- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Обмен белков. Источники и пути обезвреживания аммиака. (Лекция 15) презентация
Содержание
- 1. Обмен белков. Источники и пути обезвреживания аммиака. (Лекция 15)
- 2. В процессе эволюции сформировались различные типы азотистого
- 3. Обмен аммиака. Основной источник аммиака - АК.
- 4. Количество азотсодержащих веществ в моче (%) при нормальном белковом питании.
- 5. поступление аммиака из кишечника в портальную вену
- 7. Функции аммиака Аммиак вовлекается (непосредственно или
- 8. Аммиак - токсичное соединение. Даже небольшое повышение
- 9. Недостаточность α-кетоглутарата приводит к снижению концентрации метаболитов
- 10. Обезвреживание аммиака: Основной путь — это связывание
- 11. Пути использования глутамина в организме.
- 12. Амидирование остатков глутаминовой и аспарагиновой кислот в
- 16. Метаболизм амидного азота глутамина в почках Общее обезвреживание аммиака
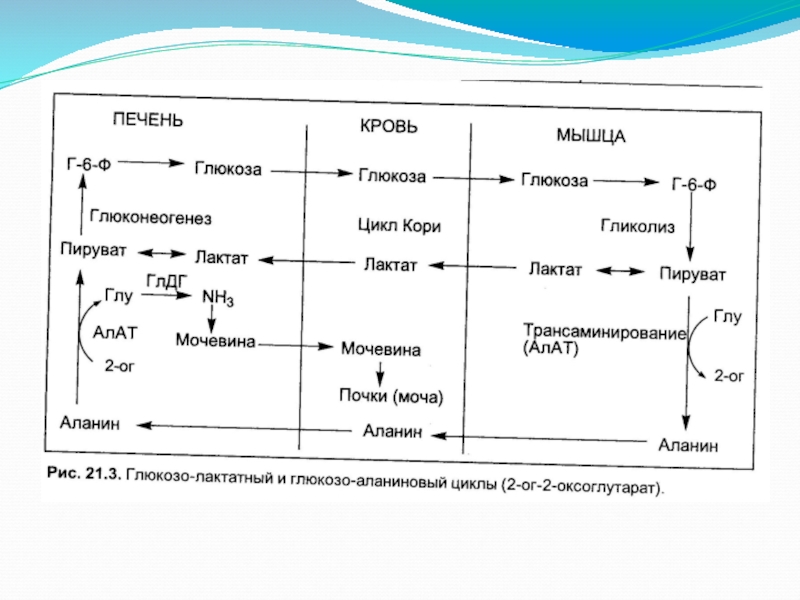
- 17. Глюкозо-аланиновый цикл.
- 18. Орнитиновый цикл Кребса-Гензелейта. Окислительное дезаминирование
- 19. Синтез мочевины
- 20. Цикл регенерации аспартата, сопряжённый с орнитиновым циклом.
- 21. Орнитиновый цикл в печени выполняет 2 функции:
- 22. Взаимосвязь орнитинового цикла и общего пути катаболизма.
- 23. Пути выведения аммиака при включении в диету
- 24. Нарушения синтеза мочевины
- 25. Нарушение синтеза и выведения мочевины
- 26. Количество азотсодержащих веществ в моче (%) при нормальном белковом питании
- 27. Синтез и распад пуриновых и пиримидиновых оснований
- 28. Строение мононуклеотида
- 29. Пуриновые основания нуклеиновых кислот
- 30. Пиримидиновые основания нуклеиновых кислот
- 31. Углеводы нуклеотидов
- 32. Гидролиз нуклеопротеинов
- 33. Катаболизм пуриновых оснований
- 34. Ингибитор ксантиноксидазы
- 35. Распад пиримидиновых оснований
- 36. Синтез инозиновой кислоты
- 37. Происхождение атомов пуринового кольца
- 38. Синтез пуриновых нуклеотидов
- 39. Биосинтез пиримидинов
- 40. Синтез пиримидиновых нуклеотидов
- 41. Синтез дезоксирибонуклеотидов
- 42. Пуриновые основания нуклеиновых кислот
- 43. Пиримидиновые основания нуклеиновых кислот
- 44. Углеводы нуклеотидов
- 45. Строение мононуклеотида
- 46. Комплиментарность азотистых оснований

