- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Инженерная энзимология. Механизмы инактивации ферментов презентация
Содержание
- 1. Инженерная энзимология. Механизмы инактивации ферментов
- 2. Механизмы инактивации ферментов 1. Изменение первичной структуры:
- 3. Решение: В литературе практически отсутствуют примеры удачной реактивации подобным образом инактивированных ферментов.
- 4. 1.2.Окисление функциональных групп фермента SH-группы цистеина
- 5. Решение: Реактивировать с помощью восстанавливающих агентов, в частности низкомолекулярных тиолов (например, цистеин или дитиотрейтол) .
- 6. 1.3. Расщепление дисульфидных связей Вызывают : Тиолы
- 7. 1.3. Расщепление дисульфидных связей Щелочной гидролиз цистеина
- 8. Решение: Добавление в среду тиолов приведет к
- 9. 1.4. Химическая модификация каталитических SH-групп. Катионы
- 10. 1.5. Фосфорилирование белков in vivo. Под
- 11. Решение: В литературе практически отсутствуют примеры удачной реактивации подобным образом инактивированных ферментов.
- 12. 1.6. Дезаминирование остатков аспарагина. При
- 13. Решение: В литературе практически отсутствуют примеры удачной реактивации подобным образом инактивированных ферментов.
- 14. 1.7. Радиационная инактивация ферментов γ-облучение и УФ-свет
- 15. 2. Агрегация Наблюдается при повышенной температуре, при
- 16. Решение: Необходимо разрушить межмолекулярные ковалентные и нековалентные
- 17. 3. Инактивация ферментов поверхностным натяжением Поверхностное натяжение
- 18. Решение: Добавление ПАВ снижает поверхностное натяжение до 1 дин/см.
- 19. 4. Сорбция белка на стенках реакционного сосуда
- 20. Решение: Десорбция фермента со стенок реакционного сосуда
- 21. 5. Диссоциация олигомерных белков на субъединицы Вызывают:
- 22. 6. Десорбция кофактора из активного центра фермента
- 23. Регенерация кофакторов Способы регенерации:
- 24. Ферментативный способ Использование сопряженных субстратов. В систему
- 25. Ферментативный способ Использование сопряженных субстратов. Недостатка: •
- 26. Ферментативный способ 2. Использование сопряженных ферментативных реакций
- 27. Неферментативные способы Химические методы. Используются дитионит
- 28. Неферментативные способы 2. Электрохимические методы. Прямое электрохимическое
- 29. СТАБИЛИЗАЦИЯ ФЕРМЕНТОВ В БИОТЕХНОЛОГИЧЕСКИХ СИСТЕМАХ
- 30. Проблемы возникающие при использовании ферментов в биотехнологических
- 31. Основные подходы для стабилизации ферментов :
- 32. Стабилизация ферментов с помощью: Субстратов или
- 33. Стабилизация ферментов с помощью: 2. Органических
- 34. Стабилизация ферментов с помощью: 3. Солей: При низких концентрациях солей (
- 36. Химическая модификация фермента 1. Фермент принимает более
- 37. Иммобилизация фермента позволяет: Повысить устойчивость фермента (нагреванию,
- 38. Иммобилизованные ферменты – это препараты ферментов, молекулы
- 39. Методы иммобилизации: В качестве носителей могут применяться:
- 40. Методы физической иммобилизации: 1) адсорбция фермента на
- 41. Методы физической иммобилизации: 1)адсорбция фермента на нерастворимом
- 42. Методы физической иммобилизации: 1)адсорбция фермента на нерастворимом
- 43. Методы физической иммобилизации: 1)адсорбция фермента на нерастворимом
- 44. Методы физической иммобилизации: 2) включение фермента в полупроницаемую капсулу, в полупроницаемую мембрану
- 45. Методы физической иммобилизации: 2) включение фермента в
- 46. Методы физической иммобилизации: 3) механическое включение фермента
- 47. Методы физической иммобилизации: 3) механическое включение фермента
- 48. Методы физической иммобилизации: 3) механическое включение фермента
- 49. Методы физической иммобилизации: 4) Включение в двухфазную
- 50. Методы химической иммобилизации: Образовании ковалентных связей между
- 51. При иммобилизации ферментов необходимо соблюдать следующие условия:
- 52. Весьма перспективным является использование в качестве
- 53. Термозимы Стабильны в условиях высокой температуры, высоких
- 54. Механизмы ответственны за термоустойчивость ферментов у термозимов:
- 55. Было обнаружено, что дегидрогеназа из Pyrococcus furiosus
- 56. Возросшая термостабильность коррелирует: с увеличением жесткости белковой
- 57. Механизмы стабилизации: • минимизация доступной площади гидрофобной
- 58. Применение ферментов из экстремофилов Современные технологии
- 59. Применение ферментов из экстремофилов: Крахмал используется для
- 60. Применение ферментов из экстремофилов: Наиболее термостабильные α-амилазы
- 61. Применение ферментов из экстремофилов: Протеолитические ферменты Сериновые
- 62. Применение ферментов из экстремофилов: ДНК-полимеразы Термостабильные ДНК-полимеразы
- 63. Благодарю за внимание!

