти) остатков моносахаридов, соединённых гликозидной
 остатков моносахаридов, соединённых гликозидной связью. Дисахариды - наиболее")
Слайд 19Дисахариды пищи
Сахароза - дисахарид, состоящий из α-D-глюкозы и β-D-фруктозы, соединённых α,β-1,2-гликозидной связью.
Лактоза - молочный сахар; важнейший дисахарид молока млекопитающих. В коровьем молоке содержится до 5% лактозы, в женском молоке - до 8%. В лактозе аномерная ОН-группа первого углеродного атома остатка D-галактозы связана β-гликозидной связью с четвёртым углеродным атомом D-глюкозы (β-1,4-связь). Поскольку аномерный атом углерода остатка глюкозы не участвует в образовании гликозидной связи, следовательно, лактоза относится к восстанавливающим сахарам.
Мальтоза поступает с продуктами, содержащими частично гидролизованный крахмал, например, солод, пиво. Мальтоза также образуется при расщеплении крахмала в кишечнике. Мальтоза состоит из двух остатков D-глюкозы, соединённых α-1,4-гликозидной связью.

Слайд 20Полисахариды (классификация)
Структурные различия между полисахаридами определяются:
строением моносахаридов, составляющих цепь.
типом гликозидных связей, соединяющих мономеры в цепи;
последовательностью остатков моносахаридов в цепи.
 Структурные различия между полисахаридами определяются:строением моносахаридов, составляющих цепь. В зависимости от строения остатков")
Слайд 21 В зависимости от выполняемых ими функций полисахариды можно разделить
резервные полисахариды, выполняющие энергетическую функцию. Эти полисахариды служат источником глюкозы, используемым организмом по мере необходимости. Резервная функция этих углеводов обеспечивается их полимерной природой. Полисахариды менее растворимы, чем моносахариды, следовательно они не влияют на осмотическое давление и поэтому могут накапливаться в клетке, например, крахмал - в клетках растений, гликоген - в клетках животных;
структурные полисахариды, обеспечивающие клеткам и органам механическую прочность;
полисахариды, входящие в состав межклеточного матрикса, принимают участие в образовании тканей, а также в пролиферации и дифференцировке клеток.

Слайд 22Полисахариды пищи (крахмал, целлюлоза, гликоген)
В пище человека в основном содержатся полисахариды
Крахмал - это резервный полисахарид растений, содержащийся в наибольшем количестве в зёрнах злаков (пшеница, кукуруза, рис и др.), а также луковицах, стеблях и клубнях растений (в картофеле примерно 65%). Крахмал - разветвлённый полисахарид, состоящий из остатков глюкозы (гомогликан). Он находится в клетках растений в виде гранул, практически нерастворим в воде.
В пище человека в основном содержатся полисахариды растительного происхождения - крахмал,")
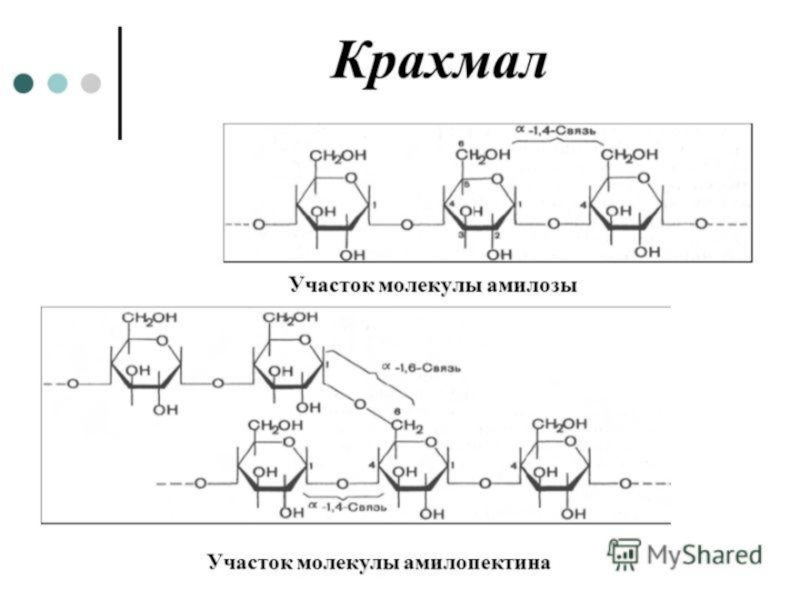
Слайд 23Крахмал
Крахмал состоит из амилозы и амилопектина. Амилоза - неразветвлённый полисахарид, включающий

Слайд 25Полисахариды
Целлюлоза (клетчатка) - основной структурный полисахарид растений. Это самое распространённое органическое соединение
Целлюлоза - линейный полисахарид гомогликан, построенный из остатков глюкозы, соединённых между собой β-1,4-гликозидными связями. Пищеварительная система человека не имеет ферментов, гидролизующих β-связи в полисахаридах. Поэтому целлюлоза - неиспользуемый углевод, но этот пищевой компонент необходим для нормального протекания переваривания.
 - основной структурный полисахарид растений. Это самое распространённое органическое соединение на земле. Доля целлюлозы")
Слайд 26Полисахариды
Гликоген - полисахарид животных и человека. Так же, как крахмал в растениях,
Гликоген представляет собой структурный аналог крахмала, но имеет большую степень ветвления: примерно на каждые 10 остатков глюкозы приходится одна α-1,6-гликозидная связь.

