Понятие о нативных формах. Энергетические переходы в молекуле хлорофилла. Белковые комплексы, содержащий хлорофилл. Миграция энергии. Окислительно-восстановительные реакции с участием хлорофилла. Продукция активных форм кислорода.
2. Каротиноиды. Общее представление о биосинтезе. Протекторная роль каротиноидов в фотосистемах. Виолаксантиновый цикл и его роль в регуляции распределения энергии. Защитная функция каротиноидов. Каротиноиды как предшественники АБК. Экологическая роль каротиноидов.
3. Продукция активных форм кислорода с участием возбужденного хлорофилла. Экологические факторы, способствующие образованию синглетного кислорода. Защитные механизмы. Роль виолаксантинового (ксантофиллового) цикла в регуляции распределения энергии квантов света.
4. Антенные комплексы. Подвижные и неподвижные комплексы. Фикобилисомы. Фикобилины как дополнительные ферменты фотосинтеза у водорослей и цианобактерий. Нативные формы хлорофиллов в антенных комплексах. Понятие о фотосинтической единице. Факторы, влияющие на ассоциацию светособирающего комплекса с ФС II и ФС I.
5. Строение и функционирование ФС I. Ассоциация и диссоциация с подвижным светособирающим комплексом. Кооперация работы ФС I и ФС II. Локализация ФС I в мембране тилакоидов.
6. Строение и функционирование фотосистемы II. Водоокисляющий комплекс и реакции образования кислорода. Работа реакционного центра. Участие ФС II в нециклическом потоке ē. Работа ФС II в циклическом режиме. Локализация ФС II и взаимодействие со светособирающим комплексом.
7. Нециклический, циклический и псевдоциклический транспорт электрона. Последовательность переносчиков. Цикл вокруг фотосистемы II. Реакция хлородыхания как регуляция редокс-статуса пула пластохинонов. Подвижные переносчики в составе комплексов. Одно- и двухэлектронные переносчики.
8. Взаимосвязь между фотосинтетической функцией и ультраструктурой хлоропластов. Локализация белковых комплексов на мембранах тилакоидов (ССК, ФСII, ФСI, цитохром-b/f-комплекс, АТФ-синтаза). Переключение с нециклического на циклический поток электронов по ЭТЦ фотосинтеза и связанное с ним изменение локализации комплексов.
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Хлорофиллы. Общие принципы организации молекулы. Основные этапы биосинтеза презентация
Содержание
- 1. Хлорофиллы. Общие принципы организации молекулы. Основные этапы биосинтеза
- 3. Функции пластид Фотосинтез – NB Синтез: все
- 4. Взаимопревращения пластид контролируются ядерным геномом
- 6. ССКII
- 7. Гетерогенная организация тилакоидных структур
- 8. Гены хлоропластов Транскрипция. 4 гена субъединиц
- 9. Фоторецепторная система фотосинтеза строится на основе двух
- 10. ФИКОБИЛИНЫ
- 11. Основные структурные особенности молекулы хлорофилла Конъюгированная система
- 12. Хлорофиллов >10: Хл. а, b, c1, с2,
- 13. Спектры поглощения ФАР : 380 – 710
- 14. E = hν =
- 15. Биосинтез хлорофилла Глутамат → Глутамил-тРНК →Глутамат-1-семиальдегид →
- 17. Основные структурные особенности молекулы хлорофилла Конъюгированная система
- 18. Хлорофилл способен к окислительно-восстановительным превращениям. «Реакция
- 19. S0 – основное синглетное состояние
- 20. Энергетические уровни хлорофилла
- 21. Схема Яблонского
- 22. R СН3 СН3 СН3 СН3
- 23. Два пути синтеза изопреноидов в растениях: «мевалонатный» в цитозоле и «альтернативный» в хлоропластах
- 24. Каротиноиды
- 25. Другие важные изопреноиды 1. Фитольный хвост хлорофилов
- 27. Функции каротиноидов 1. Антенны (400 – 500
- 28. Защитная функция каротиноидов
- 29. 1.Зеаксантин 2. Антераксантин 3. Виолаксантин Виолаксантиновый цикл
- 30. 2О2 + 4Н. (НАДФН)
- 31. Виолаксантиновый цикл Энергия S1 ниже S1
- 32. Кофакторы ЭТЦ фотосинтеза: знакомые все лица...
- 33. Строение хинонов O || || CH3 CH3
- 34. Редокс превращения в молекулах хинонов
- 35. Кванты света запускают последовательность окислительно-восстановительных реакций на внутренней мб хлоропласта
- 36. Z-схема
- 37. -1,0 -0,8 -0,6 -0,4
- 39. Гетерогенная организация тилакиодный структур
- 40. Фотосинтетичекая единица Фотосинтетическая единица– представляет взаимодействие светособирающего комплекса и реакционного центра фотосистемы.
- 41. ССК (II) Белки - 25,27 и 28
- 42. ССК I Белки 17-24 кДа Хл
- 43. СветоСобирающий Комплекс (ССК) Реакционный Центр
- 45. ЭТЦ фотосинтеза
- 46. Структура фотосистемы II Феофитин
- 47. Водоокисляющая система
- 48. So Mn2+ Mn3+ Mn4+ Mn4+ S1
- 49. Mn Структура Mn кластера
- 50. Модели работы системы фотоокисления воды
- 51. Организация ЭТЦ фотосинтетического аппарата
- 52. В6f-комплекс: два такта работы Q-цикла
- 53. В6f-комплекс: два такта работы Q-цикла
- 54. Структура RC фотосистемы I 13 белков: А
- 55. Нециклический транспорт электронов е Итоги: Генерация
- 56. Нециклический электронный транспорт
- 57. Циклический транспорт электронов е Итог: Генерация протонного градиента
- 58. ЦИКЛИЧЕСКИЙ ЭЛЕКТРОННЫЙ ТРАНСПОРТ
- 59. ПСЕВДОЦИКЛИЧЕСКИЙ ЭЛЕКТРОННЫЙ ТРАНСПОРТ Н2О
- 60. АТФ-азный комплекс P.Boyer&J.Walker (нобелевская премия, 1997)
- 61. Механизм работы АТФ-синтазы P. Boyer и J.
- 62. Вращение нити актина в реальном времени F1 Нить актина флуоресцирующая
Слайд 11. Хлорофиллы. Общие принципы организации молекулы. Основные этапы биосинтеза. Спектр поглощения хлорофиллов.

