- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Общий путь катаболизма. (Лекция 9) презентация
Содержание
- 1. Общий путь катаболизма. (Лекция 9)
- 2. Общий путь катаболизма включает: окислительное
- 3. Стадии катаболических превращений ПВК (пируват) образуется
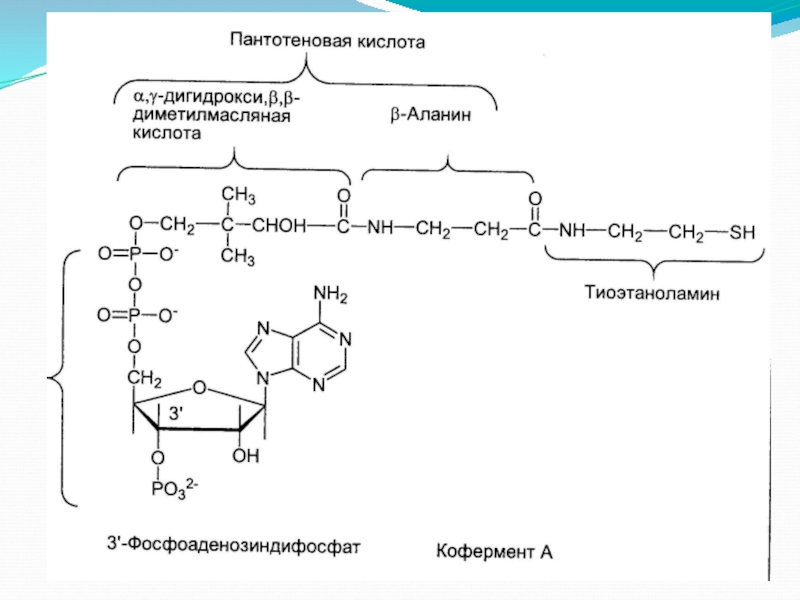
- 6. Пируватдегидрогеназа (Е1, ПВК-ДГ), ее коферментом является тиаминдифосфат
- 7. II этап На втором этапе происходит взаимодействие
- 8. II этап - продолжение III этап
- 9. Регуляция пируватдегидрогеназного комплекса Превращение пирувата в ацетил-КоА
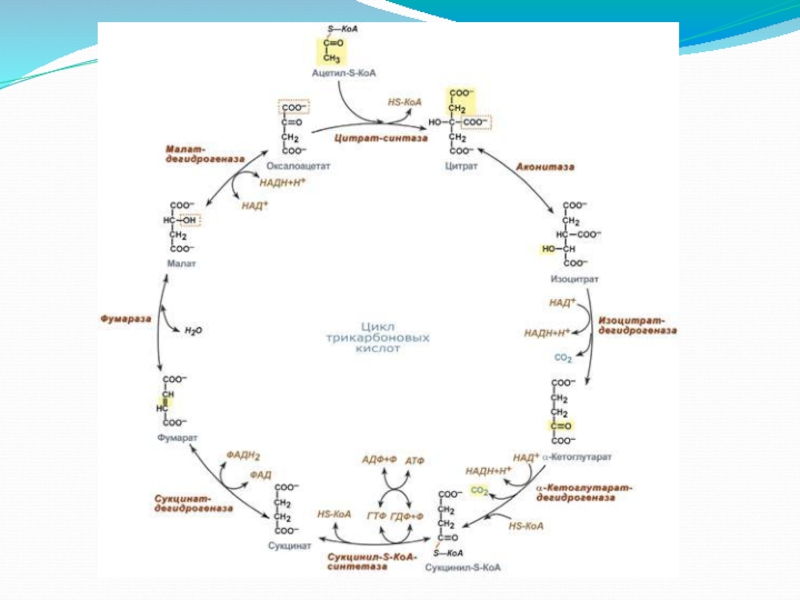
- 14. ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ
- 16. Реакции лимонного цикла 1-я реакция цикла Свободную
- 17. Реакции лимонного цикла 2-я реакция цикла Аконитат-гидратаза
- 18. Реакции лимонного цикла 3-я реакция цикла Изоцитратдегидрогеназа
- 19. Реакции лимонного цикла 4-я реакция цикла 2-Оксоглутаратдегидрогеназный
- 20. Реакции лимонного цикла 5-я реакция цикла Сукцинилтиокиназа
- 21. Реакции лимонного цикла 6-я реакция цикла Сукцинатдегидрогеназа
- 22. Реакции лимонного цикла 7-я реакция цикла Фумарат-гидратаза
- 23. Реакции лимонного цикла 8-я реакция цикла Малатдегидрогеназа
- 25. Интегративная функция — цикл является связующим звеном между
- 26. Анаплеротические реакции Анаплероmuческuе (пополняющие) реакции – специальные
- 27. Ферментативное карбоксилирование пирувата Наиболее важная анаплеротическая реакция
- 28. Зависимое от АТP и биотина карбоксилирование
- 29. Глиоксилатный путь окисления для пополнения ЦТК 9у растений, грибов и бактерий)
- 30. Регуляция цикла Кребса на уровне цитратсинтазы,
- 31. Цикл лимонной кислоты – это один из
- 32. Участие кислот ЦТК в анаболизме Роль кислот ЦТК в анаболизме