Слайд 27Переваривание углеводов
Эпителиальные клетки кишечника способны всасывать только моносахариды. Поэтому процесс переваривания


Слайд 29Всасывание моносахаридов в кишечнике
Моносахариды, образовавшиеся в результате переваривания, всасываются эпителиальными клетками
Всасывание моносахаридов из кишечника происходит путём облегчённой диффузии с помощью специальных белков-переносчиков (транспортёров). Кроме того, глюкоза и галактоза транспортируются в энтероцит путём вторично-активного транспорта, зависимого от градиента концентрации ионов натрия. Белки-транспортёры, зависимые от градиента Na+, обеспечивают всасывание глюкозы из просвета кишечника в энтероцит против градиента концентрации. Концентрация Na+, необходимая для этого транспорта, обеспечивается Nа+,К+-АТФ-азой, которая работает как насос, откачивая из клетки Na+ в обмен на К+. В отличие от глюкозы, фруктоза транспортируется системой, не зависйщей от градиента натрия.


Слайд 31Влияние инсулина на перемещение транспортёров глюкозы из цитоплазмы в плазматическую мембрану.
1

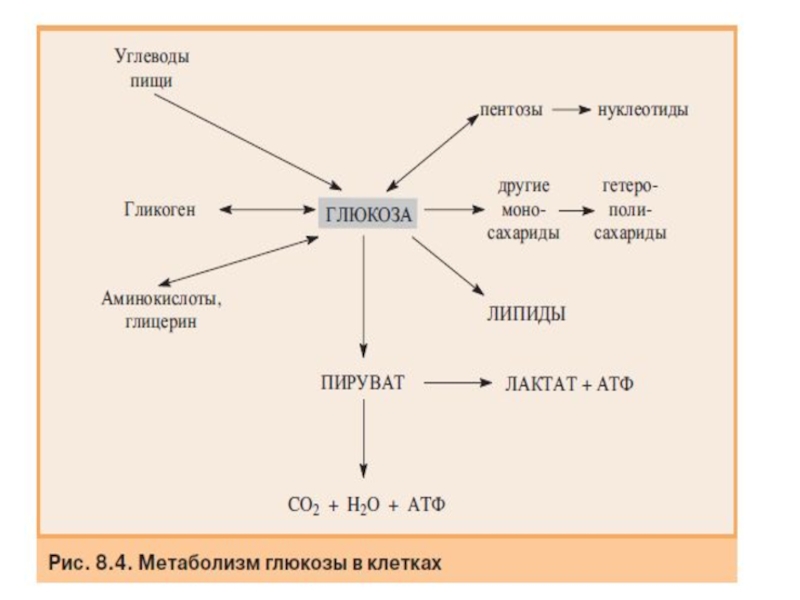
Слайд 32Метаболизм углеводов
После всасывания в кишечнике моносахариды поступают в воротную вену и

Слайд 34Фосфорилирование глюкозы
В дальнейших превращениях в клетках глюкоза и другие моносахариды
Глюкоза, поступающая в клетки органов и тканей, сразу же подвергается фосфорилированию с использованием АТФ. Эту реакцию во многих тканях катализирует фермент гексокиназа, а в печени и поджелудочной железе - фермент глюкокиназа. Фосфорилирование глюкозы - практически необратимая реакция, так как она протекает с использованием значительного количества энергии. Образование глюкозо-6-фосфата в клетке - своеобразная "ловушка" для глюкозы, так как мембрана клетки непроницаема для фосфорилированной глюкозы (нет соответствующих транспортных белков). Кроме того, Фосфорилирование уменьшает концентрацию свободной глюкозы в цитоплазме. В результате создаются благоприятные условия для облегчённой диффузии глюкозы в клетки из крови.

Слайд 35Фосфорилирование глюкозы
Глюкокиназа. Фосфорилирование глюкозы в гепатоцитах в период пищеварения обеспечивается свойствами глюкокиназы,
Гексокиназа отличается от глюкокиназы высоким сродством к глюкозе (Кm <0,1 ммоль/л). Следовательно, этот фермент, в отличие от глюкокиназы, активен при низкой концентрации глюкозы в крови, что характерно для постабсорбтивного состояния. Печень в этот период поглощает гораздо меньше глюкозы, так как скорость её внутриклеточного фосфорилирования глюкокиназой резко снижается. Тогда как потребление глюкозы мозгом, эритроцитами и другими тканями обеспечивается активной в этих условиях гексокиназой.

Слайд 36Метаболизм глюкозо-6-фосфата
Глюкозо-6-фосфат может использоваться в клетке в различных превращениях, основными из

Слайд 37Дефосфорилирование глюкозо-6-фосфата
Превращение глюкозо-6-фосфата в глюкозу возможно в печени, почках и клетках
Глюкозо-6-фосфат +Н2О → Глюкоза + Н3РО4
Образовавшаяся свободная глюкоза способна диффундировать из этих органов в кровь. В других органах и тканях глюкозо-6-фосфатазы нет, и поэтому дефосфорилирование глюкозо-6-фосфата невозможно. Пример подобного необратимого проникновения глюкозы в клетку - мышцы, где глюкозо-6-фосфат может использоваться только в метаболизме этой клетки.

