- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Мускулатура рыб. Физиологические основы движения. Лекция 4 презентация
Содержание
- 1. Мускулатура рыб. Физиологические основы движения. Лекция 4
- 2. Механическое движение занимает важное место в
- 3. Плавает рыба благодаря гребным движениям плавников
- 4. Одиночный цикл движения плавника подразделяется на
- 5. Движение грудных плавников у большинства рыб
- 6. Наибольшие скорости достигаются рыбами при периодическом,
- 7. Упор при ундуляционном движении создается искривлением
- 8. скорость рыбы относительно воды можно вычислить,
- 9. Мускулатура это специализированная ткань, основным назначением
- 10. Гладкие мышцы образуют стенки (мышечный слой) внутренних органов и кровеносных сосудов.
- 11. В миофибрилах гладких мышц отсутствует поперечная
- 12. Физиологические свойства гладких мышц значительно отличаются
- 13. Возбуждение в гладких мышцах может передаваться
- 14. Характерными для гладких мышц являются также
- 15. Адекватным раздражителем для гладких мышц является
- 16. Гладкие мышцы инервируются симпатическими и парасимпатическими
- 17. Поперечно-полосатая мускулатура (туловищно-хвостовая) рыб состоит из
- 18. К миосептам присоединяются мышечные волокна, с
- 19. Светлая и темная мускулатура рыб во
- 20. Количество и расположение темных мышц зависит
- 21. Темные мышцы Особенностью этих мышц является обильное
- 22. Светлые мышцы снабжены, в отличие от темных,
- 23. В естественных условиях к мышечным волокнам
- 24. Если раздражающие импульсы сближены так, что
- 25. Зубчатый тетанус возникает
- 26. Исследования мышечного волокна сегодня проводятся на
- 29. На миозиновых нитях есть поперечные выступы
- 30. Главным в теории является то, что
- 31. Этапы сокращения Механизм мышечного сокращения кратко
- 32. Для лучшего понимания процесса активации волокна
- 33. Когда мышца расслаблена, в ней на

Слайд 2Механическое движение
занимает важное место в жизнедеятельности всех без исключения живых
организмов, в том числе и рыб.

Слайд 3
Плавает рыба благодаря гребным движениям плавников и волнообразным движениям тела (ундуляция).
Рыбы перемещаются в воде, создавая упор, т.е. отталкиваясь от среды, обладающей некоторой плотностью, вязкостью и податливостью. Это особенно наглядно видно на примере работы грудных плавников.
При движении плавника назад во время гребка, он максимально расправлен и создает максимальный упор. При движении в обратном направлении, т.е. вперед лучи плавника сведены и создают минимальный тормозящий упор.
При движении плавника назад во время гребка, он максимально расправлен и создает максимальный упор. При движении в обратном направлении, т.е. вперед лучи плавника сведены и создают минимальный тормозящий упор.
. Рыбы перемещаются в воде,")
Слайд 4Одиночный цикл движения плавника
подразделяется на 3 фазы:
1) движение вперед,
т.е. подготовка гребка;
2) движение назад (гребок)
3) плавник прижат к телу (рефрактерная фаза).
2) движение назад (гребок)
3) плавник прижат к телу (рефрактерная фаза).
 движение вперед, т.е. подготовка гребка; 2)")
Слайд 5
Движение грудных плавников у большинства рыб поочередное (щука, колюшка, судак), но
у некоторых видов рыб встречается одновременное (сазан, линь, карась и др.).
Скорость плавания при работе одних только плавников незначительна, расход энергии тоже низкий, т.к. в работу включены небольшие участки мускулатуры, в частности мышцы грудного пояса.
С помощью плавников рыбы могут перемещаться, как правило, скоростью 1-2 длины тела в секунду (L/c). Коэффициент полезного действия при таком типе плавания достаточно высок.
При таком типе плавания, т.е. в котором участвуют как двигатель плавники, туловищно-хвостовая мускулатура напряжена и поддерживает тело в определенной позе.
Скорость плавания при работе одних только плавников незначительна, расход энергии тоже низкий, т.к. в работу включены небольшие участки мускулатуры, в частности мышцы грудного пояса.
С помощью плавников рыбы могут перемещаться, как правило, скоростью 1-2 длины тела в секунду (L/c). Коэффициент полезного действия при таком типе плавания достаточно высок.
При таком типе плавания, т.е. в котором участвуют как двигатель плавники, туловищно-хвостовая мускулатура напряжена и поддерживает тело в определенной позе.
, но у некоторых видов рыб")
Слайд 6Наибольшие скорости
достигаются рыбами при периодическом, волнообразном (движении) изгибании тела (ундуляция).
В данном случае движителем служит вся поверхность тела, за исключением головы. В процессе движения, таким образом, напрягается вся мускулатура туловища и хвоста, которая составляет около 50%, т.е. половину массы тела.
 изгибании тела (ундуляция). В данном случае движителем")
Слайд 7
Упор при ундуляционном движении создается искривлением тела и движением локомоторной (волнообразующие
движения) волны от головы к хвосту.
 волны от головы")
Слайд 8скорость рыбы
относительно воды можно вычислить, определив ее длину и частоту
ударов хвоста, т.е. чем выше частота ундуляций (т.е. частота ударов хвоста), тем выше скорость рыбы относительно воды. Максимальная частота ундуляций у рыб может достигать 30 Гц, а скорость движения до 22,5 длины тела в секунду, т.е. 40 м/сек или 150 км/ч.
Наилучшими пловцами являются пелагические рыбы: меч-рыба, рыба-пила, тунцы, ставриды, скумбрии и др. Неважные пловцы бентофаги, такие как осетровые, угри, камбала и др.
Наилучшими пловцами являются пелагические рыбы: меч-рыба, рыба-пила, тунцы, ставриды, скумбрии и др. Неважные пловцы бентофаги, такие как осетровые, угри, камбала и др.