Слайд 2В процессе эволюции сформировались различные типы азотистого обмена, которые классифицируются по
основному конечному продукту:
1) аммониотелический тип, главный конечный продукт — аммиак (рыбы);
2) урикотелинеский тип, главный конечный продукт — мочевая кислота (рептилии, птицы);
3) уреотелический тип, главный конечный продукт — мочевина (млекопитающие).
1) аммониотелический тип, главный конечный продукт — аммиак (рыбы);
2) урикотелинеский тип, главный конечный продукт — мочевая кислота (рептилии, птицы);
3) уреотелический тип, главный конечный продукт — мочевина (млекопитающие).
")
Слайд 3Обмен аммиака. Основной источник аммиака - АК. Большая часть образовавшегося аммиака
обезвреживается в орнитиновом цикле в печени и выделяется в виде мочевины. Основной реакцией обезвреживания аммиака в тканях является синтез глутамина, который затем используется в анаболических процессах и для обезвреживания веществ в печени. Ферменты глутаматдегидрогеназа и глутаминсинтетаза являются регуляторными и обусловливают скорость процессов образования и обезвреживания аммиака.

 при нормальном белковом питании.")

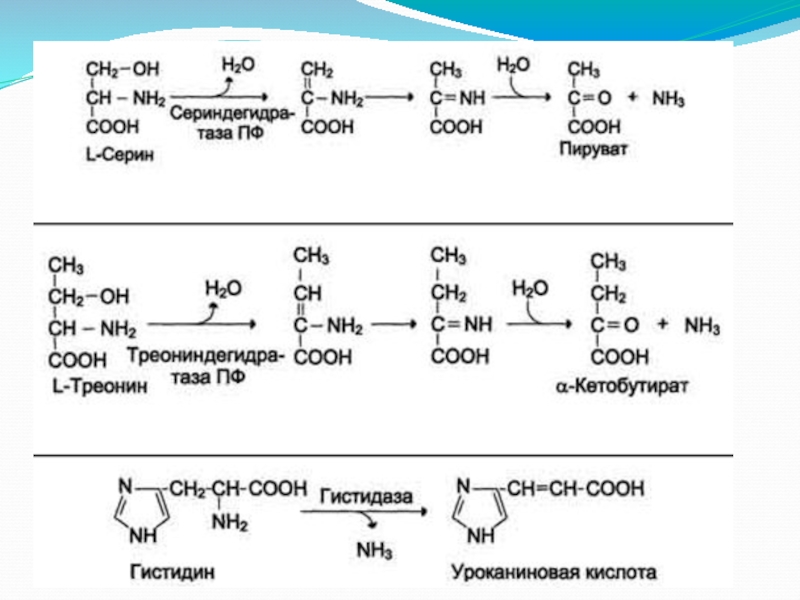
Слайд 7Функции аммиака
Аммиак вовлекается (непосредственно или через глутамин) в синтез многих компонентов
организма: заменимые аминокислоты, пурины, пиримидины, аминосахара и т. д.
 в синтез многих компонентов организма: заменимые аминокислоты, пурины,")
Слайд 8Аммиак - токсичное соединение. Даже небольшое повышение его концентрации оказывает неблагоприятное
действие на организм, и прежде всего на ЦНС. Так, повышение концентрации аммиака в мозге до 0,6 ммоль вызывает судороги. К симптомам гипераммониемии относят тремор, нечленораздельную речь, тошноту, рвоту, головокружение, судорожные припадки, потерю сознания. В тяжёлых случаях развивается кома с летальным исходом.
Механизм токсического действия аммиака на мозг и организм в целом, очевидно, связан с действием его на несколько функциональных систем.
Аммиак легко проникает через мембраны в клетки и в митохондриях сдвигает реакцию, катализируемую глутаматдегидрогеназой, в сторону образования глутамата:
α-Кетоглутарат + NADH + Н+ + NH3 → Глутамат + NAD+.
Уменьшение концентрации α-кетоглутарата вызывает:
угнетение обмена аминокислот (реакции транса-минирования) и, следовательно, синтеза из них нейромедиаторов (ацетилхолина, дофамина и др.);
гипоэнергетическое состояние в результате снижения скорости ЦТК.
Механизм токсического действия аммиака на мозг и организм в целом, очевидно, связан с действием его на несколько функциональных систем.
Аммиак легко проникает через мембраны в клетки и в митохондриях сдвигает реакцию, катализируемую глутаматдегидрогеназой, в сторону образования глутамата:
α-Кетоглутарат + NADH + Н+ + NH3 → Глутамат + NAD+.
Уменьшение концентрации α-кетоглутарата вызывает:
угнетение обмена аминокислот (реакции транса-минирования) и, следовательно, синтеза из них нейромедиаторов (ацетилхолина, дофамина и др.);
гипоэнергетическое состояние в результате снижения скорости ЦТК.

