самовоспроизведение – двухфазный процесс: образование структуры на основе вложенной в нее информации, и копирование информации с помощью образуемой на ее основе структуры (Дж. Фон Нейман). Идентичность особей поколений n и n+1 гарантируется только идентичностью информации и правил ее считывания. При этом устраняется влияние структуры в поколении n на структуру в поколении n+1 (Вейсман). «Самовоспроизведение» превращается в образование другой структуры, которая совпадает с предыдущей только из-за использования одной и той же информации.
Аналог – письменная культура
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Онтогнез. Периоды онтогенеза презентация
Содержание
- 1. Онтогнез. Периоды онтогенеза
- 2. Элементарной единицей, способной к консервативному самовоспроизведению, и,
- 3. Для того чтобы шел отбор по приспособленности
- 4. Для неконсервативного воспроизведения ключевым является понятие сериальной
- 5. Образование и последовательное воспроизведение модулей структуры основано
- 6. «МОРФОГЕНЕТИЧЕСКИЙ РОСТ» РАСТИТЕЛЬНОГО ПОБЕГА Рост апикальной меристемы
- 7. НЕКОНСЕРВАТИВНОЕ ВОСПРОИЗВЕДЕНИЕ МОДУЛЕЙ СТРУКТУРЫ – ОСНОВА МОРФОГЕНЕЗА
- 8. Морфогенез создает точки бифуркации, а с ними
- 9. НЕКОНСЕРВАТИВНОЕ ВОСПРОИЗВЕДЕНИЕ ТРЕБУЕТ, ЧТОБЫ В ПОКОЛЕНИИ N+1
- 10. ОДНОКЛЕТОЧНЫЕ ВОЛОСКИ КРУПОК (КРЕСТОЦВЕТНЫЕ РОДА DRABA) –
- 11. Образование доменной структуры (A), затухание роста (B)
- 12. Варьируя величину вклада пассивных (упругих) напряжений в
- 13. ПРЕВРАЩЕНИЕ МОРФОГЕНЕЗА СТРУКТУРЫ В ЕЕ ОНТОГЕНЕЗ БЕЗ
- 14. На листовой пластинке осцилляции формы растущего волоска
- 15. Упорядочение (канализация) изменчивости с возникновением онтогенеза может
- 16. У инфузории Tetrachymena соматическое размножение устроено точно
- 17. МОДЕЛЬНЫМ ПРИМЕРОМ ВОЗНИКНОВЕНИЯ И ЭВОЛЮЦИИ ОНТОГЕНЕЗА ЯВЛЯЕТСЯ
- 18. Простейший онтогенез на клеточном уровне – метазойный
- 19. Эволюция онтогенеза Protozoa связана с полимеризацией субклеточных
- 20. Инфузории (Tetrahymena) – вершина эволюции онтогенеза полимерных
- 21. Варианты (индивидуальная изменчивость) чередования фаз жизненного цикла
- 23. Передний конец будущего стебля – физиологически активная часть плазмодия, будущая споровая часть пассивна
- 24. Жизненный цикл социальных амеб (Dyctiostelium) – модельный
- 25. * - клетки в автоколебательном режиме секреции
- 26. Изменчивы число и размер агрегатов, и соотношение
- 27. I – постоянная и слабая секреция цАМФ,
- 28. В агрегации клеток Dyctiostelium действуют те же
- 29. НЕЗАВИСИМОСТЬ ОТ ФИЛОГЕНЕЗА У растений и
- 30. ВОЗНИКНОВЕНИЕ ОНТОГЕНЕЗА (ЖИЗНЕННОГО ЦИКЛА) METAZOA A –
- 31. ЭВОЛЮЦИОННАЯ УСТОЙЧИВОСТЬ ОНТОГЕНЕЗА METAZOA: память об эволюционном
- 32. Эволюционно исходной биологической формой онтогенеза является чередование
- 33. ЭПИТЕЛИО-МЕЗЕНХИМАЛЬНЫЙ ЦИКЛ У КНИДАРИЙ Жгутиковая бластула состоит
- 34. В КОНЦЕ ЭВОЛЮЦИИ У ПОЛИПА ПОЯВЛЯЕТСЯ СОБСТВЕННЫЙ ОНТОГЕНЕЗ
- 35. В КОНЦЕ ЭВОЛЮЦИИ У МЕДУЗЫ ПОЯВЛЯЕТСЯ СОБСТВЕННЫЙ ОНТОГЕНЕЗ
- 36. Общая схема перехода от чередования фаз к
- 37. На языке взаимодействия генов необходимыми и достаточными
- 38. КРОСС-ИНГИБИЦИЯ (блок переключения онтогенеза у фага λ
- 39. Уравнение кросс-ингибиции, объясняющее, зачем регулятор транскрипции и
- 41. РЕТРОГРАДНАЯ ЭВОЛЮЦИЯ КАТАЛИТИЧЕСКОЙ ЦЕПИ КОМПЕНСИРУЕТ УХУДШЕНИЕ СРЕДЫ
- 42. Эпигенетический код дифференцировки Эукариоты соединяют (+)
Слайд 1Онтогенез – это самовоспроизведение особи, имеющее консервативную и неконсервативную компоненту. Консервативное