Слайд 9Мускулатура это специализированная ткань,
основным назначением которой в организме является сокращение,
лежащее в основе бесконечно разнообразной двигательной деятельности. Мышечная ткань животных, в том числе и рыб, подразделяется на два основных типа:
гладкие мышцы,
поперечно-полосатые мышцы.
гладкие мышцы,
поперечно-полосатые мышцы.

 внутренних органов и кровеносных сосудов.")
Слайд 11
В миофибрилах гладких мышц отсутствует поперечная исчерченность. Это обусловлено хаотическим расположением
сократительных белков в волокнах гладких мышц. Волокна гладких мышц относительно короче поперечно-полосатых и достигают длины от 50 до 200 мкм.

Слайд 12
Физиологические свойства гладких мышц значительно отличаются от физиологических свойств поперечно-полосатых мышц.
Гладкие мышцы менее возбудимы, чем поперечно-полосатые. Возбуждение по ним распространяется с относительно небольшой скоростью 2-15см/сек.

Слайд 13
Возбуждение в гладких мышцах может передаваться с одного волокна на другое,
что отсутствует у поперечно-полосатых мышц. Сокращение гладкой мускулатуры происходит более медленно и длительно.

Слайд 14Характерными для гладких мышц
являются также длительные тонические сокращения.
Рефрактерный период
в гладких мышцах более продолжителен, чем в скелетных (до нескольких секунд).
Важным свойством гладкой мышцы является ее высокая пластичность, то есть способность сохранять приданную растяжением длину без изменения напряжения. Мочевой, плавательной, желчный пузыри способные значительно растягиваться.
Характерной особенностью гладких мышц является способность к автоматической деятельности, которая обеспечивается нервными элементами, заложенными в стенках гладкомышечных органов..
Важным свойством гладкой мышцы является ее высокая пластичность, то есть способность сохранять приданную растяжением длину без изменения напряжения. Мочевой, плавательной, желчный пузыри способные значительно растягиваться.
Характерной особенностью гладких мышц является способность к автоматической деятельности, которая обеспечивается нервными элементами, заложенными в стенках гладкомышечных органов..

Слайд 15
Адекватным раздражителем для гладких мышц является их быстрое и сильное растяжение,
что имеет большое значение для функционирования многих гладкомышечных полых органов (желудочно-кишечный тракт, сосуды и т.д.).
Особенностью гладких мышц является также их высокая чувствительность к некоторым биологически активным веществам (ацетилхолин, адреналин, серотонин и др.).
Особенностью гладких мышц является также их высокая чувствительность к некоторым биологически активным веществам (ацетилхолин, адреналин, серотонин и др.).