Слайд 38Катаболизм глюкозы
Катаболизм глюкозы - основной поставщик
Основные пути катаболизма глюкозы
Окисление глюкозы до СО2 и Н2О (аэробный распад). Аэробный распад глюкозы можно выразить суммарным уравнением:
С6Н12О6 + 6 О2 → 6 СО2 + Н2О + 2820 кДж/моль.
Этот процесс включает несколько стадий .
1. Аэробный гликолиз - процесс окисления глюкозы с образованием двух молекул пирувата;
2. Общий путь катаболизма, включающий превращение пирувата в ацетил-КоА и его дальнейшее окисление в цитратом цикле;
3. ЦПЭ на кислород, сопряжённая с реакциями дегидрирования, происходящими в процессе распада глюкозы.
В определённых ситуациях обеспечение кислородом тканей может не соответствовать их потребностям. Например, на начальных стадиях интенсивной мышечной работы при стрессе сердечные сокращения могут не достигать нужной частоты, а потребности мышц в кислороде для аэробного распада глюкозы велики. В подобных случаях включается процесс, который протекает без кислорода и заканчивается образованием лактата из пировиноградной кислоты. Этот процесс называют анаэробным распадом, или анаэробным гликолизом. Анаэробный распад глюкозы энергетически малоэффективен, но именно этот процесс может стать единственным источником энергии для мышечной клетки в описанной ситуации. В даньнейшем, когда снабжение мышц кислородом будет достаточным в результате перехода сердца на ускоренный ритм, анаэробный распад переключается на аэробный. Пути катаболизма глюкозы и их энергетический эффект показаны на рис.

Слайд 39Аэробный гликолиз
Аэробным гликолизом называют процесс окисления глюкозы до
В аэробном гликолизе можно выделить 2 этапа.
Подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта серия реакций протекает с использованием 2 молекул АТФ.
Этап, сопряжённый с синтезом АТФ. В результате этой серии реакций фосфотриозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10 моль АТФ.

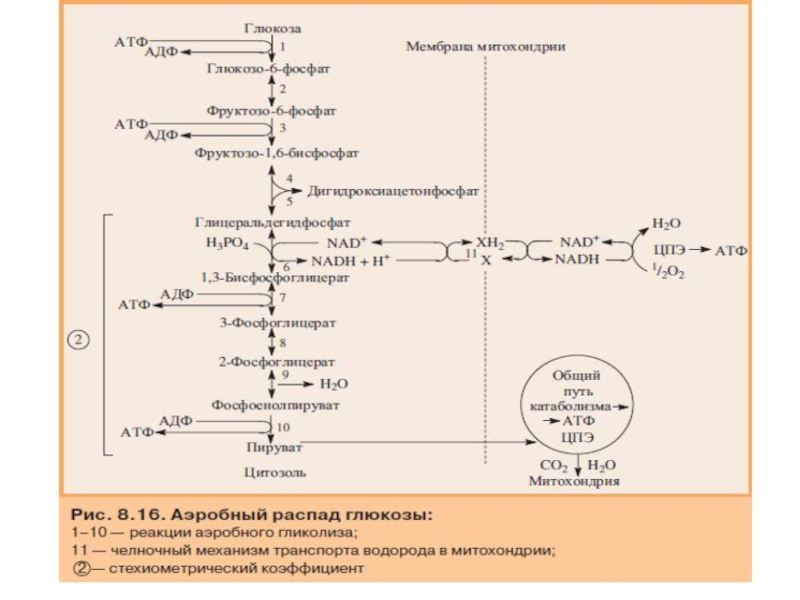
Слайд 40Реакции аэробного распада глюкозы
Гликолитическое расщепление глюкозы начинается с
1. Превращение глюкозо-6-фосфата в 2 молекулы глицеральдегид-3-фосфата (подготовительный этап)
Глюкозо-6-фосфат, образованный в результате фосфорилирования глюкозы с участием АТФ, в ходе следующей реакции превращается в фруктозо-6-фосфат. Эта обратимая реакция изомеризации протекает под действием фермента глюкозофосфатизомеразы.
Затем следует ещё одна реакция фосфорилирования с использованием фосфатного остатка и энергии АТФ. В ходе этой реакции, катализируемой фосфофруктокиназой, фруктозо-6-фосфат превращается в фруктозо-1,6-бисфосфат. Данная реакция, так же, как гексокиназная, практически необратима, и, кроме того, она наиболее медленная из всех реакций гликолиза. Реакция, катализируемая фосфофруктокиназой, определяет скорость всего гликолиза, поэтому, регулируя активность фосфофруктокиназы, можно изменять скорость катаболизма глюкозы.
Фруктозо-1,6-бисфосфат далее расщепляется на 2 триозофосфата: глицеральдегид-3-фосфат и дигидроксиацетонфосфат. Реакцию катализирует фермент фруктозобисфосфатальдолаза, или просто альдолаза. Этот фермент катализирует как реакцию альдольного расщепления, так и альдольной конденсации, т.е. обратимую реакцию. Продукты реакции альдольного расщепления - изомеры. В последующих реакциях гликолиза используется только глицеральдегид-3-фосфат, поэтому дигидроксиацетонфосфат превращается с участием фермента триозофосфатизомеразы в глицероальдегид-3-фосфат .
В описанной серии реакций дважды происходит фосфорилирование с использованием АТФ. Однако расходование двух молекул АТФ (на одну молекулу глюкозы) далее будет компенсировано синтезом большего количества АТФ.