Слайд 2Общий путь катаболизма включает:
окислительное декарбоксилирование пирувата до ацетил-КоА;
окисление ацетильной
выделение и аккумулирование энергии при дегидрировании метаболитов общего пути катаболизма в митохондриальных цепях переноса электронов (рассмотрели на предыдущей лекции).

Слайд 3Стадии катаболических превращений
ПВК (пируват) образуется из углеводов(глюкоза), глицерола, гликоген-ных аминокислот и
Ацетил-КоА занимает централь-ное место в общем пути катаболизма и образуется в мито- хондриях: 1) при окислительном декарбоксилировании пирувата; 2) при β- окислении жирных кислот; 3) из кето-генных аминокислот.
 образуется из углеводов(глюкоза), глицерола, гликоген-ных аминокислот и лактата. Ацетил-КоА занимает централь-ное")
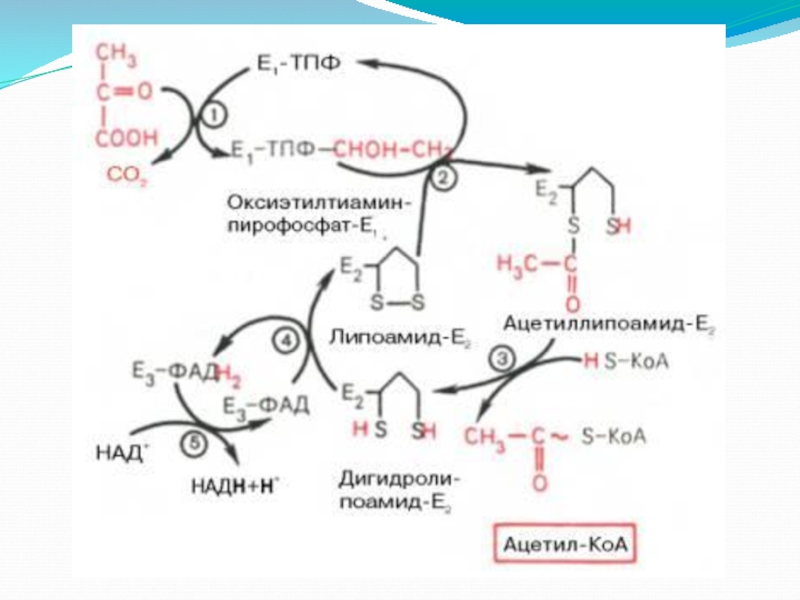