Слайд 9Недостаточность α-кетоглутарата приводит к снижению концентрации метаболитов ЦТК, что вызывает ускорение
реакции синтеза оксалоацетата из пирувата, сопровождающейся интенсивным потреблением СО2. Усиленное образование и потребление диоксида углерода при гипераммониемии особенно характерны для клеток головного мозга.
Повышение концентрации аммиака в крови сдвигает рН в щелочную сторону (вызывает алкалоз). Это, в свою очередь, увеличивает сродство гемоглобина к кислороду, что приводит к гипоксии тканей, накоплению СО2 и гипоэнергетическому состоянию, от которого главным образом страдает головной мозг.
Высокие концентрации аммиака стимулируют синтез глутамина из глутамата в нервной ткани (при участии глутаминсинтетазы):
Глутамат + NH3 + АТФ → Глутамин + АДФ + Н3Р04.
Накопление глутамина в клетках нейроглии приводит к повышению осмотического давления в них, набуханию астроцитов и в больших концентрациях может вызвать отёк мозга. Снижение концентрации глутамата нарушает обмен аминокислот и нейромедиаторов, в частности синтез ϒ-аминомасляной кислоты (ГАМК), основного тормозного медиатора. При недостатке ГАМК и других медиаторов нарушается проведение нервного импульса, возникают судороги.
Ион NH4+ практически не проникает через цитоплазматические и митохондриальные мембраны. Избыток иона аммония в крови способен нарушать трансмембранный перенос одновалентных катионов Na+ и К+, конкурируя с ними за ионные каналы, что также влияет на проведение нервных импульсов.
Повышение концентрации аммиака в крови сдвигает рН в щелочную сторону (вызывает алкалоз). Это, в свою очередь, увеличивает сродство гемоглобина к кислороду, что приводит к гипоксии тканей, накоплению СО2 и гипоэнергетическому состоянию, от которого главным образом страдает головной мозг.
Высокие концентрации аммиака стимулируют синтез глутамина из глутамата в нервной ткани (при участии глутаминсинтетазы):
Глутамат + NH3 + АТФ → Глутамин + АДФ + Н3Р04.
Накопление глутамина в клетках нейроглии приводит к повышению осмотического давления в них, набуханию астроцитов и в больших концентрациях может вызвать отёк мозга. Снижение концентрации глутамата нарушает обмен аминокислот и нейромедиаторов, в частности синтез ϒ-аминомасляной кислоты (ГАМК), основного тормозного медиатора. При недостатке ГАМК и других медиаторов нарушается проведение нервного импульса, возникают судороги.
Ион NH4+ практически не проникает через цитоплазматические и митохондриальные мембраны. Избыток иона аммония в крови способен нарушать трансмембранный перенос одновалентных катионов Na+ и К+, конкурируя с ними за ионные каналы, что также влияет на проведение нервных импульсов.

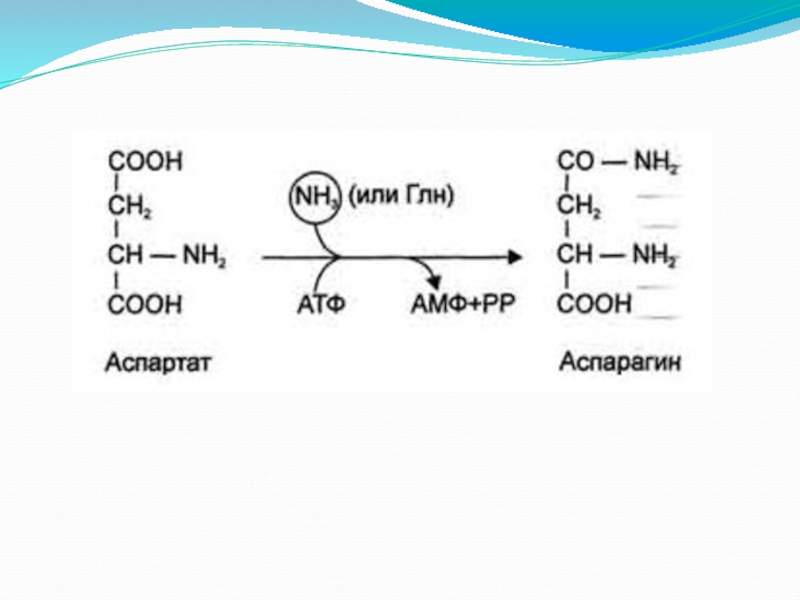
Слайд 10Обезвреживание аммиака:
Основной путь — это связывание аммиака с глутаминовой и реже
аспарагиновой кислотами с образованием соответствующих амидов — глутамина и аспарагина (фермент глутаминсинтетаза). Глутамин является нетоксической транспортной формой аммиака и его концентрация в крови значительно выше концентрации других аминокислот.


