случаев определяется энергией геотермальных источников.
Тепловой эффект солнечного излучения зависит от ИК-лучей.
Тепло распределяется по Земле в зависимости от высоты стояния Солнца над горизонтом и угла падения солнечных лучей (поэтому тепловой режим неодинаков на разных широтах и на разной высоте над уровнем моря).
Температурный фактор характеризуется ярко выраженными сезонными и суточными колебаниями.
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Температура как экологический фактор презентация
Содержание
- 1. Температура как экологический фактор
- 2. Распределение энергии Солнца на Земле
- 3. Распределение солнечной радиации на поверхности Земли (кВт· ч/м2/год)
- 4. Изменение температуры по мере подъема в
- 5. Температура влияет на организмы через ускорение
- 6. Температура как экологический фактор Все организмы (и
- 7. По типу терморегуляции организмы
- 9. Эктотермные организмы
- 10. Эндотермия
- 11. Концепция градусо-дней Температура влияет на скорость роста
- 12. «Эффективная температура» - разность между реально наблюдаемой
- 13. Примеры: Для икры форели (вид обитает
- 14. Почему в тундре не растут деревья? Распространение
- 15. Растения и температура По степени адаптации растений
- 16. Растения и температура 3) льдоустойчивые, или морозоустойчивые, растения– произрастают в областях
- 17. Растения и температура По степени адаптации к
- 18. Действие высоких температур Для большинства организмов высокие
- 19. Действие высоких температур Обитатели жарких пустынь Африки
- 20. Действие низких температур При температурах ниже -1оС
- 21. Температура для эндотермных животных Эндотермные организмы в
- 22. Температура для эндотермных животных Основные отличия гомойотермных
- 23. Эндотермные животные всегда вырабатывают определенный минимум
- 24. Потребление кислорода млекопитающими разной величины (поSchmidt–Nielsen, 1975).
- 25. Изменение температуры тела и теплопродукции
- 26. В пределах некоторого диапазона внешних температур
- 27. Физическая терморегуляция экологически выгодна, так как адаптация
- 28. Противоточная система сохранения тепла внутренних тканей в
- 29. Гетеротермия у эндотермных животных. Температура конечностей арктических
- 30. Тепловая одышка млекопитающих Вверху. Поток воздуха
- 31. Теплокровным свойственна химическая терморегуляция – рефлекторное увеличение теплопродукции
- 32. Поддержание температуры за счет возрастания теплопродукции
- 33. Температура для эндотермных животных Для защиты от
- 34. правило Бергмана Чем крупнее животное и компактнее
- 35. Правило Бергмана Если существует род, виды которого
- 36. Температура для эндотермных животных Правило Аллена
- 37. Правило Аллена Животные, обитающие в областях с
- 38. Температура для эндотермных животных Из правил Аллена
- 39. Адаптации к температурному фактору отражены в правиле
- 40. Поведенческая терморегуляция Особенности строения нор и
- 41. Температура как экологический фактор Эндотермы придерживаются стратегии,
Слайд 1
Температура как экологический фактор
Температура местообитания связана с солнечным излучением, в ряде


")
Слайд 4
Изменение температуры по мере подъема в воздушной среде, а также погружения
в водную или почвенную среду называют температурной стратификацией. Обычно наблюдается непрерывное снижение температуры с определенным градиентом.
В летний период в атмосфере возможна температурная инверсия - охлажденные слои воздуха смещаются вниз и располагаются под теплыми слоями. Вследствие этого происходит накопление загрязняющих веществ в приземном слое воздуха
В летний период в атмосфере возможна температурная инверсия - охлажденные слои воздуха смещаются вниз и располагаются под теплыми слоями. Вследствие этого происходит накопление загрязняющих веществ в приземном слое воздуха