Слайд 2Элементарной единицей, способной к консервативному самовоспроизведению, и, как следствие, селекционной эволюции
является гиперцикл, где J – информация, E – структура
ОТБОР: dJ/dt = (AQ)J, где A – скорость, Q – точность репликации i-той матрицы, а так как чем больше доля таких матриц, тем точнее их воспроизведение, Q = F(J) >0, и dJ/dt = A(F(J)J. A и Q – компоненты приспособленности Wi: даже в простейшем гиперцикле приспособленность раскладывается на компоненты

Слайд 3Для того чтобы шел отбор по приспособленности (дарвиновский отбор), необходимо, чтобы
механизм самовоспроизведения особи (гиперцикла) либо вовсе не менялся, либо менялся гораздо медленнее, чем относительные частоты особей с разными значениями A и Q. Это условие может нарушаться, как при эволюции РНКового фага Q-бета в бесклеточной среде
В бесклеточной среде размер РНК фага уменьшается в 5 раз за несколько циклов репликации за счет элиминации всех участков РНК, не нужных для ее опознавания полимеразой фага. Приспособленность вообще не оценивается, но ЛЮБЫЕ короткие нуклеотидные последовательности получают преимущество, поскольку их репликация идет быстрее. Требуется только, чтобы они содержали ген полимеразы и палиндромы, позволяющие полимеразе присоединиться к РНК фага и начать транскрипцию.
ЭТО НЕ ОТБОР, ТАК КАК ВОСПРОИЗВЕДЕНИЕ ПЕРЕСТАЕТ БЫТЬ КОНСЕРВАТИВНЫМ
ЕСЛИ, КАК ПРИ ФОРМИРОВАНИИ ГЕНЕТИЧЕСКОГО ХОДА, ИДЕТ ОТБОР (ИЛИ ВЫБОР) МЕХАНИЗМА САМОВОСПРОИЗВЕДЕНИЯ, ТО КАЧЕСТВО (ТОЧНОСТЬ) ВОСПРОИЗВЕДЕНИЯ СТАНОВИТСЯ ОДНОЙ ИЗ КОМПОНЕНТ ПРИСПОСОБЛЕННОСТИ ФЕНОТИПА
, необходимо, чтобы механизм самовоспроизведения особи (гиперцикла)")
Слайд 4Для неконсервативного воспроизведения ключевым является понятие сериальной (модульной) гомологии: развитие модуля
структуры создает условия и вектор развития ее сериального модуля. Аналог – бесписьменная культура
Все конечности позвоночных исторически гомологичны, а их структурные блоки сериально гомологичны
Сериальная гомология означает параметрическую модификацию развития, ее отсутствие – смену программы развития структуры
«Программа развития» - уравнение, описывающее конденсацию мезенхимы (клетки движутся вверх по градиенту концентрации вдоль нормальных радиусов кривизны наружной поверхности). Параметр – форма поверхности поперечного сечения почки конечности. По мере ее роста сечение из округлого (А) становится эллиптическим (Б), и вместо одного возникает два центра конденсации, а при дальнейшем уплощении эллипса сегменты мезенхимы формируются вдоль оси роста
 гомологии: развитие модуля структуры создает условия и")
Слайд 5Образование и последовательное воспроизведение модулей структуры основано на действии циклических алгоритмов
морфогенеза, применяемых к реальным клеткам, или гипотетическим «морфогенам»
Двойная стрелка – движение волны сомитогенеза, одинарная стрелка – движение клеток против градиента концентрации (навстречу движению волны). Алгоритм воспроизведения подразумевает обратную связь развития сомита n с формированием программы развития следующего сомита n+1
Реакционно-диффузионная система Тьюринга - Гирера - Мейнхарда
Образование доменной структуры виде пиков концентрации активатора и ингибитора отражает структуру их взаимодействия
Повторное образование доменов – логическое следствие механизма образования доменной структуры