Слайд 2Механизмы инактивации ферментов
1. Изменение первичной структуры:
1.1. Разрыв полипептидной цепи:
Жесткие условия (длительное
кипячение в HCl) – гидролиз до отдельных ам-т.
При нагревании до 100 °С (pH 7-8), гидролиз пептидных связей незначителен.
Наиболее чувствительными к высокотемпературному гидролизу являются пептидные связи, образованные остатками аспарагиновой кислоты.
Протеазы (бактериальное загрязнение, автолиз).
При нагревании до 100 °С (pH 7-8), гидролиз пептидных связей незначителен.
Наиболее чувствительными к высокотемпературному гидролизу являются пептидные связи, образованные остатками аспарагиновой кислоты.
Протеазы (бактериальное загрязнение, автолиз).
 –")
Слайд 3Решение:
В литературе практически отсутствуют примеры удачной реактивации подобным образом инактивированных ферментов.

Слайд 41.2.Окисление функциональных групп фермента
SH-группы цистеина и индольные фрагменты триптофана, при повышенной
температуре, могут окисляться (сульфокси- соединения цистеина (SOH, SO2H) и образовываться продукты раскрытия индольного кольца триптофана.

Слайд 5Решение:
Реактивировать с помощью восстанавливающих агентов, в частности низкомолекулярных тиолов (например, цистеин
или дитиотрейтол) .
 .")
Слайд 61.3. Расщепление дисульфидных связей
Вызывают : Тиолы и другие восстановленные соединений серы,
например Na2SO3, Na2S2O3.
Продуктом восстановления дисульфидной связи (S-S) является:
1) тиольная форма (белок–SH)
2) смешанный дисульфид тиольной формы белка с восстанавливающим реагентом, например белок–S–SO3).
Продуктом восстановления дисульфидной связи (S-S) является:
1) тиольная форма (белок–SH)
2) смешанный дисульфид тиольной формы белка с восстанавливающим реагентом, например белок–S–SO3).

Слайд 71.3. Расщепление дисульфидных связей
Щелочной гидролиз цистеина →дегидроаланин→ Благодаря нуклеофильным свойствам взаимодействует
с NH2-группами лизина и SH- цистеина →лизиноаланин и лантионин.
Для полной деструкции всех S–S связей требу-
ются достаточно жесткие условия (0,1–1М щелочь, 100 °С).
Однако деструкция наиболее реакционноспособных S–S связей может проходить в достаточно мягких условиях – например, при температурах 60–80 °С и слабощелочных значениях рН.
Cледует учитывать при использовании ферментов в качестве добавок к моющим средствам.
Для полной деструкции всех S–S связей требу-
ются достаточно жесткие условия (0,1–1М щелочь, 100 °С).
Однако деструкция наиболее реакционноспособных S–S связей может проходить в достаточно мягких условиях – например, при температурах 60–80 °С и слабощелочных значениях рН.
Cледует учитывать при использовании ферментов в качестве добавок к моющим средствам.