Слайд 5
Температура влияет на организмы через ускорение или торможение биохимических реакций. Скорость
реакции с увеличением температуры на каждые 10оС увеличивается в 1,5-2 раза, т.е. и биохимические реакции подчиняются правилу Ван-Гоффа.
Диапазон оптимальных температур для живых организмов – от 0оС до +45оС.
Температуры ниже ОоС опасны образованием льда в клетках, который механически режет клеточные структуры; а кристаллы льда начинают отнимать воду из органоидов и из цитоплазмы, приводя к обезвоживанию клетки.
Температуры выше +45о опасны денатурацией ферментов.
Однако живые организмы могут переносить температуры ниже и выше оптимальных значений, благодаря анатомо-морфологическим, физиологическим и поведенческим адаптациям.
Диапазон оптимальных температур для живых организмов – от 0оС до +45оС.
Температуры ниже ОоС опасны образованием льда в клетках, который механически режет клеточные структуры; а кристаллы льда начинают отнимать воду из органоидов и из цитоплазмы, приводя к обезвоживанию клетки.
Температуры выше +45о опасны денатурацией ферментов.
Однако живые организмы могут переносить температуры ниже и выше оптимальных значений, благодаря анатомо-морфологическим, физиологическим и поведенческим адаптациям.

Слайд 6Температура как экологический фактор
Все организмы (и растения и животных) по способности
регулировать температуру тела принято делить на две группы:
Эктотермные - получают тепло от внешних источников, сами тепло не вырабатывают, это растения и холоднокровные животные.
Эндотермные - способны сами регулировать температуру тела за счет внутренней теплопродукции, это птицы и млекопитающие.
Эктотермные - получают тепло от внешних источников, сами тепло не вырабатывают, это растения и холоднокровные животные.
Эндотермные - способны сами регулировать температуру тела за счет внутренней теплопродукции, это птицы и млекопитающие.
 по способности регулировать температуру тела принято")


Слайд 10
Эндотермия у пойкилотермного позвоночного.
А. Самка питона высиживает яйца, генерируя тепло
за счет изометрического сокращения мышц, которое мало полезно для движения, но обеспечивает теплообразование.
Б. По мере снижения температуры окружающей среды частота сокращений повышается, С ростом частоты сокращений температура тела питона соответственно увеличивается. (Hutchinson et al., 1966.)
Б. По мере снижения температуры окружающей среды частота сокращений повышается, С ростом частоты сокращений температура тела питона соответственно увеличивается. (Hutchinson et al., 1966.)

Слайд 11Концепция градусо-дней
Температура влияет на скорость роста и развития эктотермов,
При температурах,
превышающих порог развития, скорость роста и развития возрастает линейно с повышением температуры.
Для завершения развития яиц кузнечика необходима То= +16о С ( порог развития, ниже этой То развитие не идет). При +20оС (на 4о выше порога развития) для развития личинок необходимо 17,5 суток, а при +30оС (на 14о выше порога) развитие личинок происходит за 5 суток.
(30о-16о) х 5 = 70 град/сут. (20о-16о) х 17,5 = 70 град/сут.
Для завершения развития кузнечиков необходима некая комбинация времени и температуры – сумма эффективных температур. Время для эктотермных организмов зависит от температуры, и, если она упадет ниже порога развития, то время для них поистине остановится.
Для завершения развития яиц кузнечика необходима То= +16о С ( порог развития, ниже этой То развитие не идет). При +20оС (на 4о выше порога развития) для развития личинок необходимо 17,5 суток, а при +30оС (на 14о выше порога) развитие личинок происходит за 5 суток.
(30о-16о) х 5 = 70 град/сут. (20о-16о) х 17,5 = 70 град/сут.
Для завершения развития кузнечиков необходима некая комбинация времени и температуры – сумма эффективных температур. Время для эктотермных организмов зависит от температуры, и, если она упадет ниже порога развития, то время для них поистине остановится.

Слайд 12«Эффективная температура» - разность между реально наблюдаемой температурой - t и
условным (биологическим) нулем t0
Сумма температур – это как правило «сумма эффективных температур», т.е. величин (t – t0)
 нулем t0")
Слайд 13Примеры:
Для икры форели (вид обитает в высоких широтах) «биологический нуль» приходится
примерно на 0о. Для полного развития икры соответственно требуется:
при температуре 2оС - 205 суток,
при 5оС - 82 суток,
при 10оС - 41 сутки.
Сумма температур во всех случаях оказывается равной 410 градусо-дней.
при температуре 2оС - 205 суток,
при 5оС - 82 суток,
при 10оС - 41 сутки.
Сумма температур во всех случаях оказывается равной 410 градусо-дней.
 «биологический нуль» приходится примерно на 0о.")
Слайд 14Почему в тундре не растут деревья? Распространение леса на севере Евразии ограничено
суммой эффективных температур
(превышающих t0 = + 10o)
700-800 градусо-дней,
но на Таймыре –
500-600 градусо-дней