Слайд 6«МОРФОГЕНЕТИЧЕСКИЙ РОСТ» РАСТИТЕЛЬНОГО ПОБЕГА
Рост апикальной меристемы побега связан с периодическими изменениями
формы: избыток внутреннего давления «сбрасывается» путем усложнения формы поверхности: образования модулей в виде сначала листовых (А), а затем цветочных зачатков (Б)
Модель Сакса: для развития апикальной меристемы и пазушных почек нужно сначала повышение локальной концентрации ауксина (приток ауксина), а потом ее снижение (отток ауксина) с помощью поляризованного транспорта (белки PIN)

Слайд 7НЕКОНСЕРВАТИВНОЕ ВОСПРОИЗВЕДЕНИЕ МОДУЛЕЙ СТРУКТУРЫ – ОСНОВА МОРФОГЕНЕЗА ПОКРЫТОСЕМЕННЫХ РАСТЕНИЙ (КРУПКИ)
Морфогенез
всего растения – последовательная редукция начальных стадий развития его боковых побегов, развивающихся из пазушных почек: 1 и 2 – побеги возобновления, копирующие начало морфогенеза растения; 3-5 – побег образует флоральную меристему на более ранней стадии собственного развития (потому что он развивается уже как часть всего растения); 6 – от побега остается только цветок, развивающийся на материнском растении; 7 - образование соцветия, цветок – предел развития как крайняя степень редукции побега. Одинарные стрелки – онтогенез растения (c последовательным сокращением ранних стадий развития бокового побега), двойные стрелки – апикальное доминирование (снижающееся по мере сокращения онтогенеза боковых побегов)
Различие последовательных побегов не наследственно, но организация их изменчивости различна, а значит – различен и их ответ на отбор
 Морфогенез всего растения – последовательная")
Слайд 8Морфогенез создает точки бифуркации, а с ними формы, которые могут быть
разными организмами, или разными частями одного организма – морфогенезу это все равно. Путь от морфогенеза к онтогенезу – превращение внутри-индивидуальных различий в различия организмов, или в последовательные стадии развития особи
Морфогенез является неконсервативной компонентой самовоспроизведения из-за бифуркации пути развития при образовании дочерних особей (или дочерних структур)
В простейшем случае физического разделения областей на две одинаковые области в какой-то момент (точка бифуркации) возникают два зачатка с равными радиусами R1 и R2, имеющими единый центр (А). Это «негрубое положение», которое при «малом шевелении» формы превращается в «грубое» различие дочерних областей I и II (Б).

Слайд 9НЕКОНСЕРВАТИВНОЕ ВОСПРОИЗВЕДЕНИЕ ТРЕБУЕТ, ЧТОБЫ В ПОКОЛЕНИИ N+1 ПОВТОРЯЛИСЬ УСЛОВИЯ РАЗВИТИЯ СТРУКТУРЫ
В ПОКОЛЕНИИ N, ЗАМЕНЯЮЩИЕ ЕЕ ФИЗИЧЕСКОЕ ПРИСУТСТВИЕ
Обычный путь перехода от морфогенеза к онтогенезу структуры – смещение асимметрии зачатков I и II на более ранние стадии развития материнской области, предшествующие появлению самих зачатков. Асимметрия должна поддерживаться асимметрией окружения области, а векторы развития (одинарная стрелка) и эволюции (двойная стрелка) имеют взаимно противоположное направление. Вместо бифуркации получается детерминированная последовательность стадий развития
Направление эволюции НЕ зависит от направления отбора: достаточно, чтобы он изменял предшествующее появлению зачатков развитие области, не снижая общей приспособленности