")
Слайд 422. Эта часть аэробного гликолиза включает реакции, связанные с синтезом АТФ.
Образование АТФ описанным способом не связано с дыхательной цепью, и его называют субстратным фосфорилированием АДФ. Образованный 3-фосфоглицерат уже не содержит макроэргической связи.


Слайд 44В следующих реакциях происходят внутримолекулярные перестройки, смысл которых сводится к тому,
Превращение фосфоенолпирувата в пируват - необратимая реакция. Это вторая в ходе гликолиза реакция субстратного фосфорилирования. Образующаяся енольная форма пирувата затем неферментативно переходит в более термодинамически стабильную кетоформу.


Слайд 46Баланс АТФ при аэробном гликолизе
и распаде глюкозы до СО2 и Н2О
Выход АТФ при аэробном гликолизе:
На образование фруктозо-1,6-бисфосфата из одной молекулы глюкозы требуется 2 молекулы АТФ. Реакции, связанные с синтезом АТФ, происходят после распада глюкозы на 2 молекулы фосфотриозы, т.е. на втором этапе гликолиза. На этом этапе происходят 2 реакции субстратного фосфорилирования и синтезируются 2 молекулы АТФ (реакции 7 и 10). Кроме того, одна молекула глицеральдегид-3-фосфата дегидрируется (реакция 6), a NADH передаёт водород в митохондриальную ЦПЭ, где синтезируется 3 молекулы АТФ путём окислительного фосфорилирования. В данном случае количество АТФ (3 или 2) зависит от типа челночной системы. Следовательно, окисление до пирувата одной молекулы глицеральдегид-3-фосфата сопряжено с синтезом 5 молекул АТФ. Учитывая, что из глюкозы образуются 2 молекулы фосфотриозы, полученную величину нужно умножить на 2 и затем вычесть 2 молекулы АТФ, затраченные на первом этапе. Таким образом, выход АТФ при аэробном гликолизе составляет (5×2) - 2 = 8 АТФ.

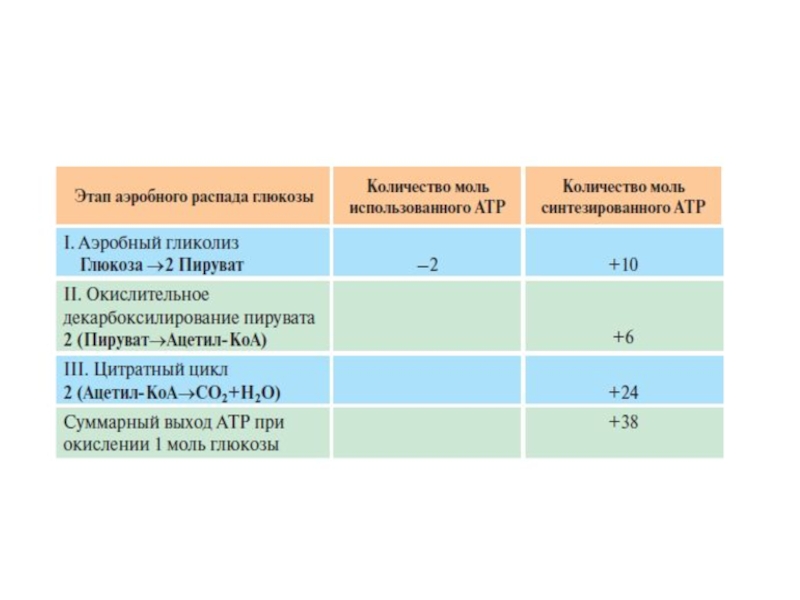
Слайд 47Выход АТФ при аэробном распаде глюкозы до конечных продуктов
В результате гликолиза
Таким образом, выход АТФ при окислении 1 моль глюкозы до СО2 и Н2О составляет 38 моль АТФ.
В процессе аэробного распада глюкозы происходят 6 реакций дегидрирования. Одна из них протекает в гликолизе и 5 в ОПК (см. раздел 6). Субстраты для специфических NAD-зависимых дегидрогеназ: глицеральдегид-3-фосфат, пируват, изоцитрат, α-кетоглутарат, малат. Одна реакция дегидрирования в цитратном цикле под действием сукцинатдегидрогеназы происходит с участием кофермента FAD. Общее количество АТФ, синтезированное путём окислительного фофорилирования, составляет 17 моль АТФ на 1 моль глицеральдегидфосфата. К этому необходимо прибавить 3 моль АТФ, синтезированных путём субстратного фосфорилирования (две реакции в гликолизе и одна в цитратном цикле).
Учитывая, что глюкоза распадается на 2 фос-фотриозы и что стехиометрический коэффициент дальнейших превращений равен 2, полученную величину надо умножить на 2, а из результата вычесть 2 моль АТФ, использованные на первом этапе гликолиза.