Слайд 15Растения и температура
По степени адаптации растений к условиям крайнего дефицита тепла
можно выделить три группы:
1) нехолодостойкие растения– сильно повреждаются или гибнут при температурах, еще не достигающих точки замерзания воды. Гибель связана с инактивацией ферментов, нарушением обмена нуклеиновых кислот и белков, проницаемости мембран и прекращением тока ассимилятов. Это растения дождевых тропических лесов, водоросли теплых морей;
2) неморозостойкие растения– переносят низкие температуры, но гибнут, как только в тканях начинает образовываться лед. При наступлении холодного времени года у них повышается концентрация осмотически активных веществ в клеточном соке и цитоплазме, что понижает точку замерзания до – (5–7) °С. Вода в клетках может охлаждаться ниже точки замерзания без немедленного образования льда. некоторые вечнозеленые субтропические растения – лавры, лимоны и др.
2) неморозостойкие растения– переносят низкие температуры, но гибнут, как только в тканях начинает образовываться лед. При наступлении холодного времени года у них повышается концентрация осмотически активных веществ в клеточном соке и цитоплазме, что понижает точку замерзания до – (5–7) °С. Вода в клетках может охлаждаться ниже точки замерзания без немедленного образования льда. некоторые вечнозеленые субтропические растения – лавры, лимоны и др.

Слайд 16Растения и температура
3) льдоустойчивые, или морозоустойчивые, растения– произрастают в областях с сезонным климатом, с холодными
зимами. Во время сильных морозов надземные органы деревьев и кустарников промерзают, но тем не менее сохраняют жизнеспособность, так как в клетках кристаллического льда не образуется. Растения подготавливаются к перенесению морозов постепенно, проходя предварительную закалку после того, как заканчиваются ростовые процессы. Закалка заключается в накоплении в клетках сахаров (до 20–30 %), производных углеводов, некоторых аминокислот и других защитных веществ, связывающих воду. При этом морозоустойчивость клеток повышается, так как связанная вода труднее оттягивается образующимися во внеклеточных пространствах кристаллами льда.
 льдоустойчивые, или морозоустойчивые, растения– произрастают в областях с сезонным климатом, с холодными зимами. Во время сильных")
Слайд 17Растения и температура
По степени адаптации к высоким температурам можно выделить следующие
группы растений:
1) нежаростойкие растения повреждаются уже при +(30–40) °С (эукариотические водоросли, водные цветковые, наземные мезофиты);
2) жаровыносливые растения переносят получасовое нагревание до +(50–60) °С (растения сухих местообитаний с сильной инсоляцией – степей, пустынь, саванн, сухих субтропиков и т. п.).
Некоторые растения регулярно испытывают влияние пожаров, когда температура кратковременно повышается до сотен градусов. Пожары особенно часты в саваннах, в сухих жестколистных лесах и кустарниковых зарослях типа чапарраля. Там выделяют группу растений-пирофитов,
устойчивых к пожарам. У деревьев саванн на стволах толстая корка, пропитанная огнеупорными веществами, надежно защищающими внутренние ткани. Плоды и семена пирофитов имеют толстые, часто одревесневшие покровы, которые растрескиваются, будучи опалены огнем.
Некоторые растения регулярно испытывают влияние пожаров, когда температура кратковременно повышается до сотен градусов. Пожары особенно часты в саваннах, в сухих жестколистных лесах и кустарниковых зарослях типа чапарраля. Там выделяют группу растений-пирофитов,
устойчивых к пожарам. У деревьев саванн на стволах толстая корка, пропитанная огнеупорными веществами, надежно защищающими внутренние ткани. Плоды и семена пирофитов имеют толстые, часто одревесневшие покровы, которые растрескиваются, будучи опалены огнем.
 нежаростойкие растения повреждаются")
Слайд 18Действие высоких температур
Для большинства организмов высокие температуры опасны денатурацией ферментов.
температуры до
уровня денатурации приводят к нарушениям равновесия между различными процессами, например, при повышении температуры дыхание идет быстрее, чем фотосинтез, в этих условиях растения испытывают голодание, так как не успевают синтезировать сахара.
На многие организмы высокие температуры действуют косвенно через обезвоживание. Всем наземным организмам приходится беречь воду, быстрая потеря воды при высоких температурах может привести к гибели.
На многие организмы высокие температуры действуют косвенно через обезвоживание. Всем наземным организмам приходится беречь воду, быстрая потеря воды при высоких температурах может привести к гибели.