Слайд 8Решение:
Добавление в среду тиолов приведет к расщеплению смешанного дисульфида и последующему
образованию правильной S–S связи

Слайд 91.4. Химическая модификация каталитических SH-групп.
Катионы тяжелых металлов (Hg, Pb и
Cu) связываются с SH-групп активного центра фермента
↓ Образование соответствующих меркаптидов
↓
Фермент инактивируется
↓ Образование соответствующих меркаптидов
↓
Фермент инактивируется
 связываются с SH-групп")
Слайд 101.5. Фосфорилирование белков in vivo.
Под действием фосфорилазы и фосфатазы, содержащихся
в полуочищенных ферментативных препаратах в виде примесей
↓
Фосфорная кислота связывается с ОН-группами серина и треонина.
↓
Конформационные изменения в белковой молекуле
↓
инактивацию фермента.
↓
Фосфорная кислота связывается с ОН-группами серина и треонина.
↓
Конформационные изменения в белковой молекуле
↓
инактивацию фермента.

Слайд 11Решение:
В литературе практически отсутствуют примеры удачной реактивации подобным образом инактивированных ферментов.

Слайд 121.6. Дезаминирование остатков аспарагина.
При температурах (порядка 100 °С) и рН
(порядка 4,0–5,0) происходит дезаминирование остатков аспарагина.
↓
инактивации фермента.
↓
инактивации фермента.
 и рН (порядка 4,0–5,0) происходит дезаминирование")
Слайд 13Решение:
В литературе практически отсутствуют примеры удачной реактивации подобным образом инактивированных ферментов.

Слайд 141.7. Радиационная инактивация ферментов
γ-облучение и УФ-свет
Воздействуют на функциональные группы ферментов, пептидные
связи и SH-группы остатков цистеина.

Слайд 152. Агрегация
Наблюдается при повышенной температуре, при экстремальных значениях рН, в присутствии
некоторых химических соединений.
Чем выше концентрация, тем быстрее идет агрегация.
Гидрофобные взаимодействия и водородные связи, возможно образование дисульфидных мостиков между отдельными белковыми молекулами
Чем выше концентрация, тем быстрее идет агрегация.
Гидрофобные взаимодействия и водородные связи, возможно образование дисульфидных мостиков между отдельными белковыми молекулами

Слайд 16Решение:
Необходимо разрушить межмолекулярные ковалентные и нековалентные контакты c помощью концентрированных растворов
мочевины и гуанидинхлорида, экстремальных значений рН.
Если при агрегации ферментов образовались межмолекулярные S-S -мостики, в среду вносят в относительно невысоких концентрациях (мкмоль/л) тиолсодержащие реагенты (например, цистеин или дитиотрейтол).
При таких концентрациях внутримолекулярные S–S связи в белке, как правило, не затрагиваются.
Если при агрегации ферментов образовались межмолекулярные S-S -мостики, в среду вносят в относительно невысоких концентрациях (мкмоль/л) тиолсодержащие реагенты (например, цистеин или дитиотрейтол).
При таких концентрациях внутримолекулярные S–S связи в белке, как правило, не затрагиваются.

Слайд 173. Инактивация ферментов поверхностным натяжением
Поверхностное натяжение на границе раздела между воздухом
и чистой водой составляет 80 дин/см.
Пенообразование вызывает денатурацию ферментов, адсорбированных на границе раздела фаз.
Пенообразование вызывает денатурацию ферментов, адсорбированных на границе раздела фаз.


Слайд 194. Сорбция белка на стенках реакционного сосуда
Сорбция за счет нековалентных взаимодействий
приводит к уменьшению концентрации фермента в растворе.
Необходимо учитывать при работе с разбавленными белковыми растворами (концентрация 10-8–10-10 моль/л).
Под влиянием денатурирующих факторов способность белков сорбироваться на стенках реакционного сосуда может возрастать.
Необходимо учитывать при работе с разбавленными белковыми растворами (концентрация 10-8–10-10 моль/л).
Под влиянием денатурирующих факторов способность белков сорбироваться на стенках реакционного сосуда может возрастать.

Слайд 20Решение:
Десорбция фермента со стенок реакционного сосуда достигается за счет разрушения неспецифических
взаимодействий между
белком и сорбционными центрами на поверхности сосуда.
Можно использовать экстремальные значения рН, концентрированные растворы мочевины или гуанидинхлорида.
белком и сорбционными центрами на поверхности сосуда.
Можно использовать экстремальные значения рН, концентрированные растворы мочевины или гуанидинхлорида.