Слайд 16
Гладкие мышцы инервируются симпатическими и парасимпатическими вегетативными нервами, которые, как правило,
оказывают противоположное влияние на их функциональное состояние.
Так, например, сигналы, поступающие по симпатическим нервам, способствуют сокращению мускулатуры желудочно-кишечного тракта, а сигналы идущие по парасимпатическим нервам , наоборот способствуют расслаблению.
Так, например, сигналы, поступающие по симпатическим нервам, способствуют сокращению мускулатуры желудочно-кишечного тракта, а сигналы идущие по парасимпатическим нервам , наоборот способствуют расслаблению.

Слайд 17
Поперечно-полосатая мускулатура (туловищно-хвостовая) рыб состоит из миомеров (сегментов), число которых равно
числу позвонков. Число миомеров варьирует от 20 (бычки) до 100 (угорь) и зависит от видовой принадлежности. Поперечно-полосатая мускулатура представлена белыми и красными мышцами. Соединительно-тканные элементы, отделяющие миомеры, называются миосептами.
 рыб состоит из миомеров (сегментов), число которых равно числу позвонков. Число миомеров")
Слайд 18
К миосептам присоединяются мышечные волокна, с помощью которых сокращение мускулатуры передается
костному скелету, что вызывает локомоторные волнообразные движения тела. Миомеры имеют сложную геометрическую форму и представляют собой как бы серию вложенных один в другой конусов. Мышечные волокна в миомерах ориентированы таким образом, что одно волокно является, как бы продолжением другого и такие составные нити косо направлены от средней линии к хвосту и краям тела.

Слайд 19
Светлая и темная мускулатура рыб во многом различны, как по строению
и биохимическому составу, так и по функциям. Число сокращений волокон темной мускулатуры равно числу плавательных движений, а сокращения белой мускулатуры не регулярны.
Белая мускулатура поддерживает ритмическую работу темных мышц энергичными мощными движениями, которые производятся с интервалом примерно и 1 секунду (секунду энергично сокращается, а затем секунду отдыхает).
Белая мускулатура поддерживает ритмическую работу темных мышц энергичными мощными движениями, которые производятся с интервалом примерно и 1 секунду (секунду энергично сокращается, а затем секунду отдыхает).

Слайд 20
Количество и расположение темных мышц зависит от видовой принадлежности и образа
жизни рыб, так у пелагических рыб (тунца, макрели, ставриды, сельди) доля темной мускулатуры значительная и достигает 20% от всей мышечной массы, а у малоподвижных рыб (скорпены, судак, бычки, окунь, щука, камбала, осетровые и. т.д.) темная мускулатура слабо развита и присутствует только в области плавников. Цвет темных мышц обусловлен наличием и них геминовых пигментов миоглобина и цитохромов. Основным энергетическим веществом являются липиды (жирные кислоты, глицерин и продукты их распада).

Слайд 21Темные мышцы
Особенностью этих мышц является обильное снабжение их кровеносными сосудами. В
темных мышцах даже при интенсивной работе анаэробные процессы (т.е. окислительные процессы, идущие без кислорода) компенсируются аэробными (т.е. процессами, протекающими при обязательном наличии кислорода).
Темные мышцы относятся к мышцам тонического типа (поддерживают тонус мышечной массы). Они обильно иннервированы нервными волокнами.
Темные мышцы относятся к мышцам тонического типа (поддерживают тонус мышечной массы). Они обильно иннервированы нервными волокнами.

Слайд 22Светлые мышцы
снабжены, в отличие от темных, значительно меньшим количеством кровеносных сосудов,
а мощные анаэробные процессы при их сокращении не компенсируются аэробными.
Основным энергетическим веществом («топливом») в них являются углеводы. Белые мышцы относятся к типу фазических или быстрых, их мощность и скорость сокращения в 2 раза больше, чем у темных.
Иннервация белой мускулатуры у костистых рыб также как и красной мускулатуры очень обильная.
Сократительные свойства белых мышц у различных рыб разнообразны: у одних рыб движения плавны и равномерны, у других - резкие с быстрым ускорением. В соответствии с этим находятся и свойства мышечных волокон.
Основным энергетическим веществом («топливом») в них являются углеводы. Белые мышцы относятся к типу фазических или быстрых, их мощность и скорость сокращения в 2 раза больше, чем у темных.
Иннервация белой мускулатуры у костистых рыб также как и красной мускулатуры очень обильная.
Сократительные свойства белых мышц у различных рыб разнообразны: у одних рыб движения плавны и равномерны, у других - резкие с быстрым ускорением. В соответствии с этим находятся и свойства мышечных волокон.