Слайд 19Действие высоких температур
Обитатели жарких пустынь Африки и Южной Америки способны перенести
температуры, выше порога денатурации ферментов, в этих условиях выживают такие растения как толстянки, алоэ, кактусы.
Доказано, что в цитоплазме таких растений содержатся белки теплового шока, которые защищают ферменты от денатурации. Впервые эти соединения обнаружены в алоэ, способном жить при температуре +80о С. Издавна арабы называли алоэ “сабр”- терпеливый.
Биологически активные вещества алоэ способны ускорить адаптацию организмов к различным негативным факторам окружающей среды, поэтому препараты алоэ используются в медицине в качестве адаптогенов.
Доказано, что в цитоплазме таких растений содержатся белки теплового шока, которые защищают ферменты от денатурации. Впервые эти соединения обнаружены в алоэ, способном жить при температуре +80о С. Издавна арабы называли алоэ “сабр”- терпеливый.
Биологически активные вещества алоэ способны ускорить адаптацию организмов к различным негативным факторам окружающей среды, поэтому препараты алоэ используются в медицине в качестве адаптогенов.

Слайд 20Действие низких температур
При температурах ниже -1оС эктотермы, погибают из-за повреждения клеток
кристаллами льда. Выживающие при этих условиях организмы (а чаще при более низких температурах) могут предотвратить образование льда. Механизмы - физиологические: в цитоплазме и вакуолях растительной клетки увеличивается концентрация растворенных соединений – криопротекторов, с тем, чтобы понизить температуру льдообразования.
Организмы заранее приспосабливаются к наступлению холодов. В их организме накапливаются вещества, обладающие действием антифризов, предотвращающих образование кристаллов льда. Постепенное понижение температуры пропорционально замедляет обмен веществ до полного прекращения. Эктотермные животные впадают в оцепенение- криптобиоз.
Организмы заранее приспосабливаются к наступлению холодов. В их организме накапливаются вещества, обладающие действием антифризов, предотвращающих образование кристаллов льда. Постепенное понижение температуры пропорционально замедляет обмен веществ до полного прекращения. Эктотермные животные впадают в оцепенение- криптобиоз.

Слайд 21Температура для эндотермных животных
Эндотермные организмы в термонейтральной зоне потребляют энергию с
некоторой основной скоростью.
С отклонением температуры в ту или другую сторону животному требуется все больше энергии для поддержания постоянной температуры тела. Даже в оптимальной зоне эндотерму нужно больше энергии, чем эктотермному животному такого же размера.
Поддержание постоянной температуры регулируется и контролируется термостатической системой головного мозга. Обычно температура тела таких животных поддерживается на уровне 35о-40оС.
С отклонением температуры в ту или другую сторону животному требуется все больше энергии для поддержания постоянной температуры тела. Даже в оптимальной зоне эндотерму нужно больше энергии, чем эктотермному животному такого же размера.
Поддержание постоянной температуры регулируется и контролируется термостатической системой головного мозга. Обычно температура тела таких животных поддерживается на уровне 35о-40оС.