Слайд 215. Диссоциация олигомерных белков на субъединицы
Вызывают: Мочевина, детергенты, кислоты или же
нагревание.
Приводят к:
конформационным изменениям отдельных субъединиц;
агрегации субъединиц;
диссоциации кофакторов из активных центров;
модификации функциональных групп, которые в олигомерном белке были экранированы от контакта с растворителем.
Приводят к:
конформационным изменениям отдельных субъединиц;
агрегации субъединиц;
диссоциации кофакторов из активных центров;
модификации функциональных групп, которые в олигомерном белке были экранированы от контакта с растворителем.

Слайд 226. Десорбция кофактора из активного центра фермента
Вызывает: нагревание, действие хелаторов, диализ
Если
диссоциация кофактора сопровождается значительными конформационными сдвигами или химической модификацией важных
функциональных групп → фермент инактивируется необратимо.
Если при этом не произошло существенного изменения белковой конформации, то добавление в среду избытка кофактора приводит к реактивации фермента.
функциональных групп → фермент инактивируется необратимо.
Если при этом не произошло существенного изменения белковой конформации, то добавление в среду избытка кофактора приводит к реактивации фермента.

Слайд 23Регенерация кофакторов
Способы регенерации:
Ферментативный (методы с использованием со-
пряженных субстратов или ферментов)
Неферментативный
(химические и электрохимические подходы)
Неферментативный (химические и электрохимические подходы)")
Слайд 24Ферментативный способ
Использование сопряженных субстратов.
В систему вводят избыточное количество сопряженного субстрата того
же фермента:
Пример: при работе алкогольдегидрогеназы расходуется НАДН.
Пример: при работе алкогольдегидрогеназы расходуется НАДН.

Слайд 25Ферментативный способ
Использование сопряженных субстратов.
Недостатка:
• используются высокие концентрации сопряженного субстрата, так
как равновесие реакции сильно сдвинуто в сторону образования спирта;
усложняет процедуру выделения основного продукта из реакционной смеси.
усложняет процедуру выделения основного продукта из реакционной смеси.

Слайд 26Ферментативный способ
2. Использование сопряженных ферментативных реакций
В систему, дополнительно вводят фермент 2,
функционирование которого обеспечивает регенерацию кофермента.
Используемые в системе ферменты должны иметь разную субстратную специфичность
Используемые в системе ферменты должны иметь разную субстратную специфичность

Слайд 27Неферментативные способы
Химические методы.
Используются дитионит натрия и некоторые соли пиридиния:
+ Низкая
стоимость.
могут ингибировать отдельные ферменты.
Флавиновые коферменты
могут ингибировать отдельные ферменты.
Флавиновые коферменты

Слайд 28Неферментативные способы
2. Электрохимические методы.
Прямое электрохимическое восстановление или
окисление.
“-” появления в процессе
регенерации ферментативно неактивных форм кофермента, например в результате его димеризации.


Слайд 30Проблемы возникающие при использовании ферментов в биотехнологических процессах:
Повышенные температуры
Экстремальные значения pH
Высокие концентрации органических растворителей или ПАВ.
Невозможность многократно использовать фермент.
Сложность при разделении фермента от продукта.

Слайд 31Основные подходы для стабилизации ферментов :
Добавление стабилизирующих веществ в среду, в
которой хранится фермент или проводится ферментативная реакция.
2. Химическая модификация фермента.
3. Иммобилизация фермента.
2. Химическая модификация фермента.
3. Иммобилизация фермента.