Слайд 10ОДНОКЛЕТОЧНЫЕ ВОЛОСКИ КРУПОК (КРЕСТОЦВЕТНЫЕ РОДА DRABA) – ПРОСТАЯ МОДЕЛЬ ВОЗНИКНОВЕНИЯ ОНТОГЕНЕЗА
ИЗ НЕКОНСЕРВАТИВНОГО САМОВОСПРОИЗВЕДЕНИЯ
Начало развития волосков из волосковых клеток у разных видов крупок (A – F)
«Морфогенетический рост» волосковой клетки: рельеф поверхности – запись ростовых осцилляций
Рост волосковой клетки – автоколебательный цикл с чередованием роста и уменьшения отношения поверхности к объему апекса (зоны роста поверхности). Возникновение повторяющихся модулей волоска – прямое следствие автоколебаний (осцилляций), независимо от формы роста.
 – ПРОСТАЯ МОДЕЛЬ ВОЗНИКНОВЕНИЯ ОНТОГЕНЕЗА ИЗ НЕКОНСЕРВАТИВНОГО САМОВОСПРОИЗВЕДЕНИЯ Начало")
Слайд 11Образование доменной структуры (A), затухание роста (B) и собственно осцилляции (C)
возникают при параметрической модификации одного и того же уравнения движения растущей поверхности апекса
, затухание роста (B) и собственно осцилляции (C) возникают при параметрической модификации")
Слайд 12Варьируя величину вклада пассивных (упругих) напряжений в изменение формы апекса (А),
или начальную геометрию волосковой клетки (Б), получаем весь спектр возможных форм волоска у крупок
Переход от однолучевого к ветвящемуся волоску получается при уменьшении вклада пассивных напряжений (увеличении темпов роста поверхности апекса)
Различие между однолучевым (верхний ряд) и ветвящимся волоском получается из-за различия латерального диаметра апекса волосковой клетки (из-за конкуренции между латеральным ростом апекса и ростом волоска вдоль его главной оси)
 напряжений в изменение формы апекса (А), или начальную геометрию волосковой")
Слайд 13ПРЕВРАЩЕНИЕ МОРФОГЕНЕЗА СТРУКТУРЫ В ЕЕ ОНТОГЕНЕЗ БЕЗ ИЗМЕНЕНИЯ ГЕНОТИПА
На цветоносе
крупки Draba sibirica плотная упаковка клеток эпидермиса выдавливает волосковые клетки из клеточного пласта (а). Образуются образуются однолучевые (б, в) или двулучевые волоски (г). Стадии развития волоска отсутствуют, есть только чередование расширения и сужения верхушечной зоны роста (осцилляции)
Выбор между образованием однолучевого или двулучевого волоска зависит только от случайных флюктуаций диаметра верхушечной зоны

Слайд 14На листовой пластинке осцилляции формы растущего волоска превращаются в последовательность стадий
нормального развития (1 → 4) четырехлучевых волосков, диагностических для D. sibirica
PD – проксимодистальная ось листа, * - «быстрый» зачаток первого луча, направленный к P полюсу (против вектора роста листа), X – «усомнившийся» волосок со слабой латеральной полярностью
Латеральная полярность волосковой клетки (клетка 1) возникает еще до оформления «быстрого» зачатка луча
Появление у волосковой клетки онтогенеза объясняется изменением условий развития: возникает отрицательная корреляция между ростом поверхности волосковой клетки и ростом площади поверхности листовой пластинки, отсутствующая на цветоносе из-за его цилиндрической формы

Слайд 15Упорядочение (канализация) изменчивости с возникновением онтогенеза может происходить не только в
ответ на изменение условий развития, но и на генетической основе
Плезиоморфный вид D. nivalis: глядя на изменчивость развивающихся волосков, невозможно выделить линейную серию стадий развития
Апоморфный вид D. nemorosa: для выделения стадий развития (1→4) достаточно взглянуть на изменчивость, не прослеживая развитие отдельных волосков
Стрелка – изменение формы роста, * - «быстрый» зачаток луча, 5 – волосок с «лишним» зачатком луча
Так как форма волоска не имеет адаптивного значения, возникновение ее онтогенеза не зависит от направления отбора
 изменчивости с возникновением онтогенеза может происходить не только в ответ на изменение условий")
Слайд 16У инфузории Tetrachymena соматическое размножение устроено точно так же, как неконсервативное
воспроизведение модулей в реакционно-диффузионной системе. Сигнал, исходящий от A-полюса, является близкодействующим активатором и дальнодействующим ингибитором его развития. Когда размер инфузории превышает поле действия ингибитора, начинается новый виток морфогенетического цикла
Биологической единицей является не особь, а клон, где все особи ранжированы по возрасту A-полюса (ротового аппарата)