Слайд 3Функции пластид
Фотосинтез – NB
Синтез: все жирные кислоты, многие аминокислоты, синтез пуринов
и пиримидинов, альтернативный путь синтеза изопреноидов (в том числе в спецпластидах – лейкопластах), шикиматный путь (параллельно цитозолю)
Восстановление нитритов, сульфатов
Запас (крахмал) – временный (хлоропласты), долгосрочный (амилопласты)
Экологические – окраска плодов, цветков (хромопласты – каротиноиды).
Пластиды – «фабрика горячих и вредных производств» растительной клетки
Восстановление нитритов, сульфатов
Запас (крахмал) – временный (хлоропласты), долгосрочный (амилопласты)
Экологические – окраска плодов, цветков (хромопласты – каротиноиды).
Пластиды – «фабрика горячих и вредных производств» растительной клетки




Слайд 8Гены хлоропластов
Транскрипция. 4 гена субъединиц пластидной РНК-полимеразы (rpo)
Синтез белка.
- 4 гена рРНК (оперон rrn)
- около 20 генов белков пластидных рибосом (rpl/rps)
- около 30 генов тРНК (trn)
Фотосинтез. - 6 генов белков фотосистемы I (psa)
- 14 генов белков фотосистемы II (psb)
- 6 генов ЭТЦ фотосинтеза (pet)
- 6 генов пластидной АТФ-зы (atp)
- ген большой субъединицы Рубиско (rbcL)
4. Около 20 генов с другими функциями
- гены пластидной НАД Н-дегидрогеназа,
- гены биосинтеза жирных кислот и др.
Всего: 110 - 120 генов, из них около 40 – «рабочих»
и около 60 – «домашнего хозяйства».
- около 20 генов белков пластидных рибосом (rpl/rps)
- около 30 генов тРНК (trn)
Фотосинтез. - 6 генов белков фотосистемы I (psa)
- 14 генов белков фотосистемы II (psb)
- 6 генов ЭТЦ фотосинтеза (pet)
- 6 генов пластидной АТФ-зы (atp)
- ген большой субъединицы Рубиско (rbcL)
4. Около 20 генов с другими функциями
- гены пластидной НАД Н-дегидрогеназа,
- гены биосинтеза жирных кислот и др.
Всего: 110 - 120 генов, из них около 40 – «рабочих»
и около 60 – «домашнего хозяйства».
Синтез белка. - 4 гена рРНК (оперон rrn)")
Слайд 9Фоторецепторная система фотосинтеза строится на основе двух основных химических структур:
1.-
тетрапирролы, образующие циклическую структуру хлорофилла (магний-порфирины), а также открытую тетрапиррольную структуру другой группы пигментов – фикобилинов;
2. – полиизопреноиды, которые являются структурной основой большого и разнообразного класса пигментов – каротиноидов.
2. – полиизопреноиды, которые являются структурной основой большого и разнообразного класса пигментов – каротиноидов.