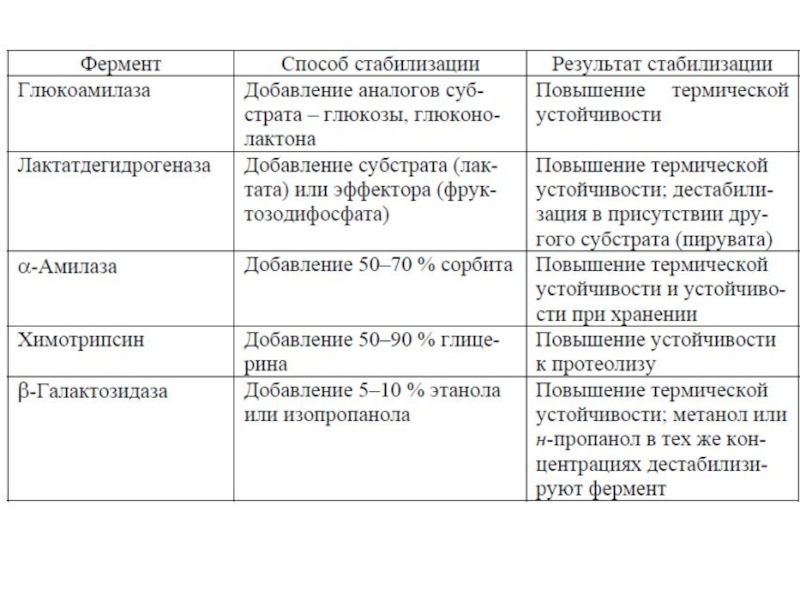
Слайд 32Стабилизация ферментов с помощью:
Субстратов или их аналогов:
Фермент-субстратный комплекс часто
более устойчив, чем свободный фермент.
Пример: Лактатдегидрогеназа в присутствии лактата более термоустойчив.
Пример: Лактатдегидрогеназа в присутствии лактата более термоустойчив.

Слайд 33Стабилизация ферментов с помощью:
2. Органических растворителей:
Многоатомные спирты стабилизируют некоторые ферменты
за счет повышения устойчивости внутримолекулярных водородных связей белка.
Пример: Химотрипсин в присутствии 50–90 % глицерина более устойчив к протеолизу
Пример: Химотрипсин в присутствии 50–90 % глицерина более устойчив к протеолизу

Слайд 34Стабилизация ферментов с помощью:
3. Солей:
При низких концентрациях солей (
Zn2+, Mn2+,Fe2+ и др. могут специфично взаимодействовать с металлопротеинами. Некоторые из них - кофакторы.
Са2+ способен стабилизировать третичную структуру ряда белков благодаря образованию ионных связей с двумя различными аминокислотными остатками.
Пример: у α-Амилазы (из bacillus caldolyticus)
Са2+ значительно повышает термическую устойчивость.
Са2+ способен стабилизировать третичную структуру ряда белков благодаря образованию ионных связей с двумя различными аминокислотными остатками.
Пример: у α-Амилазы (из bacillus caldolyticus)
Са2+ значительно повышает термическую устойчивость.

Слайд 36Химическая модификация фермента
1. Фермент принимает более стабильную конформацию.
2. Введение в белок
новых функциональных групп приводит к образованию дополнительных стабилизирующих водородных связей или солевых мостиков.
3. При использовании неполярных соединений усиливаются гидрофобные взаимодействия.
4. Модификация гидрофобных областей поверхности белка гидрофильными соединениями уменьшает площадь неблагоприятного контакта внешних неполярных остатков с водой.
Пример: глутаровый альдегид
3. При использовании неполярных соединений усиливаются гидрофобные взаимодействия.
4. Модификация гидрофобных областей поверхности белка гидрофильными соединениями уменьшает площадь неблагоприятного контакта внешних неполярных остатков с водой.
Пример: глутаровый альдегид

Слайд 37Иммобилизация фермента позволяет:
Повысить устойчивость фермента (нагреванию, автолизу, действию агрессивных сред и
т. д)
Многократно использовать фермент
Отделять фермент от реагентов и продуктов реакции.
Прерывать реакцию в нужный момент.
Многократно использовать фермент
Отделять фермент от реагентов и продуктов реакции.
Прерывать реакцию в нужный момент.
Многократно использовать ферментОтделять")
Слайд 38Иммобилизованные ферменты – это препараты ферментов, молекулы которых связаны с носителем,
сохраняя при этом полностью или частично свои каталитические свойства.
Методы иммобилизации:
Химические
Физические
Методы иммобилизации:
Химические
Физические