Слайд 17МОДЕЛЬНЫМ ПРИМЕРОМ ВОЗНИКНОВЕНИЯ И ЭВОЛЮЦИИ ОНТОГЕНЕЗА ЯВЛЯЕТСЯ ВОЗНИКНОВЕНИЕ ЖИЗНЕННОГО ЦИКЛА И
ОНТОГЕНЕЗА METAZOA
Все Metazoa имеют метазойный митоз (всего одна центросома). Это доказывает единство происхождения Metazoa от примитивных Protozoa с метазойным митозом и дает ключ к пониманию причин эволюции. ОСНОВНАЯ «ЦЕЛЬ» ЭВОЛЮЦИИ – УВЕЛИЧЕНИЕ РАЗМЕРА ОСОБИ. У примитивных Protozoa есть всего два пути увеличения размера особей, оба связанные с полимеризацией. Один из них – полимеризация внутриклеточных органелл (эволюция высших Protozoa), а тем организмам, у которых в клетке так и остается один митотический центр, остается полимеризоваться за счет увеличения числа клеток
ВОЗНИКНОВЕНИЕ METAZOA – ЭТО ВОЗНИКНОВЕНИЕ НОВОГО МЕХАНИЗМА САМОВОСПРОИЗВЕДЕНИЯ: ЕГО СУТЬ – ВЫНУЖДЕНИЕ ОТКАЗА СОМАТИЧЕСКИХ КЛЕТОК ОТ СОБСТВЕННОГО ВОСПРОИЗВЕДЕНИЯ (ВОЗНИКНОВЕНИЕ СОЦИАЛЬНОСТИ)

Слайд 18Простейший онтогенез на клеточном уровне – метазойный клеточный цикл. Он включает
в себя морфогенетический цикл – чередование потери и возобновления клеточной полярности (AP различий) в каждой дочерней клетке независимо от полярности материнской клетки
Одна из дочерних клеток ближе к материнской клетке, так как в ней находится исходный (материнский) митотический центр

Слайд 19Эволюция онтогенеза Protozoa связана с полимеризацией субклеточных органелл (энергид) и переходом
к клональному самовоспроизведению
При протозойном митозе все потомки данной клетки имеют общую AP полярность, единую для всего клона дочерних клеток (все клетки «плывут в одну и ту же сторону»)
Циклический алгоритм чередования полярной и биполярной формы клетки остается неизменным при всех модификациях клеточного цикла (например, при выпадении G1 или G2 фазы)
 и переходом к клональному самовоспроизведению При")
Слайд 20Инфузории (Tetrahymena) – вершина эволюции онтогенеза полимерных Protozoa. Реснички становятся морфогенетическими
аналогами клеток Metazoa, митоз – сложным морфогенезом, а модификации фенотипа клонируются так же, как и сам фенотип
 – вершина эволюции онтогенеза полимерных Protozoa. Реснички становятся морфогенетическими аналогами клеток Metazoa, митоз")
Слайд 21Варианты (индивидуальная изменчивость) чередования фаз жизненного цикла раковинных амеб Foraminifera: *
- палинтомия, ** - мейоз, *** - рост
РОСТ МНОГОЯДЕРНОЙ РАКОВИННОЙ АМЕБЫ
Чередование монотомии и расселения
Чередование монотомии и инцистирования
Стрелки – точки бифуркации цикла
 чередования фаз жизненного цикла раковинных амеб Foraminifera: * - палинтомия, ** -")
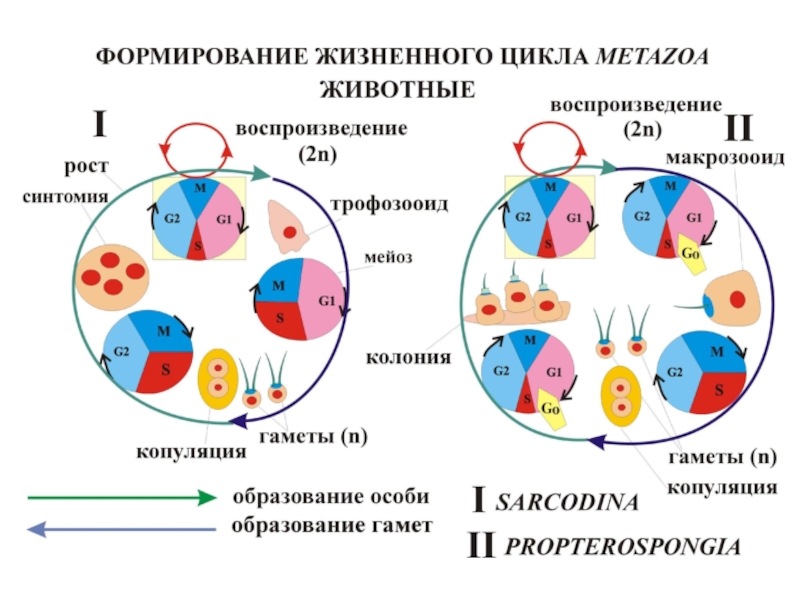
Слайд 23Передний конец будущего стебля – физиологически активная часть плазмодия, будущая споровая
часть пассивна