Слайд 50Анаэробный распад глюкозы (анаэробный гликолиз)
Анаэробным гликолизом называют процесс расщепления глюкозы с
Суммарное уравнение процесса:
С6Н12О6 + 2Н3РО4 + 2АDР = 2С3Н6О3 + 2АТФ + 2Н2О.
Анаэробным гликолизом называют процесс расщепления глюкозы с обра-зованием в качестве конечного")
Слайд 51Характеристика реакции анаэробного гликолиза
При анаэробном гликолизе в цитозоле протекают все 10

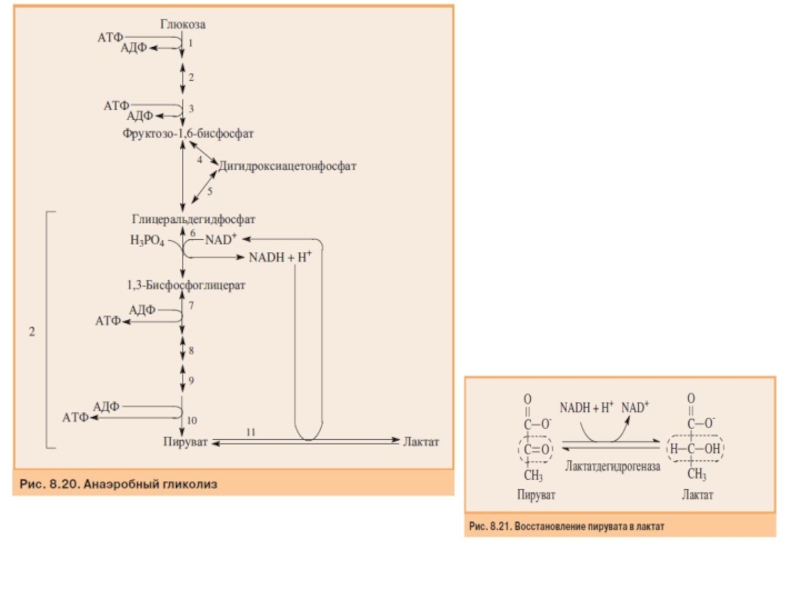
Слайд 53Баланс АТФ при анаэробном гликолизе
Анаэробный гликолиз по сравнению с аэробным менее
Итак, 10 цитозольных ферментов, катализирующих превра- щение глюкозы в пируват вместе с лактатдегидрогеназой обеспечивают в анаэробном гликолизе синтез 2 моль АТФ (на 1 моль глюкозы) без участия кислорода.

Слайд 54Значение катаболизма глюкозы
Основное физиологическое назначение катаболизма глюкозы заключается в использовании энергии,
Кроме энергетической функции процесс катаболизма глюкозы может выполнять и анаболические функции. Метаболиты гликолиза используются для синтеза новых соединений. Так, фруктозо-6-фосфат и глицеральдегид-3-фосфат участвуют в образовании рибозо-5-фосфата — структурного компонента нуклеотидов; 3-фосфоглицерат может включаться в синтез аминокислот, таких, как серин, глицин, цистеин. В печени и жировой ткани ацетил-КоА, образующийся из пирувата, используется как субстрат при биосинтезе жирных кислот, холестерина, а дигидроксиацетонфосфат как субстрат для синтеза глицерол-3-фосфата .

Слайд 55Регуляция катаболизма глюкозы
Поскольку основное значение гликолиза состоит в синтезе АТФ, его
Отношение уровня АТФ к АДФ и АМФ характеризует энергетический статус клетки, а его составляющие служат аллостерическими регуляторами скорости как общего пути катаболизма, так и гликолиза. Существенное значение для регуляции гликолиза имеет изменение актив- ности фосфофруктокиназы, потому что этот фермент, как упоминалось ранее, катализирует наиболее медленную реакцию процесса.

Слайд 56Фосфофруктокиназа активируется АМФ, но ингибируется АТФ. АМФ, связываясь с аллостерическим центром

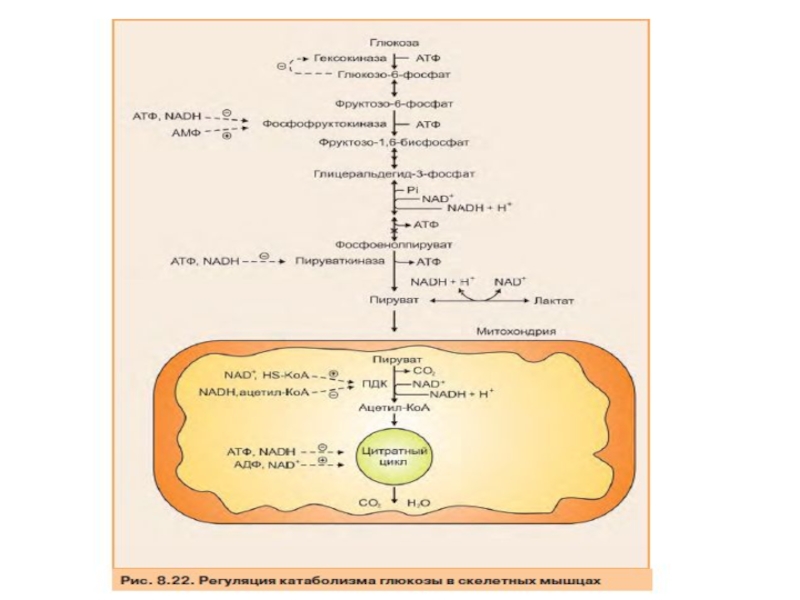
Слайд 58Значение гликолиза в печени для синтеза жиров
Основным значением ускорения гликолиза в

Слайд 59Синтез глюкозы (глюконеогенез)
Некоторые ткани, например мозг, нуждаются в постоянном поступлении глю-
Некоторые ткани, например мозг, нуждаются в постоянном поступлении глю- козы. Когда поступление углеводов")
Слайд 60 Первичными субстратами глюконеогенеза являются лактат, аминокислоты и глицерол. Включение
• Лактат является продуктом анаэробного гликолиза. Он образуется при любых состояниях организма (эритроциты, работающие мышцы). Таким образом, лактат используется в глюконеогенезе постоянно.
• Глицерол высвобождается при гидролизе жиров в жировой ткани в период голодания или при длительной физической нагрузке.
• Аминокислоты образуются в результате распада мышечных белков и включаются в глюконеогенез при длительном голодании или продолжительной мышечной работе.