Слайд 23
В естественных условиях к мышечным волокнам поступают не одиночные, а ряд
нервных импульсов, на которые мышцы отвечают длительным сокращением, которое получило название тетанического сокращения или тетануса. К тетаническому сокращению способны только поперечно полосатые мышцы. Гладкие мышцы не способны к тетаническому сокращению вследствие наличия продолжительного рефрактерного периода.
Рефрактерность это временное снижение возбудимости ткани, возникающее после ее возбуждения. Ели раздражающие импульсы сближены и каждый из них приходится на тот момент, когда мышца только начала расслабляться, то возникает зубчатый тип сокращения, который получил название неполного, несовершенного, зубчатого, тетануса (клонус).
Рефрактерность это временное снижение возбудимости ткани, возникающее после ее возбуждения. Ели раздражающие импульсы сближены и каждый из них приходится на тот момент, когда мышца только начала расслабляться, то возникает зубчатый тип сокращения, который получил название неполного, несовершенного, зубчатого, тетануса (клонус).

Слайд 24
Если раздражающие импульсы сближены так, что каждый последующий импульс приходится на
время, когда мышца не успела перейти к расслаблению от предыдущего раздражения, т.е. происходит на высоте ее сокращения, то возникает длительное непрерывное сокращение, получившее название гладкого совершенного тетануса. Свершенный тетанус нормальное рабочее состояние поперечно-полосатой мышцы обуславливается поступлением из ЦНС (центральной нервной системы) нервных импульсов с частотой 40-50 импульсов/секунду.

Слайд 25
Зубчатый тетанус возникает при частоте нервных
импульсов до 30 импульсов/секунду. Если мышца получает 10-20 нервных импульсов/секунду, то она находится в состоянии мышечного тонуса, т.е. умеренной степени напряжения . Тетаническое сокращение имеет следующие преимущества перед одиночным сокращением. Оно сильнее и продолжительнее, что дает возможность сохранить определенное положение тела.
Следует отметить, что у рыб встречается и зубчатый тетанус в естественных условиях. Например, конвульсивные, тетанические сокращения тела при брачных играх, во время икрометания и выделения молок.
Следует отметить, что у рыб встречается и зубчатый тетанус в естественных условиях. Например, конвульсивные, тетанические сокращения тела при брачных играх, во время икрометания и выделения молок.

Слайд 26
Исследования мышечного волокна сегодня проводятся на основе рентгеноструктурного анализа, электронной микроскопии,
а также гистохимическими методами. Рассчитано, что в каждую миофибриллу, диаметр которой составляет 1 мкм, входит примерно 2500 протофибрилл, то есть удлиненных полимеризованных молекул белков (актина и миозина).
Актиновые протофибриллы в два раза тоньше миозиновых. В покое эти мышцы находятся так, что актиновые нити кончиками проникают в промежутки между миозиновыми протофибриллами. Узкая светлая полоса в диске А свободна от актиновых нитей. А мембрана Z скрепляет их. -
Актиновые протофибриллы в два раза тоньше миозиновых. В покое эти мышцы находятся так, что актиновые нити кончиками проникают в промежутки между миозиновыми протофибриллами. Узкая светлая полоса в диске А свободна от актиновых нитей. А мембрана Z скрепляет их. -