Слайд 24Жизненный цикл социальных амеб (Dyctiostelium) – модельный пример возникновения онтогенеза не
в эволюции, а в самом онтогенезе
Фаза самоорганизации (мелкомасштабная агрегация клеток). В основе – (+) обратная связь между ростом локальной концентрации клеток и цАМФ. Размер агрегатов порядка сотен клеток, их расположение беспорядочно. Работают те же гены, что у одиночных амеб, межвидовые различия отсутствуют. При дальнейшем голодании клеток агрегаты рассасываются, и их распределение снова становится равномерным
Фаза крупномасштабной агрегации (онтогенез). Тоже (+) обратная связь между концентрацией клеток и цАМФ, но цАМФ испускается квантами с рефрактерным периодом. Формируется единый агрегат из десятков тысяч клеток, есть межвидовые различия, включаются гены, не работающие у одиночных амеб.
Клетки движутся навстречу движению фронта волны цАМФ
Вся последовательность событий зависит от изменения единственного параметра – сигнала клеточного голода
 – модельный пример возникновения онтогенеза не в эволюции, а в")
Слайд 25* - клетки в автоколебательном режиме секреции АМФ (гликоген почти весь
израсходован)
** - клетки в ждущем режиме секреции АМФ (гликоген еще есть)
*** - клетки в режиме слабой постоянной секреции АМФ (много гликогена)
** - клетки в ждущем режиме секреции АМФ (гликоген еще есть)
*** - клетки в режиме слабой постоянной секреции АМФ (много гликогена)
Пространственная картина агрегации – развертка последовательных состояний клетки по мере ее голодания. Чем голоднее клетки, тем сильнее их агрегация. В центре агрегата самые голодные клетки, образующие лидирующую часть плазмодия (будущие стеблевые клетки)
 ** - клетки")
Слайд 26Изменчивы число и размер агрегатов, и соотношение размера преспоровой и престеблевой
части. Отбор в пользу более длинного стебля эффективен, несмотря на отсутствие мейоза!

Слайд 27I – постоянная и слабая секреция цАМФ, II – режим автоколебаний,
III – угасание секреции.
C – аденилатциклаза (активатор), H – глюкоза (ингибитор). Активатор образуется с помощью автокатализа (цАМФ – транскрипционный фактор), производит ингибитор (активируется распад гликогена с образованием глюкозы), ингибитор подавляет автокатализ активатора (продукты распада гликогена ингибируют активность аденилатциклазы)
C – аденилатциклаза (активатор), H – глюкоза (ингибитор). Активатор образуется с помощью автокатализа (цАМФ – транскрипционный фактор), производит ингибитор (активируется распад гликогена с образованием глюкозы), ингибитор подавляет автокатализ активатора (продукты распада гликогена ингибируют активность аденилатциклазы)
Агрегация – прямой ответ клеток на голодание без участия специальных «морфогенетических сигналов»
C (цАМФ) – активатор, H (глюкоза) - ингибитор