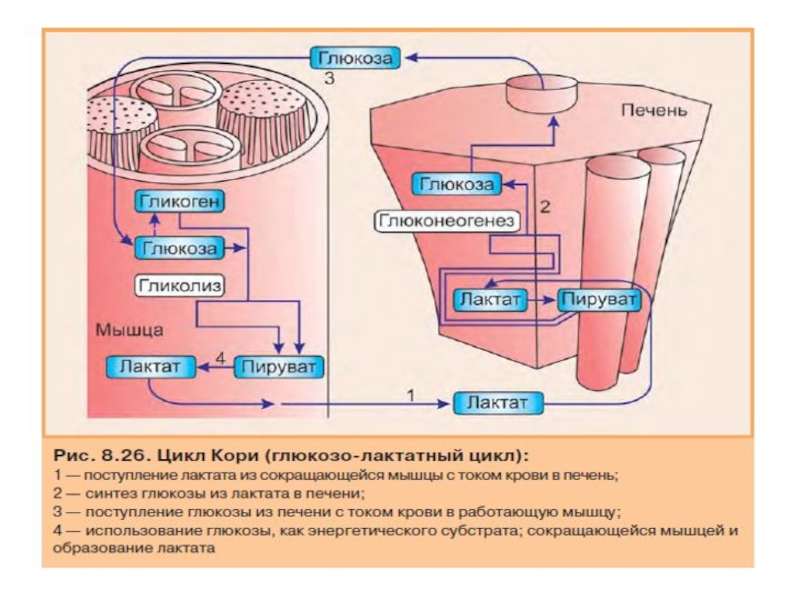
Слайд 61Синтез глюкозы из лактата
(цикл Кори)
Лактат, образовавшийся в интенсивно работающих мышцах или
Цикл Кори выполняет две важнейшие функции:
1 — обеспечивает утилизацию лактата, часть лактата используется для синтеза глюкозы;
2 — предотвращает накопление лактата и как следствие этого опасное снижение рН (лактоацидоз). Другая часть лактата окисляется печенью до СО2 и Н2О. Энергия окисления может использоваться для синтеза АТФ, необходимого для реакций глюконеогенеза.
Лактат, образовавшийся в интенсивно работающих мышцах или в клетках с")
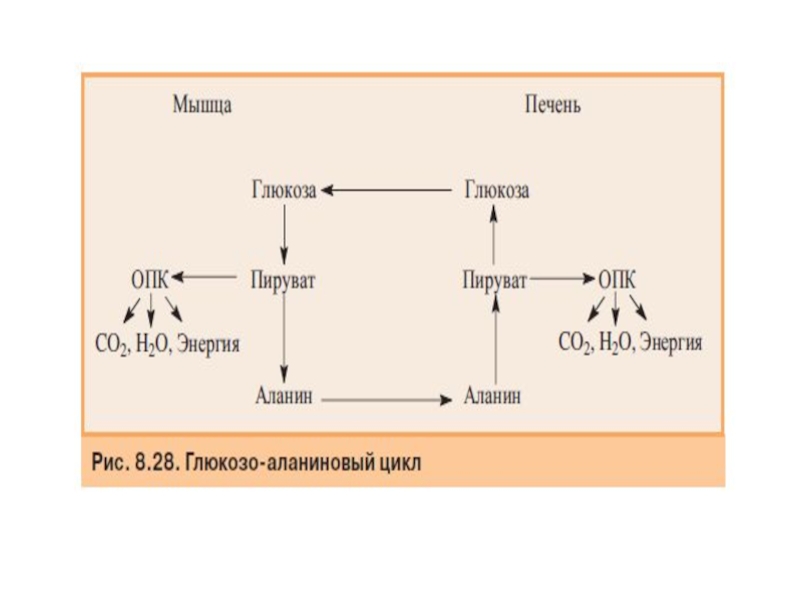
Слайд 63Синтез глюкозы из аминокислот
Аминокислоты, которые при катаболизме превращаются в пируват или

Слайд 65Синтез глюкозы из глицерола
Глицерол могут использовать только те ткани, в которых