")
Слайд 11Основные структурные особенности молекулы хлорофилла
Конъюгированная система двойных связей: основная 18-членная π-система
+ дополнительные в I, II, V кольцах.
Mg – минимум электроотрицательности; изменяет симметрию молекулы хлорофилла; «активирует» электроны пиррольных азотов
V-кольцо – «форбиновая структура»: две важных группы: карбонильная при С9 (участвует в n – π переходах) и кетоэфирная при С10 – транс- (хл-л а) или цис- (хл-л а’).
Гидрофобный «хвост» (обычно С20 – фитол). Структурная роль.
Mg – минимум электроотрицательности; изменяет симметрию молекулы хлорофилла; «активирует» электроны пиррольных азотов
V-кольцо – «форбиновая структура»: две важных группы: карбонильная при С9 (участвует в n – π переходах) и кетоэфирная при С10 – транс- (хл-л а) или цис- (хл-л а’).
Гидрофобный «хвост» (обычно С20 – фитол). Структурная роль.

Слайд 12Хлорофиллов >10:
Хл. а, b, c1, с2, d, e;
Б-хл. a, b, c,
d.
Единственная
молекула которая
может:
1. Поглощать hν и
трансформировать
эту энергию в е-*
2. Обратимо
окисляться, т.е.
отдавать е-*
Единственная
молекула которая
может:
1. Поглощать hν и
трансформировать
эту энергию в е-*
2. Обратимо
окисляться, т.е.
отдавать е-*

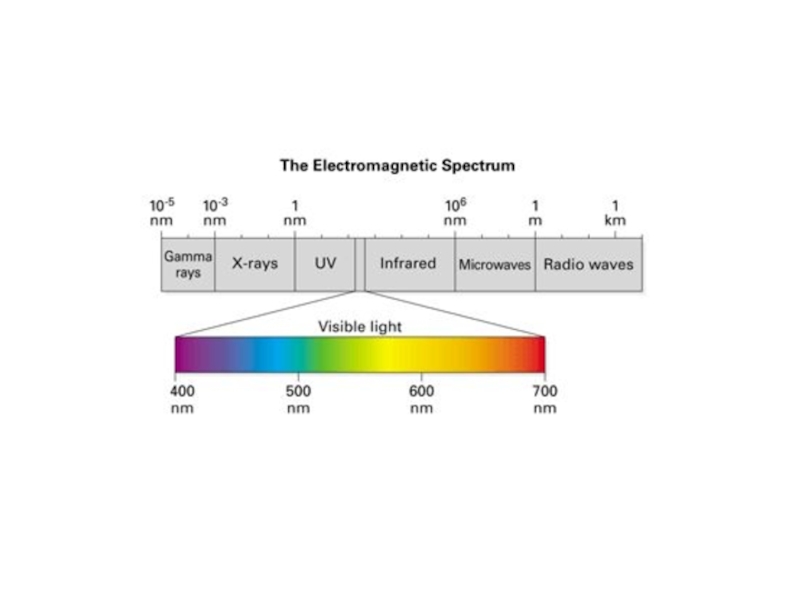
Слайд 13Спектры поглощения
ФАР : 380 – 710 нм
Каротиноиды: 400-550 нм главный максимум:
480 нм
Хлорофиллы:
в красной области спектра 640-700 нм
в синей - 400-450 нм
Почему видимый
свет ?
1. Оптимум
энергии: 1 – 3 эв.
2. Максимальная
«прозрачность»
атмосферы для
этих длин волн –
более 50%.