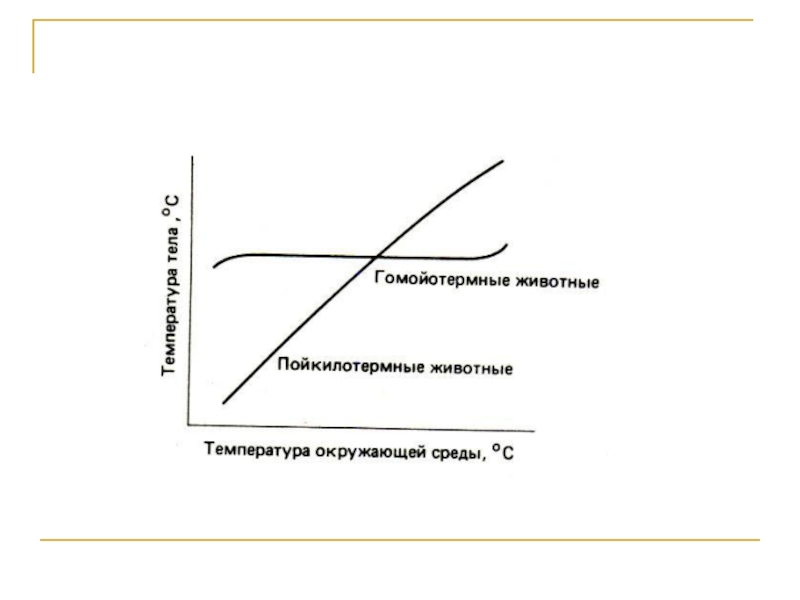
Слайд 22Температура для эндотермных животных
Основные отличия гомойотермных животных от пойкилотермных организмов: 1)
мощный поток внутреннего, эндогенного тепла; 2) развитие целостной системы эффективно работающих терморегуляторных механизмов, и в результате 3) постоянное протекание всех физиологических процессов в оптимальном температурном режиме
Гомойотермные сохраняют постоянный тепловой баланс между теплопродукцией и теплоотдачей и соответственно поддерживают постоянную высокую температуру тела. Организм теплокровного животного не может быть временно «приостановлен» так, как это происходит при гипобиозе или криптобиозе у пойкилотермных.
Гомойотермные сохраняют постоянный тепловой баланс между теплопродукцией и теплоотдачей и соответственно поддерживают постоянную высокую температуру тела. Организм теплокровного животного не может быть временно «приостановлен» так, как это происходит при гипобиозе или криптобиозе у пойкилотермных.
 мощный поток внутреннего, эндогенного")
Слайд 23
Эндотермные животные всегда вырабатывают определенный минимум теплопродукции, обеспечивающий работу кровеносной системы,
органов дыхания, выделения и других, даже находясь в покое. Это базальный метаболизм. Переход к активности усиливает выработку тепла и требует усиления теплоотдачи.

.")

Слайд 26
В пределах некоторого диапазона внешних температур гомойотермные поддерживают температуру тела, не
тратя на это дополнительной энергии, а используя эффективные механизмы физической терморегуляции, позволяющие лучше сохранять или отводить тепло базального метаболизма. Этот диапазон температур, в пределах которого животные чувствуют себя наиболее комфортно, называется термонейтральной зоной. За нижним порогом этой зоны начинается химическая терморегуляция, за верхним – траты энергии на испарение.

Слайд 27
Физическая терморегуляция экологически выгодна, так как адаптация к холоду осуществляется не за
счет дополнительной выработки тепла, а за счет сохранения его в теле животного. Кроме того, возможна защита от перегрева путем усиления теплоотдачи во внешнюю среду.
Способы физической терморегуляции множественны и разнообразными. К ним следует отнести рефлекторное сужение и расширение кровеносных сосудов кожи, меняющее ее теплопроводность, изменение теплоизолирующих свойств меха и перьевого покрова, противоточный теплообмен путем контакта сосудов при кровоснабжении отдельных органов, регуляцию испарительной теплоотдачи.
Способы физической терморегуляции множественны и разнообразными. К ним следует отнести рефлекторное сужение и расширение кровеносных сосудов кожи, меняющее ее теплопроводность, изменение теплоизолирующих свойств меха и перьевого покрова, противоточный теплообмен путем контакта сосудов при кровоснабжении отдельных органов, регуляцию испарительной теплоотдачи.

Слайд 28Противоточная система сохранения тепла внутренних тканей в ласте морской свиньи.
На поперечном
разрезе видно, что артерия полностью окружена мелкими венами. Тепло переходит от нагретой артериальной крови к холодной венозной. Благодаря такому механизму ласт остается холодным, а разница между его температурой и температурой окружающей воды сводится до минимума. (Schmidt–Nielsen, 1975.)