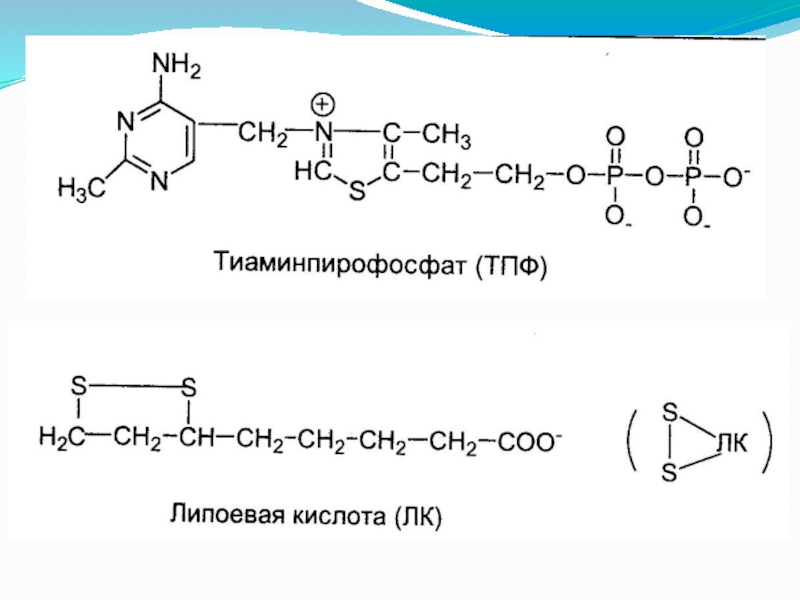
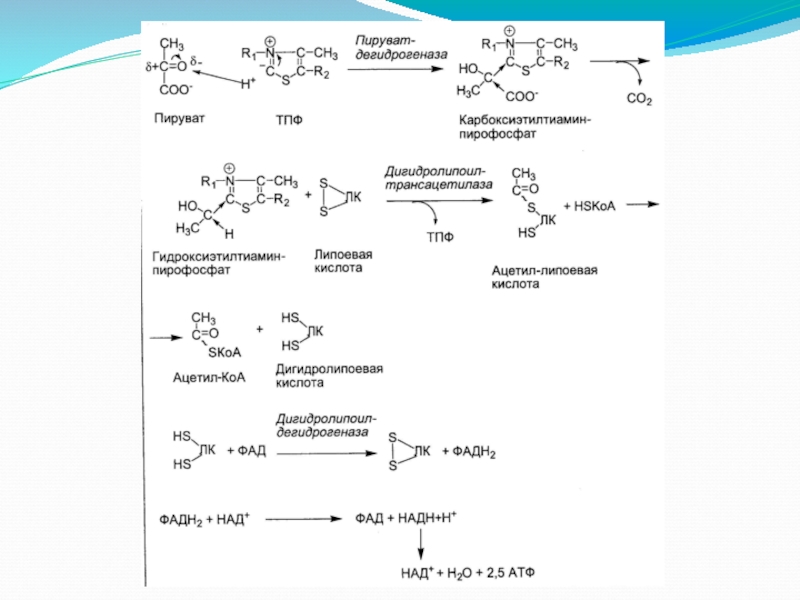
Слайд 6Пируватдегидрогеназа (Е1, ПВК-ДГ), ее коферментом является тиаминдифосфат (ТДФ) или тиаминпирофосфат (ТПФ),
I этап
ПВК взаимодействует с кофактором тиаминдифосфатом. Основную роль играет второй углеродный атом тиазольного кольца ТДФ, который легко теряет протон, превращаясь в карбанион. Карбанион атакует частично положительно заряженный α-углеродный атом пирувата с возникновением связи С-С. Сильно электрофильный атом азота в карбоксиэтил-ТДФ способствует его декарбоксилированию с образованием гидроксиэтил-ТДФ .
, ее коферментом является тиаминдифосфат (ТДФ) или тиаминпирофосфат (ТПФ), катализирует 1-ю реакцию.I этапПВК")
Слайд 7II этап
На втором этапе происходит взаимодействие гидроксиэтил-ТДФ с липоевой кислотой. Происходит
ЛК полностью восстанавливается (III этап). Образованный ацетил~SКоА отделяется от полиферментного комплекса.
Дигидролипоат-ацетилтрансфераза (Е2), ее коферментом является липоевая кислота, катализирует 2-ю и 3-ю реакции.