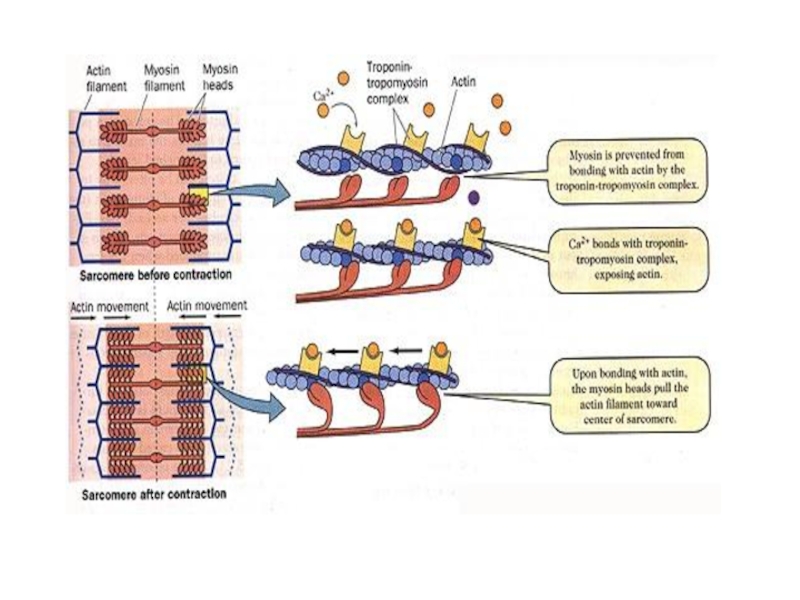
Слайд 29
На миозиновых нитях есть поперечные выступы длиной до 20 нм, в
головках которых находится порядка 150 молекул миозина. Они отходят биополярно, и каждая головка соединяет миозиновую с актиновой нитью. Когда происходит усилие актиновых центров на нитях миозина, актиновая нить приближается к центру саркомера. В конце миозиновые нити доходят до линии Z. Тогда они занимают собой весь саркомер, а актиновые находятся между ними. При этом длина диска I сокращается, а в конце он исчезает полностью, вместе с чем линия Z становится толще. -

Слайд 30
Главным в теории является то, что не нити (миозиновые и актиновые)
укорачиваются. Длина их остается неизменной и при растяжении мышц. Но пучки тонких нитей, проскальзывая, выходят между толстыми нитями, уменьшается степень их перекрытия, таким образом происходит сокращение. Молекулярный механизм мышечного сокращения посредством скольжения актиновых нитей заключается в следующем. Миозиновые головки соединяют протофибриллу с актиновой. При их наклонах происходит скольжение, двигающее актиновую нить к центру саркомера. За счет биполярной организации миозиновых молекул на обеих сторонах нитей создаются условия для скольжения актиновых нитей в разные стороны. При расслаблении мышц миозиновая головка отходит от актиновых нитей. Благодаря легкому скольжению расслабленные мышцы растяжению сопротивляются гораздо меньше. Поэтому они пассивно удлиняются. -
 укорачиваются. Длина их остается")
Слайд 31
Этапы сокращения Механизм мышечного сокращения кратко можно подразделить на следующие этапы:
Мышечное волокно стимулируется, когда потенциал действия поступает от мотонейронов из синапсов. Потенциал действия создается на мембране мышечного волокна, а затем распространяется к миофибриллам. Совершается электромеханическое сопряжение, представляющее собой преобразование электрического ПД в механическое скольжение. В этом обязательно участвуют ионы кальция. -

Слайд 32
Для лучшего понимания процесса активации волокна ионами кальция удобно рассмотреть структуру
актиновой нити. Длина ее составляет порядка 1 мкм, толщина — от 5 до 7 нм. Это пара закрученных ниток, которые напоминают мономер актина. Примерно через каждые 40 нм здесь находятся сферические тропониновые молекулы, а между цепями — тропомиозиновые. Когда ионы кальция отсутствуют, то есть миофибриллы расслабляются, длинные тропомиозиновые молекулы блокируют крепление актиновых цепей и мостиков миозина. Но при активизации ионов кальция тропомиозиновые молекулы опускаются глубже, и участки открываются. Тогда миозиновые мостики прикрепляются к актиновым нитям, а АТФ расщепляется, и сила мышц развивается. Это становится возможным за счет воздействия кальция на тропонин. При этом молекула последнего деформируется, проталкивая тем самым тропомиозин

Слайд 33
Когда мышца расслаблена, в ней на 1 грамм сырого веса содержится
больше 1 мкмоль кальция. Соли кальция изолированы и находятся в особых хранилищах. В противном случае мышцы бы все время сокращались. Хранение кальция происходит следующим образом. На разных участках мембраны клетки мышцы внутри волокна имеются трубки, через которые происходит соединение со средой вне клеток. Это система поперечных трубочек. А перпендикулярно ей находится система продольных, на концах которых — пузырьки (терминальные цистерны), расположенные в непосредственной близости к мембранам поперечной системы. Вместе получается триада. Именно в пузырьках хранится кальций. Так ПД распространяется внутрь клетки, и происходит электромеханическое сопряжение. Возбуждение проникает в волокно, переходит в продольную систему, высвобождает кальций. Таким образом осуществляется механизм сокращения мышечного волокна.