Слайд 29Гетеротермия у эндотермных животных. Температура конечностей арктических птиц и млекопитающих намного
ниже температуры центральной области тела, равной 38°С. (L. Irving. Adaptations to Cold. Copyright)

Слайд 30Тепловая одышка млекопитающих
Вверху. Поток воздуха через нос собаки во время
тепловой одышки. Горизонтальные линии слева от вертикальной черты означают вдох, справа от вертикальной черты–выдох. Средний объем вдоха и выдоха изображен стрелками у носа животного.
Внизу. Поток воздуха через рот собаки во время тепловой одышки. Вдох через рот фактически равен нулю; почти весь воздух, поступивший через нос, выдыхается через рот. (Schmidt–Nielsen et al, 1970.)
Внизу. Поток воздуха через рот собаки во время тепловой одышки. Вдох через рот фактически равен нулю; почти весь воздух, поступивший через нос, выдыхается через рот. (Schmidt–Nielsen et al, 1970.)

Слайд 31
Теплокровным свойственна химическая терморегуляция – рефлекторное увеличение теплопродукции в ответ на понижение температуры
среды. Химическая терморегуляция полностью отсутствует у пойкилотермных, у которых, в случае выделения дополнительного тепла, оно генерируется за счет непосредственной двигательной активности животных.
при действии холода в организме теплокровных животных окислительные процессы не ослабевают, а усиливаются, особенно в скелетных мышцах. У многих животных сначала наблюдается мышечная дрожь – несогласованное сокращение мышц, приводящее к выделению тепловой энергии.
Кроме того, клетки мышечной и многих других тканей выделяют тепло и без осуществления рабочих функций, приходя в состояние особого терморегуляционного тонуса. При дальнейшем снижении температуры среды тепловой эффект терморегуляционного тонуса возрастает.
при действии холода в организме теплокровных животных окислительные процессы не ослабевают, а усиливаются, особенно в скелетных мышцах. У многих животных сначала наблюдается мышечная дрожь – несогласованное сокращение мышц, приводящее к выделению тепловой энергии.
Кроме того, клетки мышечной и многих других тканей выделяют тепло и без осуществления рабочих функций, приходя в состояние особого терморегуляционного тонуса. При дальнейшем снижении температуры среды тепловой эффект терморегуляционного тонуса возрастает.

Слайд 32
Поддержание температуры за счет возрастания теплопродукции требует большого расхода энергии, поэтому
животные при усилении химической терморегуляции либо нуждаются в большом количестве пищи, либо тратят много жировых запасов, накопленных ранее. Например, землеройка бурозубка крошечная имеет исключительно высокий уровень обмена. Чередуя очень короткие периоды сна и активности, она деятельна в любые часы суток и в день съедает корма в 4 раза больше собственной массы. Также и птицам, остающимся на зиму, нужно много корма: им страшны не столько морозы, сколько бескормица.
Усиление химической терморегуляции имеет свои пределы, обусловленные возможностью добывания пищи. При недостатке корма зимой такой путь терморегуляции экологически невыгоден. Он слабо развит у всех животных, обитающих за полярным кругом: песцов, моржей, тюленей, белых медведей, северных оленей и др.
Усиление химической терморегуляции имеет свои пределы, обусловленные возможностью добывания пищи. При недостатке корма зимой такой путь терморегуляции экологически невыгоден. Он слабо развит у всех животных, обитающих за полярным кругом: песцов, моржей, тюленей, белых медведей, северных оленей и др.

Слайд 33Температура для эндотермных животных
Для защиты от неблагоприятного действия как низких, так
и высоких температур организмы вырабатывают адаптации, в первую очередь, морфологические.
правило Бергмана (1848 г.) – в пределах вида или близких видов теплокровные организмы с более крупными размерами тела распространены в более холодных областях, а по мере приближения к теплым - размеры уменьшаются.
правило Бергмана (1848 г.) – в пределах вида или близких видов теплокровные организмы с более крупными размерами тела распространены в более холодных областях, а по мере приближения к теплым - размеры уменьшаются.

Слайд 34правило Бергмана
Чем крупнее животное и компактнее его тело, тем легче поддерживать
постоянную температуру, и наоборот, чем мельче животное, тем больше относительная, излучающая тепло, поверхность его тела, естественно выше уровень основного обмена веществ. Классическим примером, иллюстрирующим правило, являются различия в размерах медведей - самый крупный - белый медведь, меньшие размеры у бурого, меньше всех гималайский.