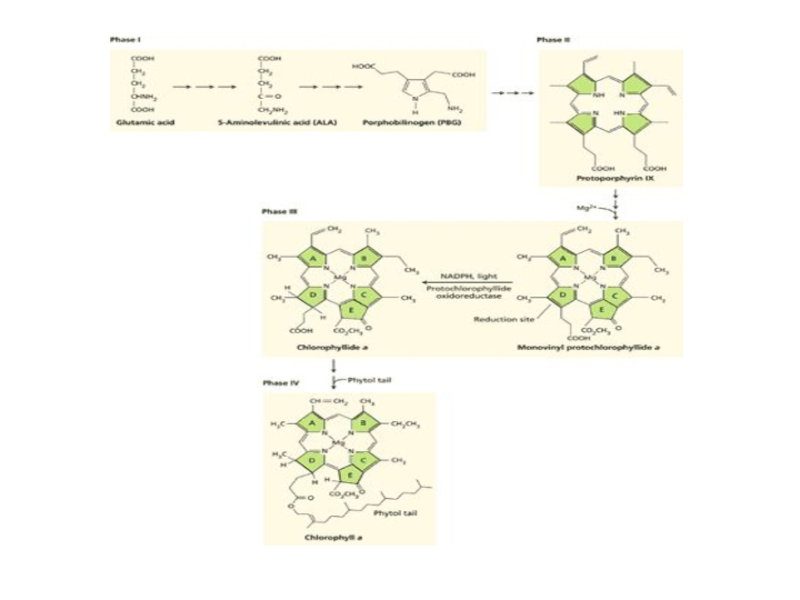
Слайд 15Биосинтез хлорофилла
Глутамат → Глутамил-тРНК →Глутамат-1-семиальдегид →
δ-Аминолевулиновая кислота
δ-Аминолевулиновая кислота →Порфобилиноген →Гидроксиметилбилан
→УропорфириногенIII →КопропорфироногенIII →ПротопирфириногенIX →Протопорфирин IX
Mg-хелатаза
Mg-протопорфирин → Mg-протопорфиринмонометиловый эфир →Дивинилпротохлорофиллид а →Моновинилпротохлорофиллид а
Протохлорофиллид →СВЕТ! →Хлорофиллид а →Хлорофилл а →Хлорофилл b
Mg-хелатаза
Mg-протопорфирин → Mg-протопорфиринмонометиловый эфир →Дивинилпротохлорофиллид а →Моновинилпротохлорофиллид а
Протохлорофиллид →СВЕТ! →Хлорофиллид а →Хлорофилл а →Хлорофилл b

Слайд 17Основные структурные особенности молекулы хлорофилла
Конъюгированная система двойных связей: основная 18-членная π-система
+ дополнительные в I, II, V кольцах.
Mg – минимум электроотрицательности; изменяет симметрию молекулы хлорофилла; «активирует» электроны пиррольных азотов
V-кольцо – «форбиновая структура»: две важных группы: карбонильная при С9 (участвует в n – π переходах) и кетоэфирная при С10 – транс- (хл-л а) или цис- (хл-л а’).
Гидрофобный «хвост» (обычно С20 – фитол). Структурная роль.
Mg – минимум электроотрицательности; изменяет симметрию молекулы хлорофилла; «активирует» электроны пиррольных азотов
V-кольцо – «форбиновая структура»: две важных группы: карбонильная при С9 (участвует в n – π переходах) и кетоэфирная при С10 – транс- (хл-л а) или цис- (хл-л а’).
Гидрофобный «хвост» (обычно С20 – фитол). Структурная роль.

Слайд 18Хлорофилл способен к окислительно-восстановительным превращениям.
«Реакция Красновского»
Д - (Хл-Хл)680*
- А
Д - (Хл-Хл) 680+ - А-
Д+ - (Хл-Хл) 680 - А-
680* - А Д - (Хл-Хл)")
Слайд 19S0 – основное синглетное состояние
S*2 , S*1 – синглетные возбужденные состояния
Т* - триплетное возбужденное состояние
Энергетические состояния молекулы хлорофилла



Слайд 22
R
СН3
СН3
СН3
СН3
СН3
СН3
СН3
СН3
СН3
СН3
(R = H-) β - каротин
(R= HO -
ксантофилл
каротиноиды
область поглощения 400-500 нм
 β - каротин(R= HO - ксантофиллкаротиноидыобласть поглощения 400-500 нм")
Слайд 23Два пути синтеза изопреноидов в растениях: «мевалонатный» в цитозоле и «альтернативный»
в хлоропластах


Слайд 25Другие важные изопреноиды
1. Фитольный хвост хлорофилов
2. Убихиноны и пластохиноны, филлохинон
3. Стероидные
соединения
4. Некоторые растительные гормоны
(гибереллины, брассиностероиды,
абсцизовая кислота)
5. Каучук и гуттаперча
4. Некоторые растительные гормоны
(гибереллины, брассиностероиды,
абсцизовая кислота)
5. Каучук и гуттаперча
Гиббереллин А1 (GA1)
Абсцизовая кислота (АБК)
Брассинолид

