- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Физиология растений. Свет презентация
Содержание
- 1. Физиология растений. Свет
- 2. Отраженный свет Поглощенный свет Свет Хлоропласт Пропущенный свет Грана
- 3. Свет ФД Комплекс цитохромов AДФ
- 4. Свет H2O Хлоропласт Световые реакции НAДФ+ Ф AДФ н + ATФ НАДФH O2
- 5. Свет H2O Хлоропласт НАДФ+ Ф AДФ н
- 6. Цикл Кальвина происходит в строме и начинается
- 7. Это самый распространенный в мире фермент, при это очень неэффективный
- 8. Продукт реакции, содержащий шесть атомов углерода, в
- 9. Для дальнейших превращений ФГК необходимы вещества световой
- 10. Затем происходит восстановление за счет НАДФН и
- 11. Дальнейшее превращение фосфоглицеринового альдегида может происходить 4
- 12. Это второй путь превращения ФГА. Таким
- 13. Фруктозо-1,6-дифосфат дефосфорилируется и превращается во фруктозо-6-фосфат (Ф-6-Ф),
- 14. Эритрозо-4-фосфат Седогептулозо-1,7-дифосфат транскетолаза альдолаза
- 15. Четвертый путь превращения ФГА связан с его
- 16. транскетолаза Пентозо- изомераза Фосфорибулозо- эпимераза фосфорибулокиназа
- 17. Фруктозо-6-фосфат – очень важный сахар, образующийся в
- 18. Цикл Кальвина можно условно подразделить на
- 20. Ribulose bisphosphate (RuBP) 3-Phosphoglycerate Short-lived intermediate
- 21. Ribulose bisphosphate (RuBP) 3-Phosphoglycerate Short-lived intermediate
- 22. Ribulose bisphosphate (RuBP) 3-Phosphoglycerate Short-lived intermediate
- 23. Цикл Кальвина (С3) восстановления СО2 до полисахаридов
- 24. сахароза глюкоза фруктоза
- 25. С2-цикл или Фотодыхание (поглощается кислород, поэтому
- 26. С2-цикл или Фотодыхание 1. Начальная (оксигеназная)
- 27. С2-цикл или Фотодыхание 2. Дефосфорилирование
- 28. С2-цикл или Фотодыхание Транспортная реакция
- 29. С2-цикл или Фотодыхание 7. Восстановление
- 30. С2-цикл или Фотодыхание 9. Детоксификация
- 32. Цикл Хетча – Слэка – Карпилова (С4-цикл)
- 33. В этих кислотах можно обнаружить в первые
Слайд 1Физиология растений
Демидчик Вадим Викторович
(д.б.н., зав. каф. физиологии
и биохимии растений)
")

Слайд 3Свет
ФД
Комплекс
цитохромов
AДФ
+
н
H+
ATФ
Ф
ATФ
синтаза
В цикл
Кальвина
Строма
(низкая концентрация H+)
Тиллакоидная
мембрана
Внутритиллакоидное
пространство
(высокая активность
H+ concentration)
Строма
(низкая активность H+)
Фотосистема II
Фотосистема I
4 H+
4 H+
ПХ
ПЦ
Свет
НАДФ+
редуктаза
НАДФ+ + H+
НАДФН
+2 H+
H2O
O2
e–
e–
1/2
Тиллакоидная мембранаВнутритиллакоидное пространство(высокая активность H+ concentration)Строма(низкая активность H+)Фотосистема")


Слайд 6Цикл Кальвина происходит в строме и начинается с присоединения СО2 к
Присоединение СО2 к веществу называется карбоксилированием, а фермент, катализирующий такую реакцию, – карбоксилазой.
В данной реакции карбоксилирование происходит с участием фермента рибулозодифосфаткарбоксилазы
(РДФ-карбоксилазы).
Это самый распространенный в мире фермент.
2 цепи – хлоропластная (2 х 55 кДа) и ядерная (8 х 13 кДа), 560 кДа. Mg2+ - кофактор для связывания СО2


Слайд 8Продукт реакции, содержащий шесть атомов углерода, в присутствии воды сразу распадается
С данной реакции начинается цикл Кальвина.
ФГК является, по современным взглядам, первичным продуктом ассимиляции углерода.

Слайд 9Для дальнейших превращений ФГК необходимы вещества световой фазы фотосинтеза: АТФ и
Сначала 3-ФГК фосфорилируется при участии АТФ и образуется 1,3-дифосфоглицериновая кислота.
Реакция катализируется ферментом фосфоглицераткиназой:

Слайд 10Затем происходит восстановление за счет НАДФН и образуется фосфоглицериновый альдегид (ФГА):
Суммарный
Процесс превращения катализируется дегидрогеназой фосфоглицеринового альдегида.
:Суммарный результат второй стадии: восстановление")
Слайд 11Дальнейшее превращение фосфоглицеринового альдегида может происходить 4 путями.
ФГА частично с помощью
Это первый путь превращения ФГА.

Слайд 12Это второй путь превращения ФГА.
Таким образом в клетку поступают две
Это первые углеводы, которые образуются при фотосинтезе.
С помощью альдолазы ФДА соединяется с другой молекулой ФГА, и образуется молекула фруктозо-1,6-дифосфата (ФДФ).