Слайд 8II этап - продолжение
III этап
Дигидролипоат-дегидрогеназа (Е3), кофермент – ФАД, катализирует
, кофермент – ФАД, катализирует 4-ю и 5-ю реакции.")
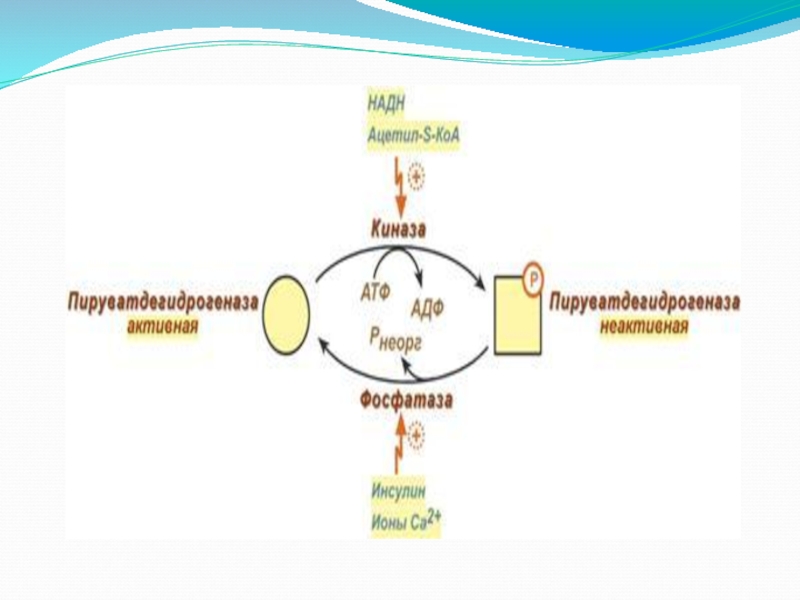
Слайд 9Регуляция пируватдегидрогеназного комплекса
Превращение пирувата в ацетил-КоА – процесс необратимый. Поэтому синтез
ПВК-ДГ комплекс регулируется методом фосфорилирования-дефосфорилирования. Повышение величин отношений НАДН/НАД+, ацетил- КоА/КоА или АТФ/АДФ способствует фосфорилированию фермента протеинкиназой и инактивации комплекса. Следовательно, комплекс инактивируется, если клетка богата энергией и биосинтетическими предшественниками.
Пируват и АДФ, наоборот, активируют ПВК-ДГ комплекс посредством ингибировния протеинкиназы


Слайд 16Реакции лимонного цикла
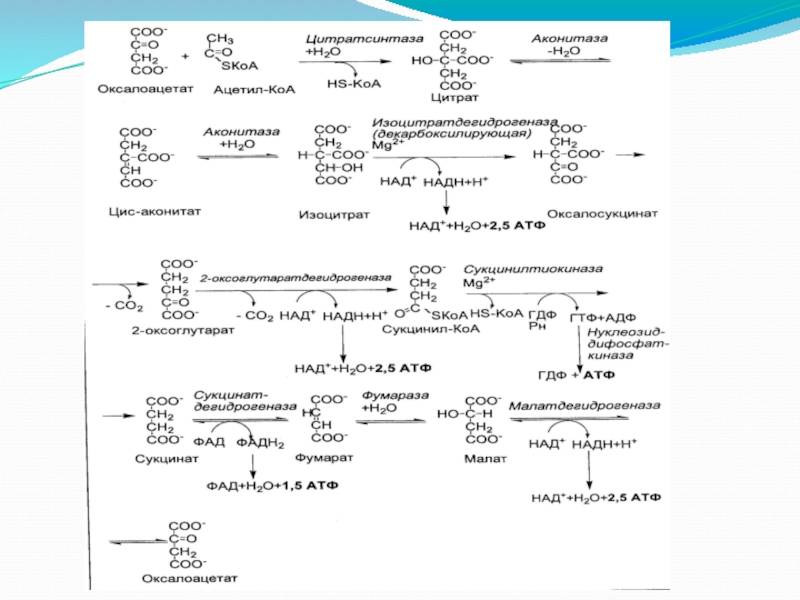
1-я реакция цикла
Свободную уксусную кислоту невозможно окислить путем дегидрирования.