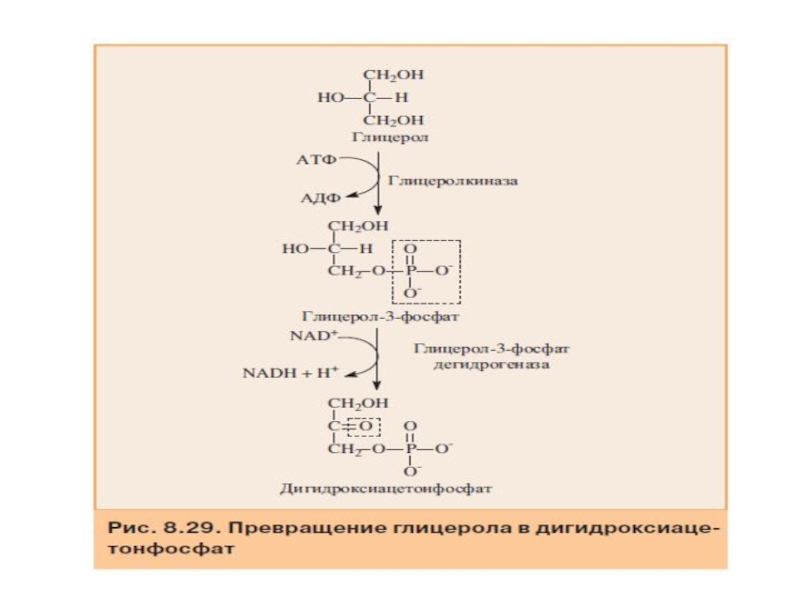
Слайд 67Пентозофосфатный путь превращения глюкозы
Пентозофосфатный путь (фосфоглюконатный) — альтернативный путь окисления глюкозо-6-фосфата.
 — альтернативный путь окисления глюкозо-6-фосфата. Пентозофосфатный путь состоит из")
Слайд 68Общий путь катаболизма.
Энергетический обмен
Живые организмы находятся в постоянной и неразрывной связи
Поступление веществ в организм происходит в результате дыхания (кислород) и питания. В ЖКТ продукты питания перевариваются (расщепляются до простых веществ). При переваривании происходит гидролиз полимеров (белков, полисахаридов и других сложных органических веществ) до мономеров, всасывающихся в кровь и включающихся в промежуточный обмен.
Промежуточный обмен (внутриклеточный метаболизм) включает 2 типа реакций: катаболизм и анаболизм (рис. 6-1).
Катаболизм - процесс расщепления органических молекул до конечных продуктов. Конечные продукты превращений органических веществ у животных и человека - СО2, Н2О и мочевина. В процессы катаболизма включаются метаболиты, образующиеся как при пищеварении, так и при распаде структурно-функциональных компонентов клеток.
Реакции катаболизма сопровождаются выделением энергии (экзергонические реакции).
Анаболизм объединяет биосинтетические процессы, в которых простые строительные блоки соединяются в сложные макромолекулы, необходимые для организма. В анаболических реакциях используется энергия, освобождающаяся при катаболизме (эндергонические реакции).

Слайд 69Общая схема обмена веществ и энергии.
1 - пищеварение; 2 - катаболизм;

Слайд 70БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ
Процессы катаболизма в клетках животных сопровождаются потреблением кислорода, который необходим

Слайд 71В живых организмах существует целая группа органических фосфатов, гидролиз которых приводит
АТФ - молекула, богатая энергией, поскольку она содержит две фосфоаншдридные связи (β, γ). При гидролизе концевой фосфоангидридной связи АТФ превращается в АДФ и ортофосфат Рi При этом изменение свободной энергии составляет -7,3 ккал/моль.



Слайд 74Углеводы, белки и жиры — основные компоненты пищи человека, являются многокомпонентными

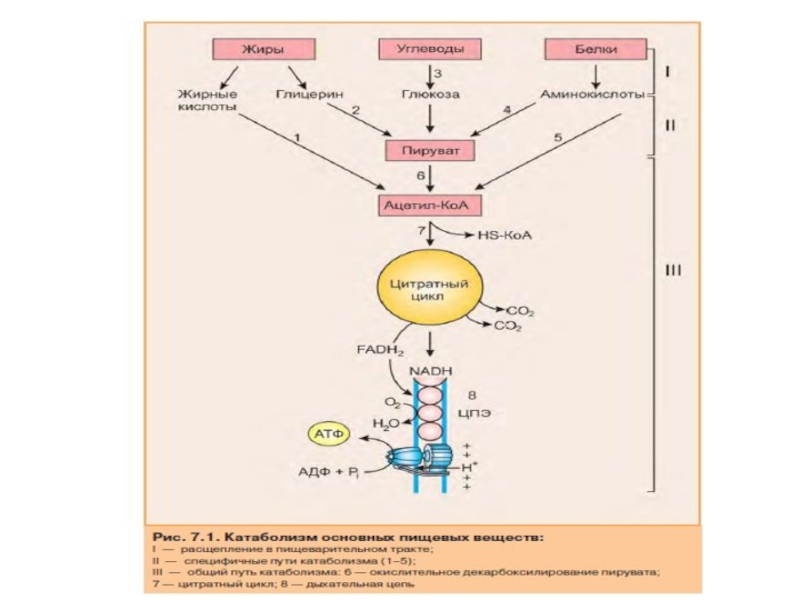
Слайд 75Основные этапы общего пути катаболизма
В процессе катаболизма можно выделить три основные
1. Расщепление в пищеварительном тракте. Это гидролитические реакции, превращающие сложные пищевые вещества в относительно небольшое число простых метаболитов: глюкозу, аминокислоты, глицерол, жирные кислоты.
2. Специфические пути катаболизма. На этом этапе простые метаболиты подвергаются специфическим реакциям расщепления, в результате которых образуется либо пировиноградная кислота, либо ацетил-КоА. Ацетил-КоА может образоваться из пирувата, а также из жирных кислот и аминокислот. В специфических путях катаболизма могут образоваться со единения, которые непосредственно включаются в цитратный цикл.
3. Окислительное декарбоксилирование пирувата, цитратный цикл и дыхательная цепь завершают расщепление пищевых веществ до конечных продуктов — СО и Н2О.
Следовательно, начиная со стадии образования пирувата, происходит унификация путей катаболизма. Из большого числа исходных соединений образуется всего два — пируват и ацетил-КоА. Процесс, начинающийся с окисления пирувата, называется общим путем катаболизма. Именно в общем пути катаболизма образуется основное количество субстратов для реакций дегидрирования. Совместно с дыхательной цепью и окислительным фосфорилированием общий путь катаболизма является основным источником энергии в форме АТФ.