Слайд 28В агрегации клеток Dyctiostelium действуют те же факторы, что при агрегации
бактерий, не создающей адаптивных структур. Разница только в масштабе и автоколебаниях
Сегрегация и морфогенез престеблевой и преспоровой зоны – классическая реакционно-диффузионная система, где близкодействующий активатор – белок Dif, закупоривающий протоновые каналы и закисляющий цитоплазму, а дальнодействующий ингибитор – аммиак. Не меньшую роль может играть то, что «голодные» и «сытые» клетки разделяются в ходе своего движения вверх по градиенту концентрации клеток, что создает дискретные различия в концентрации цАМФ
В любом случае морфогенез, основанный на движении клеток НАВСТРЕЧУ движению волн цАМФ, придает агрегации макроскопический масштаб, создавая осцилляции, а с ними онтогенез, включая возможность возникновения межвидовых различий

Слайд 29НЕЗАВИСИМОСТЬ ОТ ФИЛОГЕНЕЗА
У растений и социальных амеб многоклеточность возникает иначе, чем
у животных, но общей чертой является социальность поведения клеток (отказ физиологически активной части клеток от размножения)

Слайд 30ВОЗНИКНОВЕНИЕ ОНТОГЕНЕЗА (ЖИЗНЕННОГО ЦИКЛА) METAZOA
A – исходное чередование палинтомического (N) и
монотомического (N+1) поколений Protozoa: 1 – палинтомия, 2 – рост, 3 – образование, 4 – сброс жгутика; B – объединение циклов с неустойчивой точкой переключения (звездочка); C – онтогенез Metazoa. Два поколения жгутиковых клеток в одном организме (жгутиковые клетки бластулы и жгутиковые клетки энтодермы, у губок - хоаноциты), гаструляция ~ метаморфоз.
 METAZOAA – исходное чередование палинтомического (N) и монотомического (N+1) поколений Protozoa:")
Слайд 31ЭВОЛЮЦИОННАЯ УСТОЙЧИВОСТЬ ОНТОГЕНЕЗА METAZOA: память об эволюционном происхождении сохраняется, несмотря на
дивергенцию
Двойная стрелка – монотомия, одинарная стрелка - палинтомия
Амфибии и высшие насекомые – крайне удаленные группы с совершенно различной морфологией и биологией эмбрионального развития. Чередование монотомии и палинтомии (митозов с G1 и без G1 фазы) – их общая черта в память об эволюционном происхождении многоклеточности
Память о происхождении нельзя считать просто генетической памятью, так гены (циклины), переключающие фазы клеточного цикла, действуют и в жизненных циклах Protozoa

Слайд 32Эволюционно исходной биологической формой онтогенеза является чередование поколений (метагенез). Онтогенез в
виде линейной последовательности сменяющих друг друга стадий развития является производной формой чередования альтернативных фаз жизненного цикла
ПРИНЦИП ДИВЕРГЕНЦИИ
Дивергенция означает переход от индивидуальных к типологическим различиям: от различия особей (или частей организма) к различию регулярно воспроизводимых типов особей. Это наиболее общая закономерность биологической эволюции – селекционной или не селекционной
Простейший пример – чередование палинтомических (без G1 фазы) и монотомических (с G1 фазой) делений одноклеточного организма с последующим выпадением палинтомических делений
. Онтогенез в виде линейной последовательности сменяющих")
Слайд 33ЭПИТЕЛИО-МЕЗЕНХИМАЛЬНЫЙ ЦИКЛ У КНИДАРИЙ
Жгутиковая бластула состоит из неполноценных клеток, которым, чтобы
закончить клеточный цикл, нужно сбросить жгутики. Получается сплошная масса мезенхимных клеток, формирующих второе поколение эпителия уже из клеток с G1 фазой. Это эпителио-мезенхимальный цикл (EMC), в который вовлечены все клетки (у некоторых губок), или только часть клеток бластулы
Изменчивость связана с гетерохрониями между динамикой EMC и морфогенезом ЭП. Фиксация гетерохроний – с фиксацией пути развития на таксономическом уровне за счет усиления связи EMC с морфогенезом
В качестве «типа онтогенеза» может фиксироваться любая комбинация переходов по вертикали и горизонтали, определяющих исходное «поле изменчивости»