Слайд 12Амидирование остатков глутаминовой и аспарагиновой кислот в составе белков.
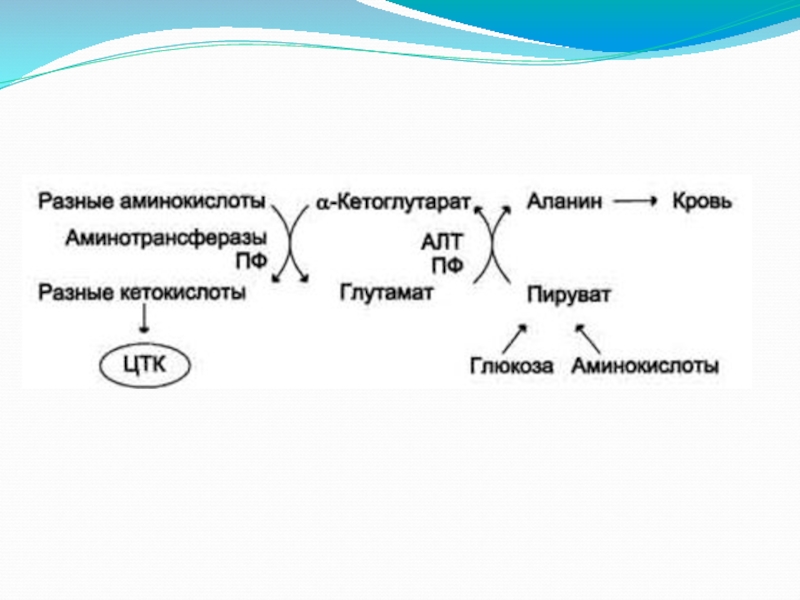
Восстановительное аминирование 2-оксоглутарата
в глутамат. Глутамат в реакциях трансаминирования с пируватом образует аланин (особенно в мышцах).



Слайд 18Орнитиновый цикл Кребса-Гензелейта.
Окислительное дезаминирование глутамата происходит в митохондриях. Ферменты орнитинового
цикла распределены между митохондриями и цитозолем. Поэтому необходим трансмембранный перенос глутамата, цитруллина и орнитина с помощью специфических транслоказ. На схеме показаны пути включения азота двух разных аминокислот (аминокислота 1 и аминокислота 2) в молекулу мочевины: одна аминогруппа - в виде аммиака в матриксе митохондрии; вторую аминогруппу поставляет аспартат цитозоля.



Слайд 21Орнитиновый цикл в печени выполняет 2 функции:
превращение азота аминокислот в
мочевину, которая экскретируется и предотвращает накопление токсичных продуктов, главным образом аммиака;
синтез аргинина и пополнение его фонда в организме.
Регуляторные стадии процесса - синтез карбамоилфосфата, синтез цитруллина и заключительная стадия, катализируемая аргиназой.
Эффективность работы орнитинового цикла при нормальном питании человека и умеренных физических нагрузках составляет примерно 60% его мощности. Запас мощности необходим для избежания гипераммониемии при изменениях количества белка в пище. Увеличение скорости синтеза мочевины происходит при длительной физической работе или длительном голодании, которое сопровождается распадом тканевых белков. Некоторые патологические состояния, характеризующиеся интенсивным распадом белков
синтез аргинина и пополнение его фонда в организме.
Регуляторные стадии процесса - синтез карбамоилфосфата, синтез цитруллина и заключительная стадия, катализируемая аргиназой.
Эффективность работы орнитинового цикла при нормальном питании человека и умеренных физических нагрузках составляет примерно 60% его мощности. Запас мощности необходим для избежания гипераммониемии при изменениях количества белка в пище. Увеличение скорости синтеза мочевины происходит при длительной физической работе или длительном голодании, которое сопровождается распадом тканевых белков. Некоторые патологические состояния, характеризующиеся интенсивным распадом белков

Слайд 22Взаимосвязь орнитинового цикла и общего пути катаболизма. Фумарат, образующийся в результате
расщепления аргининосукцината, превращается в малат, который затем переносится в митохондрии, включается в ЦТК и дегидрируется с образованием оксалоацетата. Эта реакция сопровождается выделением 3 молекул АТФ, которые и компенсируют затраты энергии на синтез одной молекулы мочевины.

Слайд 23Пути выведения аммиака при включении в диету глутамата и фенилацетата (А),
бензоата (Б), цитруллина и аргинина (В). На рисунке обозначены ферментные блоки: 1 - дефект карбамоилфосфатсинтетазы I; 2-дефект орнитинкарбамоилтрансферазы; 3 - дефект аргининосукцинатлиазы.
, бензоата (Б), цитруллина и")


 при нормальном белковом питании")

