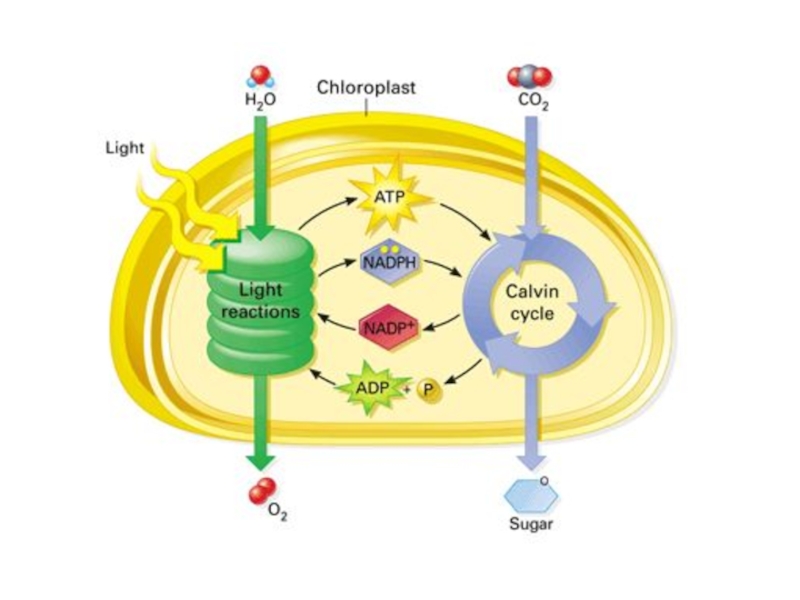
Слайд 26 ФОТОСИНТЕЗ
О2*
О2.-
Н2О2
Фотоингибирование : повреждение
белка Д1
РЦ ФС 2
разрушение
хлорофилла
перекисное окисление липидов мембран

Слайд 27Функции каротиноидов
1. Антенны (400 – 500 нм)
2. Структурная (организация ССК)
3. Фотопротекторная
(виолаксантиновый цикл)
4. Защита от УФ и высоких интенсивностей света
a/ R˚ RН
hν RH carо car
PРЦ 1Р 3Р
3car 1car 3O2 P + 1O2
3car 1car
b/ car + P+680 car+ + P680
5. Биосинтез сигнальных молекул (Абсцизовая кислота, АБК)
4. Защита от УФ и высоких интенсивностей света
a/ R˚ RН
hν RH carо car
PРЦ 1Р 3Р
3car 1car 3O2 P + 1O2
3car 1car
b/ car + P+680 car+ + P680
5. Биосинтез сигнальных молекул (Абсцизовая кислота, АБК)
2. Структурная (организация ССК)3. Фотопротекторная (виолаксантиновый цикл)4. Защита от")
Слайд 28Защитная функция каротиноидов
Хл So
Хл*S1 Хл*Т
Хл*Т + O2T Хл So + O2 S*
Кар.So + Хл*Т Кар*Т + ХлS0
Кар.So + тепло
Кар.So + O2S* Кар*Т + О2
Кар.So + тепло


")

Слайд 32Кофакторы ЭТЦ фотосинтеза: знакомые все лица...
2Fe-2S и 4Fe-4S-белки, хиноны (пластохиноны
и филохинон), цитохромы
, цитохромы")

Слайд 34Редокс превращения в молекулах хинонов
е-
е- + 2Н+
Q + 1e-
Q•– + 1e- + 2H+ QH2
||
|
O
O
O•
|
O-
|
|
OH
OH
||
пластохинон
пластохинол
семихинон

Слайд 35Кванты света запускают последовательность окислительно-восстановительных реакций на внутренней мб хлоропласта


Слайд 37
-1,0
-0,8
-0,6
-0,4
+0,5
+0,8
+1,0
Н2О/O2
S
Фео
QA,
QB
PQ
FeS
цит f
0,0
Р700
Пц
Р700*
НАДФ+/НАДФН
Хл 695
А1 витамин К
Fx
FA FeS белки
FB
Фд
Z – схема
Z-схема фотосинтеза
Р680*
Р680