Слайд 36Общая схема перехода от чередования фаз к линейному онтогенезу:
Одна из дефинитивных
форм (медузоид или гидрант) оказывается более приспособленной → На стадиях, предшествующих точке ветвления пути развития (точка бифуркации) форма изменяется в сторону большего сходства с более приспособленной дефинитивной формой (медузоидом или гидрантом) → Точка бифуркации исчезает, и эта форма становится единственной дефинитивной формой
Значит, стадии развития, предшествующие выбору и фиксации дефинитивной формы возникают в эволюции позже, чем она сама, а изменение предшествующих по времени стадий ее фиксирует. Это и есть ретроградная эволюция морфогенеза, ведущая к возникновению онтогенеза как линейной последовательности событий
")
Слайд 37На языке взаимодействия генов необходимыми и достаточными условиями возникновения онтогенеза являются:
Минус-регуляция:
перекрестное (-) взаимодействие генов (кросс- ингибиция), определяет бифуркацию пути развития (дифференцировку областей развивающейся системы), но не выбор пути, зависящий от условий среды (неконсервативное воспроизведение)
(2) Плюс-регуляция: из-за плохих промоторов транскрипция гена нуждается в действии дополнительных генов, и возникает замкнутая последовательность действия генов – последовательность стадий развития области, не требующая внешней регуляции (консервативное воспроизведение)
 взаимодействие генов")

Слайд 39Уравнение кросс-ингибиции, объясняющее, зачем регулятор транскрипции и мембранным рецептор нужна димеризация
При нелинейной динамике X и Y эффективность отбора падает!

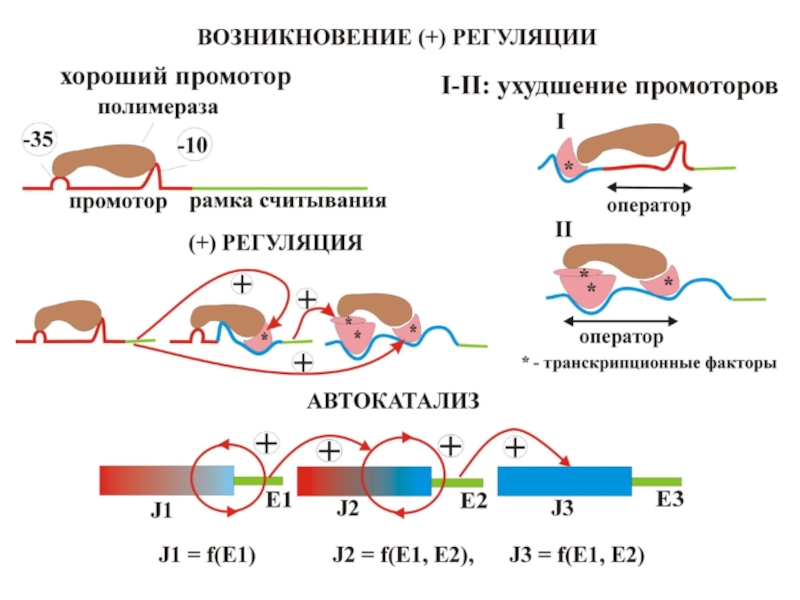
Слайд 41РЕТРОГРАДНАЯ ЭВОЛЮЦИЯ КАТАЛИТИЧЕСКОЙ ЦЕПИ КОМПЕНСИРУЕТ УХУДШЕНИЕ СРЕДЫ ИЛИ ПРОМОТОРОВ ЗА СЧЕТ
УХУДШЕНИЯ КОНСТАНТЫ МИХАЭЛИСА
В отличие от предыдущих рисунков, красный цвет – плохая часть промотора

Слайд 42Эпигенетический код дифференцировки
Эукариоты соединяют (+) и (-) регуляцию для эпигенетического кодирования
областей зародыша. В развитии дрозофилы области различаются последовательностью пройденных альтернативных состояний (условно – нулей и единиц), создаваемых бифуркациями
 и (-) регуляцию для эпигенетического кодирования областей зародыша. В развитии")