Слайд 39Методы иммобилизации:
В качестве носителей могут применяться:
1)Органические материалы:
1.1) природные (полисахариды, белки, липиды)
1.2) синтетические полимерные носители
2) Неорганические материалы (матрицы на основе силикагеля, глины, керамики, природных минералов и т. д.)
Органические материалы:1.1) природные (полисахариды, белки, липиды) 1.2) синтетические полимерные носители2)")
Слайд 40Методы физической иммобилизации:
1) адсорбция фермента на нерастворимом носителе в результате электростатических,
гидрофобных, вандер-ваальсовых и др. взаимодействий;
2) включение фермента в полупроницаемую капсулу, в полупроницаемую мембрану;
3) механическое включение фермента в гелевые структуры;
4) Включение в двухфазную систему.
2) включение фермента в полупроницаемую капсулу, в полупроницаемую мембрану;
3) механическое включение фермента в гелевые структуры;
4) Включение в двухфазную систему.
 адсорбция фермента на нерастворимом носителе в результате электростатических, гидрофобных, вандер-ваальсовых и др.")
Слайд 41Методы физической иммобилизации:
1)адсорбция фермента на нерастворимом носителе
Достигается путем контакта водного раствора
фермента с носителем.
адсорбция фермента на нерастворимом носителеДостигается путем контакта водного раствора фермента с носителем.")
Слайд 42Методы физической иммобилизации:
1)адсорбция фермента на нерастворимом носителе
Факторы влияющие на адсорбцию:
Удельная поверхность
и пористость носителя
Значение рН (на не ионообменниках мах адсорбция в изоэлектрической точке белка)
Ионная сила раствора (возрастание ионной силы –десорбция фермента, но иногда обратная ситуация “высаливание”)
Концентрация фермента.
Температура ( с одной стороны денатурация, с другой ускоренная диффузия)
Значение рН (на не ионообменниках мах адсорбция в изоэлектрической точке белка)
Ионная сила раствора (возрастание ионной силы –десорбция фермента, но иногда обратная ситуация “высаливание”)
Концентрация фермента.
Температура ( с одной стороны денатурация, с другой ускоренная диффузия)
адсорбция фермента на нерастворимом носителеФакторы влияющие на адсорбцию:Удельная поверхность и пористость носителяЗначение рН")
Слайд 43Методы физической иммобилизации:
1)адсорбция фермента на нерастворимом носителе
Преимущества:
Относительная простота методики
Доступность носителей
Недостатки:
Недостаточная прочность
связывания
Многие носители биодеградируемы
Многие носители биодеградируемы
адсорбция фермента на нерастворимом носителеПреимущества:Относительная простота методикиДоступность носителейНедостатки:Недостаточная прочность связыванияМногие носители биодеградируемы")
Слайд 44Методы физической иммобилизации:
2) включение фермента в полупроницаемую капсулу, в полупроницаемую мембрану
 включение фермента в полупроницаемую капсулу, в полупроницаемую мембрану")
Слайд 45Методы физической иммобилизации:
2) включение фермента в полупроницаемую капсулу, в полупроницаемую мембрану
Преимущества:
Относительная
простота методики
Защита от микроорганизмов
Нет диффузионных ограничений (т.к. отношение поверхности к площади велико и толщина мембраны невелика)
Недостатки:
Биодеградируемы
Не применим для высокомолекулярных субстратов
Защита от микроорганизмов
Нет диффузионных ограничений (т.к. отношение поверхности к площади велико и толщина мембраны невелика)
Недостатки:
Биодеградируемы
Не применим для высокомолекулярных субстратов
 включение фермента в полупроницаемую капсулу, в полупроницаемую мембрануПреимущества:Относительная простота методики Защита от")
Слайд 46Методы физической иммобилизации:
3) механическое включение фермента в гелевые структуры
Фермент включается в
трехмерную сетку полимерных цепей, образующих гель.
 механическое включение фермента в гелевые структурыФермент включается в трехмерную сетку полимерных цепей,")
Слайд 47Методы физической иммобилизации:
3) механическое включение фермента в гелевые структуры
Необходимо учитывать:
Соответствие размера
пор размеру фермента.
Природа матрицы (т.к. она создает микроокружение для фермента, может создавать рН отличное от рН раствора и повышать сродство субстрата к матрице, что повышает скорость ферментативной реакции )
Природа матрицы (т.к. она создает микроокружение для фермента, может создавать рН отличное от рН раствора и повышать сродство субстрата к матрице, что повышает скорость ферментативной реакции )
 механическое включение фермента в гелевые структурыНеобходимо учитывать:Соответствие размера пор размеру фермента.Природа матрицы")
Слайд 48Методы физической иммобилизации:
3) механическое включение фермента в гелевые структуры
Преимущества:
Относительная простота методики
Повышенная
механическая, химическая и тепловая стойкость матриц.
3) Происходит стабилизация фермента
4) Фермент защищен от бактериального повреждения
Недостатки:
1) Не применим для высокомолекулярных субстратов
3) Происходит стабилизация фермента
4) Фермент защищен от бактериального повреждения
Недостатки:
1) Не применим для высокомолекулярных субстратов
 механическое включение фермента в гелевые структурыПреимущества:Относительная простота методикиПовышенная механическая, химическая и тепловая")
Слайд 49Методы физической иммобилизации:
4) Включение в двухфазную систему
Фермент растворим только в одной
из фаз, а продукт - в другой
Позволяет работать в высокомолекулярными субстратами.
Позволяет работать в высокомолекулярными субстратами.
 Включение в двухфазную системуФермент растворим только в одной из фаз, а продукт")
Слайд 50Методы химической иммобилизации:
Образовании ковалентных связей между ферментом и носителем.
Преимущества:
1) Высокая прочность
конъюгата
2) Можно повышать стабильность фермента
2) Можно повышать стабильность фермента
 Высокая прочность конъюгата2) Можно повышать стабильность")
Слайд 51При иммобилизации ферментов необходимо соблюдать следующие условия:
1. Активные группы матрицы не
должны блокировать каталитический центр фермента.
2. Иммобилизации не должны приводить к потере активности фермента.
2. Иммобилизации не должны приводить к потере активности фермента.