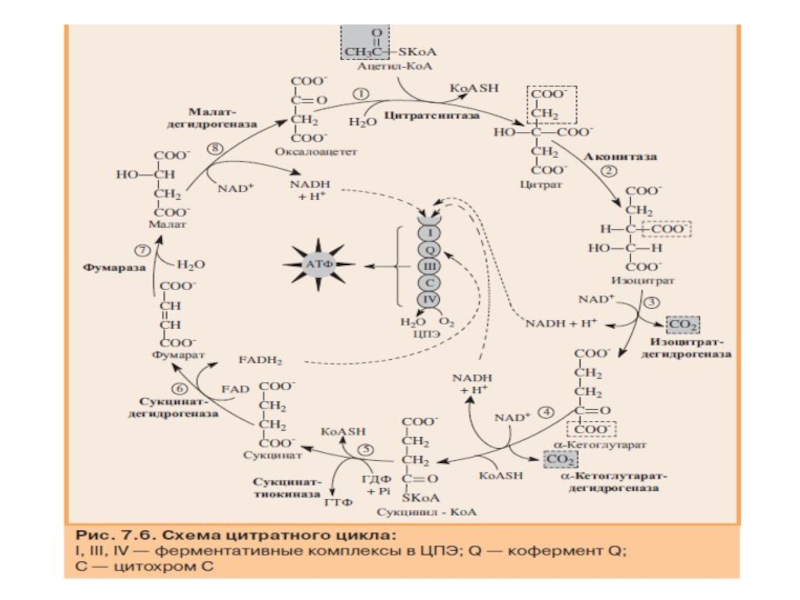
Слайд 79Цитратный цикл
Цитратный цикл (цикл Кребса, цикл трикарбоновых кислот) — это система
полное окисление ацетильного ос татка до двух молекул СО2;
образование трех молекул восстановленного NADH + H+ и одной молекулы FADH2;
синтез одной молекулы ГТФ в результате субстратного фосфори лирования.
 — это система реакций, приводящая к полному")

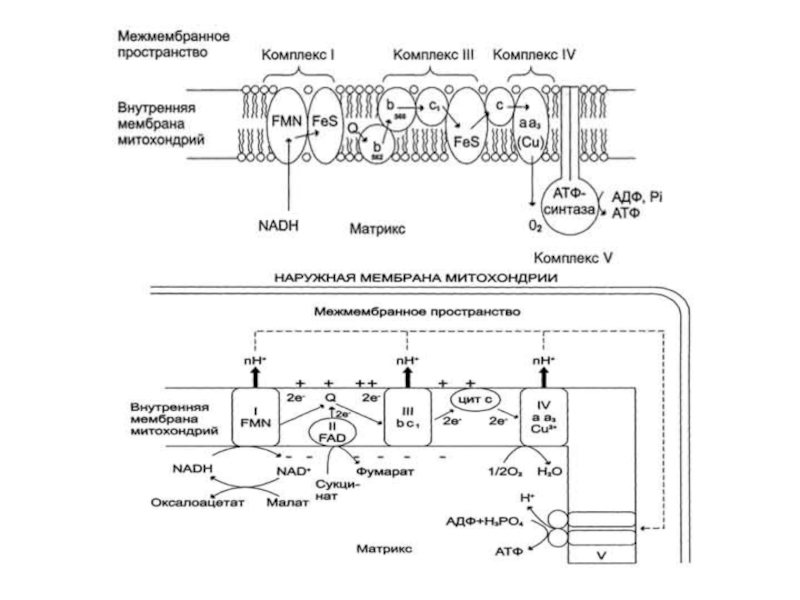
Слайд 82Дыхательная цепь (ЦПЭ)
Энергия освобождается в процессе ферментативного окисления метаболитов специфическими дегидрогеназами.
Окисление органических веществ в клетках, сопровождающееся потреблением кислорода и синтезом воды, называют тканевым дыханием, а цепь переноса электронов (ЦПЭ) - дыхательной цепью.
Электроны, поступающие в ЦПЭ, по мере их продвижения от одного переносчика к другому теряют свободную энергию. Значительная часть этой энергии запасается в форме АТФ, а часть энергии рассеивается в вице тепла. Кроме того, электроны с высоким энергетическим потенциалом, возникающие при окислении различных субстратов, могут быть использованы в реакциях биосинтеза, для которых помимо АТФ требуются восстановительные эквиваленты, например NADPH.
Перенос электронов от NADH к О2 включает ряд переносчиков, которые локализованы во внутренней мембране митохондрий. За исключением убихинона и цитохрома С, это сложные белковые комплексы.
Энергия освобождается в процессе ферментативного окисления метаболитов специфическими дегидрогеназами. В реакциях дегидрирования электроны")