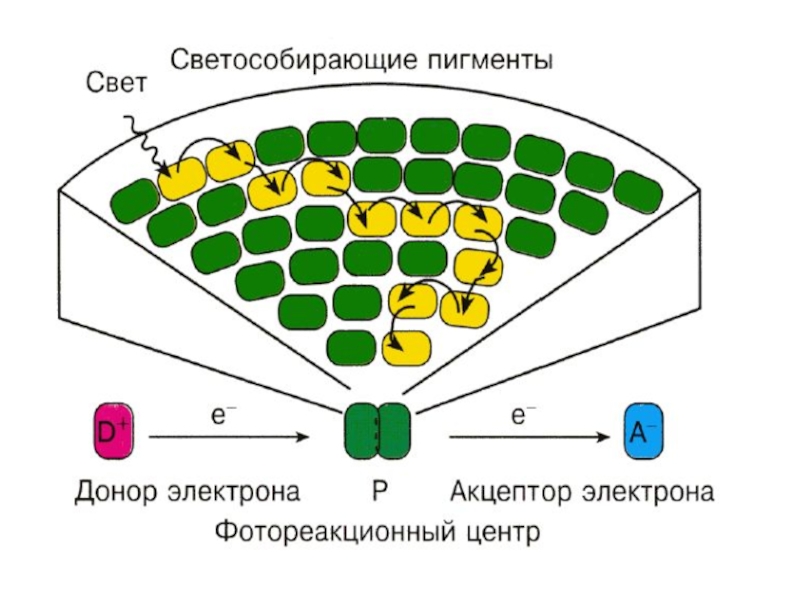
Слайд 40Фотосинтетичекая единица
Фотосинтетическая единица– представляет взаимодействие светособирающего комплекса и реакционного центра фотосистемы.

Слайд 41ССК (II)
Белки - 25,27 и 28 кДа
Хл а ~ 150
Хл b
~ 100
Каротиноиды ~ 50
Каротиноиды ~ 50
АНТЕННА ФС2
Белки -СР 24, СР26, СР29
Хл а
Хл b
ксантофиллы
РЦ
СР43
СР47
Фотосистема 2
ФОКУСНАЯ АНТЕННА НА БЕЛКАХ СР43 и СР47
Хл а ∼30; β-каротин - 2
Реакционный центр
Белки - 25,27 и 28 кДаХл а ~ 150Хл b ~ 100Каротиноиды ~ 50АНТЕННА")
Слайд 42
ССК I
Белки 17-24 кДа
Хл а ∼ 80-120
Хл b ∼ ?
каротиноиды ∼50
Кор-комплекс
Хл
а ∼ 90-100
β каротин ∼ 20
β каротин ∼ 20
РЕАКЦИОННЫЙ ЦЕНТР

Слайд 43
СветоСобирающий Комплекс
(ССК)
Реакционный
Центр
(РЦ)
Неподвижная
Антенна
Процесс миграции энергии
е-
Н20
02+2Н+
Реакционный Центр(РЦ)НеподвижнаяАнтеннаПроцесс миграции энергии е-Н2002+2Н+")



Слайд 48
So
Mn2+ Mn3+
Mn4+ Mn4+
S1
Mn3+ Mn3+
Mn4+ Mn4+
S2
Mn3+ Mn4+
Mn4+ Mn4+
S3
Mn4+ Mn4+
Mn4+ Mn4+
S4 ?
2Н2О
4Н+ +
О2
e-
e-
e-
e-
hλ
СИСТЕМА ФОТООКИСЛЕНИЯ ВОДЫ : So-S4-цикл






Слайд 54Структура RC фотосистемы I
13 белков:
А – 83 kDa, 751 a-к
В -
82,5 kDa, 735 a-к
Гетеродимер, на нем:
Р700, А0, А1, Fx
С – 8,9 kDa, - FA, FB
D (19 kDa),
E – связь с Fd
F (19 kDa) - связь с Pc
Гетеродимер, на нем:
Р700, А0, А1, Fx
С – 8,9 kDa, - FA, FB
D (19 kDa),
E – связь с Fd
F (19 kDa) - связь с Pc

Слайд 55Нециклический транспорт электронов
е
Итоги:
Генерация
протонного
градиента;
2) Синтез
НАДФН
3) Побочный
продукт -
кислород
 СинтезНАДФН3) Побочный продукт - кислород")




")
Слайд 61Механизм работы АТФ-синтазы
P. Boyer и J. Walker ( Нобелевская премия 1997).
Ротационноый механизм
O – open «открыто», T - tight «закрыто» и L – loose «слабосвязанно»
. Ротационноый механизм O –")