Слайд 52
Весьма перспективным является использование в качестве биокатализаторов иммобилизованных клеток.
Т.к. можно избежать:
дорогостоящие
стадии выделения и очистки ферментов
2) необходимости их последующей стабилизации
2) необходимости их последующей стабилизации

Слайд 53Термозимы
Стабильны в условиях высокой температуры, высоких концентраций солей и экстремальных значений
рН.
Гипертермофильные микроорганизмы, встречающиеся среди Archaea и Bacteria, живут при температурах 80–100 °С.
Гипертермофильные микроорганизмы, встречающиеся среди Archaea и Bacteria, живут при температурах 80–100 °С.

Слайд 54Механизмы ответственны за термоустойчивость ферментов у термозимов:
Между мезофильными и термофильными версиями
ферментов - высокая степень гомологии последовательности и структуры.
Так, последовательности термостабильных дегидрогеназ из Pyrococcus и Thermotoga на 35 и 55% соответственно идентичны последовательности мезофильной дегидрогеназы из Clostridium.
Так, последовательности термостабильных дегидрогеназ из Pyrococcus и Thermotoga на 35 и 55% соответственно идентичны последовательности мезофильной дегидрогеназы из Clostridium.

Слайд 55Было обнаружено, что дегидрогеназа из Pyrococcus furiosus (Tm == 105 °C)
содержит 35 изолейцинов, в то время как дегидрогеназы из Thermotoga maritima (Tm = 95 °C) и Clostridium symbiosum (Tm = 55 °C) только 21 и 20 изолейцинов соответственно.
Термостабильные ферменты содержат меньше глицина: Cs дегидрогеназа содержит 48 остатков глицина, а дегидрогеназы из Tm и Pf только
39 и 34 глицина соответственно.
Больше изолейцина и меньше глицина.
Термостабильные ферменты содержат меньше глицина: Cs дегидрогеназа содержит 48 остатков глицина, а дегидрогеназы из Tm и Pf только
39 и 34 глицина соответственно.
Больше изолейцина и меньше глицина.
 содержит 35 изолейцинов, в")
Слайд 56Возросшая термостабильность коррелирует:
с увеличением жесткости белковой структуры за счет уменьшения содержания
остатков глицина,
с улучшением гидрофобных контактов в ядре дегидрогеназы из Pf в результате замены валина изолейцином. (В результате сайт-направленного мутагенеза приводящего к замене изолейцина на валин термостабильность мутантов уменьшалась).
с улучшением гидрофобных контактов в ядре дегидрогеназы из Pf в результате замены валина изолейцином. (В результате сайт-направленного мутагенеза приводящего к замене изолейцина на валин термостабильность мутантов уменьшалась).