Слайд 35Правило Бергмана
Если существует род, виды которого отличаются только величиной, тогда более
мелкие виды этого рода будут тяготеть к более теплому климату, причем в точности в соответствии с их массой.
Карл Бергман, 1847
Карл Бергман, 1847
Отношение объема к поверхности (= теплопродукции к теплоотдаче):

Слайд 36Температура для эндотермных животных
Правило Аллена (1877г.)– в холодных областях у
животных с постоянной температурой тела наблюдается тенденция к уменьшению площади выступающих частей тела. Это правило проявляется, например, при сравнении размеров ушей систематически близких видов: песца - обитателя тундры, лисицы обыкновенной - жителя умеренных широт, лисицы-фенека - обитателя пустынь Африки.
– в холодных областях у животных с постоянной температурой")
Слайд 37Правило Аллена
Животные, обитающие в областях с преобладающими низкими температурами, имеют, как
правило, более короткие выступающие части тела (уши, лапы, хвост, нос) по сравнению с обитателями более теплых зон и областей.
Д. Аллен, 1877
Д. Аллен, 1877
Песец
Обыкновенная лисица
Фенек
Заяц-беляк
Толай
Американский заяц

Слайд 38Температура для эндотермных животных
Из правил Аллена и Бергмана есть исключения:
Не
подчиняются этим закономерностям роющие млекопитающие, клювы птиц и эктотермные организмы.
Для пресмыкающихся правило Бергмана “работает” наоборот: размеры их в направлении холодных областей Земли уменьшаются (сравните нильского крокодила и прыткую ящерицу умеренной зоны).
Для пресмыкающихся правило Бергмана “работает” наоборот: размеры их в направлении холодных областей Земли уменьшаются (сравните нильского крокодила и прыткую ящерицу умеренной зоны).

Слайд 39Адаптации к температурному фактору отражены в правиле Глогера, по которому у
теплокровных животных окраска в холодном и сухом климате сравнительно светлее, чем в теплом и влажном.

Слайд 40Поведенческая терморегуляция
Особенности строения нор и расположения гнезд разных видов млекопитающих (по
Н. П. Наумову, 1963, И. И. Барабаш-Никифорову, Л. Н. Формозову, 1963):
1– логово зайца-русака в песчаных дюнах; 2 – снежная нора зайца-русака; 3 – летняя нора полуденной песчанки; 4 – нора малого суслика; 5 – нора выхухоли; 6 – хатка ондатры; 7 – гнезда рыжей полевки в дупле дуба; 8 – зимнее гнездо обыкновенной белки
1– логово зайца-русака в песчаных дюнах; 2 – снежная нора зайца-русака; 3 – летняя нора полуденной песчанки; 4 – нора малого суслика; 5 – нора выхухоли; 6 – хатка ондатры; 7 – гнезда рыжей полевки в дупле дуба; 8 – зимнее гнездо обыкновенной белки

Слайд 41Температура как экологический фактор
Эндотермы придерживаются стратегии, приносящей большие выгоды, но требующей
больших затрат.
Эктотермы используют стратегию с низкими затратами, но с небольшими выгодами.
Как для эктотермов, так и для эндотермов, характерно наличие температурного оптимума, т.е. такой температуры окружающей среды, при которой энергозатраты минимальны, а также верхней и нижней границ губительных температур, за пределами которых способность организма к регуляции температуры тела оказывается явно недостаточной.
По мере удаления от оптимального значения температура среды становится менее благоприятной для длительного проживания, при этом эндотермам приходится расходовать все больше энергии на терморегуляцию.
Эктотермы используют стратегию с низкими затратами, но с небольшими выгодами.
Как для эктотермов, так и для эндотермов, характерно наличие температурного оптимума, т.е. такой температуры окружающей среды, при которой энергозатраты минимальны, а также верхней и нижней границ губительных температур, за пределами которых способность организма к регуляции температуры тела оказывается явно недостаточной.
По мере удаления от оптимального значения температура среды становится менее благоприятной для длительного проживания, при этом эндотермам приходится расходовать все больше энергии на терморегуляцию.