Слайд 17Реакции лимонного цикла
2-я реакция цикла
Аконитат-гидратаза (жонитаза) катализирует превращение цитрата в изоцитрат
 катализирует превращение цитрата в изоцитрат через стадию цис-аконитовой кислоты.")
Слайд 18Реакции лимонного цикла
3-я реакция цикла
Изоцитратдегидрогеназа (деарбоксилирующая) катализирует дегидрирование изолимонной кислоты в
 катализирует дегидрирование изолимонной кислоты в оксалосукцинат (щавелевоянтарную кислоту), которая")
Слайд 19Реакции лимонного цикла
4-я реакция цикла
2-Оксоглутаратдегидрогеназный комплекс (α-кетоглутаратдегидрогепазный комплекс) катализирует окислительное декарбоксилирование
 катализирует окислительное декарбоксилирование 2-оксоглутарата в сукцинил-КоА. Мультифсрментный")
Слайд 20Реакции лимонного цикла
5-я реакция цикла
Сукцинилтиокиназа катализирует расщепление сукцинил-КоА на янтарную кислоту

Слайд 21Реакции лимонного цикла
6-я реакция цикла
Сукцинатдегидрогеназа катализирует превращение сукцината в фумаровую кислоту.

Слайд 22Реакции лимонного цикла
7-я реакция цикла
Фумарат-гидратаза (фумараза) катализирует превращение фумаровой кислоты в
 катализирует превращение фумаровой кислоты в яблочную (малат) с участием")
Слайд 23Реакции лимонного цикла
8-я реакция цикла
Малатдегидрогеназа катализирует окисление яблочной кислоты в оксалоацетат.

Слайд 25Интегративная функция — цикл является связующим звеном между реакциями анаболизма и катаболизма.
Катаболическая
Жирные кислоты, пируват, Лей,Фен — Ацетил-КоА.
Арг, Гис, Глу — α-кетоглутарат.
Фен, тир — фумарат.
Анаболическая функция — использование субстратов цикла на синтез органических веществ:
Оксалацетат — глюкоза, Асп, Асн.
Сукцинил-КоА — синтез гема.
CО2 — реакции карбоксилирования.
Водорододонорная функция — цикл Кребса поставляет на дыхательную цепь митохондрий протоны в виде трех НАДН.Н+ и одного ФАДН2.
Энергетическая функция — 3 НАДН.Н+ дает 7.5 моль АТФ, 1 ФАДН2 дает 1.5 моль АТФ на дыхательной цепи. Кроме того в цикле путем субстратного фосфорилирования синтезируется 1 ГТФ, а затем из него синтезируется АТФ посредствам трансфосфорилирования: ГТФ + АДФ = АТФ + ГДФ.
Функции ЦТК

Слайд 26Анаплеротические реакции
Анаплероmuческuе (пополняющие) реакции – специальные ферментативные реакции, обеспечивающие пополнение пула
Анаплеротические реакции
 реакции – специальные ферментативные реакции, обеспечивающие пополнение пула промежуточных продуктов цикла лимонной")
Слайд 27Ферментативное карбоксилирование пирувата
Наиболее важная анаплеротическая реакция в животных тканях – это

Слайд 28 Зависимое от АТP и биотина карбоксилирование пирувата – анаплеротический путь
Ферментативное карбоксилирование пирувата
Роль биотина в реакции, катализируемой пируваткарбоксилазой

")
Слайд 30 Регуляция цикла Кребса на уровне цитратсинтазы, изоцитратдегидрогеназы и α-КГ-дегидрогеназного комплекса
Общая
Регуляция общего пути катаболизма:
1 – ПВК-ДГ комплекс активируется пируватом, НАД+, HS-КоА; ингибируется НАДН и ацетил~S-KоА;
2 – цитратсинтаза (реакция ускоряется при повышении концентрации оксалоацетата и замедляется при повышении концентрации цитрата, НАДН, АТФ и сукцинил~SKоА);
3 – изоцитратдегидрогеназа аллостерически активируется АДФ, ионами кальция, ингибируется НАДН;
4 – α-кетоглутаратдегидрогеназный комплекс ингибируется НАДН, АТФ
и сукцинил~SКоА, а активируется ионами кальция

Слайд 31Цикл лимонной кислоты – это один из амфиболических путей
Использование метаболитов ЦТК