Слайд 57Механизмы стабилизации:
• минимизация доступной площади гидрофобной поверхности белка;
• оптимизация упаковки атомов
белковой молекулы (минимизация отношения поверхность/объем);
• оптимизация распределения зарядов (достигается благодаря устранению отталкивающих взаимодействий, а также в результате организации взаимодействий между зарядами в своеобразную сеть)
Уменьшение количества впадин
• оптимизация распределения зарядов (достигается благодаря устранению отталкивающих взаимодействий, а также в результате организации взаимодействий между зарядами в своеобразную сеть)
Уменьшение количества впадин

Слайд 58Применение ферментов из экстремофилов
Современные технологии молекулярной биологии и генной инженерии позволяет:
1)
получать достаточные количества ферментов из экстремофилов для их последующего
анализа и практического применения.
2)клонирование и экспрессия этих ферментов в мезофильных организмах.
анализа и практического применения.
2)клонирование и экспрессия этих ферментов в мезофильных организмах.
 получать достаточные количества ферментов")
Слайд 59Применение ферментов из экстремофилов:
Крахмал используется для производства сахаров. Сначала процесс ведется
при (95–105 °С) и при значениях рН 6–6,5.
На следующем этапе температура снижается до 60°С и рН=4,5.
Использование термостабильных ферментов (α-амилазы, глюкоамилазы, ксилозоизомеразы), выделенных из гипертермофилов, позволит:
проводить процесс в одну стадию и при одних и тех же условиях
отказаться от дорогостоящих ионообменников
На следующем этапе температура снижается до 60°С и рН=4,5.
Использование термостабильных ферментов (α-амилазы, глюкоамилазы, ксилозоизомеразы), выделенных из гипертермофилов, позволит:
проводить процесс в одну стадию и при одних и тех же условиях
отказаться от дорогостоящих ионообменников
 и")
Слайд 60Применение ферментов из экстремофилов:
Наиболее термостабильные α-амилазы были обнаружены у archaea Pyrococcus
woesei,
Pyrococcus furiosus, Desulfurococcus mucosus, Pyrodictium abyssi и Staphylothermus
marinus. Гены амилазы из Pyrococcus sp. были
клонированы и экспрессированы в E.coli и Bacillus subtilis.
Pyrococcus furiosus, Desulfurococcus mucosus, Pyrodictium abyssi и Staphylothermus
marinus. Гены амилазы из Pyrococcus sp. были
клонированы и экспрессированы в E.coli и Bacillus subtilis.

Слайд 61Применение ферментов из экстремофилов:
Протеолитические ферменты
Сериновые щелочные протеиназы широко используются в качестве
добавок к моющим средствам.
Протеиназы из экстремофилов сохраняют нативность при высоких температурах, в присутствии высоких концентраций детергентов и других денатурирующих агентов. Pyrococcus, Thermococcus, Staphylothermus, Desulfurococcus и Sulfolobus. Максимальную активность эти ферменты проявляют при температурах
от 90 до 110 °С и значениях рН от 2 до 10
Протеиназы из экстремофилов сохраняют нативность при высоких температурах, в присутствии высоких концентраций детергентов и других денатурирующих агентов. Pyrococcus, Thermococcus, Staphylothermus, Desulfurococcus и Sulfolobus. Максимальную активность эти ферменты проявляют при температурах
от 90 до 110 °С и значениях рН от 2 до 10

Слайд 62Применение ферментов из экстремофилов:
ДНК-полимеразы
Термостабильные ДНК-полимеразы используются в ПЦР и играют важную
роль в генной инженерии. Термостабильные полимеразы были обнаружены у гипертермофилов Pyrococcus furiosus и Pyrococcus litoralis, а также у термофилов Thermus aquaticus.