Слайд 13Фруктозо-1,6-дифосфат дефосфорилируется и превращается во фруктозо-6-фосфат (Ф-6-Ф), что сопровождается накоплением в
Фруктозо-6-фосфат в дальнейшем может выйти из цикла и использоваться для синтеза запасных форм углеводов: сахарозы, крахмала, других полисахаридов.
Однако ФГА (третий путь) может реагировать с эквимолярным количеством Ф-6-Ф, в результате образуются равные количества ксилулозо-5-фосфата и эритрозо-4-фосфата (реакция катализируется транскетолазой).
Эритрозо-4-фосфат может взаимодействовать с равным количеством фосфодиоксиацетона (ФДА) при участии фермента альдолазы, что приводит к образованию
седагептулозо-1,7-дифосфата, который
дефосфорилируется до седагептулозо-7-фосфата с
участием седагептулозодифосфатазы.
, что сопровождается накоплением в среде неорганического фосфата. Фруктозо-6-фосфат")

Слайд 15Четвертый путь превращения ФГА связан с его реакцией с седагептулозо-7-фосфатом с
Ксилулозо-5-фосфат эпимеризуется (Фосфорибулозо-эпимераза),
а рибозо-5-фосфат изомерезуется (РФ-изомераза, другое название – пентозоизомераза)
до рибулозо-5-фосфата,
последняя фосфорилируется за счет АТФ (фермент фосфорибулокиназа) и образуется рибулозо-1,5-дифосфат– первичное соединение цикла Кальвина (акцептор СО2).
 количеств")

Слайд 17Фруктозо-6-фосфат – очень важный сахар, образующийся в цикле Кальвина.
Суммарное выражение его
6СО2 + 12НАДФН + 12Н+ + 18АТФ + 11Н2О →
фруктозо-6-фосфат + 12НАДФ+ + 18АДФ + 17Фн
Примерный энергетический расчет:
18 молекул АТФ содержат около 140 ккал и 12 НАДФН ~ 615 ккал. Итого около 755 ккал энергии затрачивается на суммарный цикл.
В гексозах запасается около 670 ккал/моль.
Таким образом КПД (670 / 755 х 100) составляет около 90 %.
Т.е. 10 % энергии растрачивается на поддержание цикла.

Слайд 18Цикл Кальвина можно условно подразделить
на три фазы:
– карбоксилирующую:
– восстановительную: ФГК → ФГА;
– регенерирующую: ФГА → РДФ.
Примерно одна из 6 молекул ФГА покидает цикл и используется для синтеза полисахаридов.
Остальные пять молекул обратно превращаются в
3 молекулы рибулозо-1,5-дифосфата (акцептора СО2).
По названию первичного продукта цикла Кальвина – ФГК (3 атома С) он получил название
С3-цикла ассимиляции СО2.


Слайд 20Ribulose bisphosphate
(RuBP)
3-Phosphoglycerate
Short-lived
intermediate
Phase 1: Carbon fixation
(Entering one
at a time)
Rubisco
Input
CO2
P
3
6
3
3
P
P
P
P
3-Phosphoglycerate Short-livedintermediatePhase 1: Carbon fixation(Entering oneat a time)RubiscoInputCO2P3633PPPP")
Слайд 21Ribulose bisphosphate
(RuBP)
3-Phosphoglycerate
Short-lived
intermediate
Phase 1: Carbon fixation
(Entering one
at a time)
Rubisco
Input
CO2
P
3
6
3
3
P
P
P
P
ATP
6
6 ADP
P
P
6
1,3-Bisphosphoglycerate
6
P
P
6
6
6 NADP+
NADPH
i
Phase
Reduction
Glyceraldehyde-3-phosphate
(G3P)
1
P
Output
G3P
(a sugar)
Glucose and
other organic
compounds
Calvin
Cycle
3-Phosphoglycerate Short-livedintermediatePhase 1: Carbon fixation(Entering oneat a time)RubiscoInputCO2P3633PPPPATP66 ADPPP61,3-Bisphosphoglycerate6PP666 NADP+NADPHiPhase 2:ReductionGlyceraldehyde-3-phosphate(G3P)1POutputG3P(a sugar)Glucose andother organiccompoundsCalvinCycle")
Слайд 22Ribulose bisphosphate
(RuBP)
3-Phosphoglycerate
Short-lived
intermediate
Phase 1: Carbon fixation
(Entering one
at a time)
Rubisco
Input
CO2
P
3
6
3
3
P
P
P
P
ATP
6
6 ADP
P
P
6
1,3-Bisphosphoglycerate
6
P
P
6
6
6 NADP+
NADPH
i
Phase
Reduction
Glyceraldehyde-3-phosphate
(G3P)
1
P
Output
G3P
(a sugar)
Glucose and
other organic
compounds
Calvin
Cycle
3
3 ADP
ATP
5
P
Phase 3:
Regeneration of
the CO2 acceptor
(RuBP)
G3P
3-Phosphoglycerate Short-livedintermediatePhase 1: Carbon fixation(Entering oneat a time)RubiscoInputCO2P3633PPPPATP66 ADPPP61,3-Bisphosphoglycerate6PP666 NADP+NADPHiPhase 2:ReductionGlyceraldehyde-3-phosphate(G3P)1POutputG3P(a sugar)Glucose andother organiccompoundsCalvinCycle33")
Слайд 23Цикл Кальвина (С3) восстановления СО2 до полисахаридов локализован в строме хлоропластов.
Там же локализован и биосинтез крахмала из образованных в них гексозофосфатов.
Наиболее обильный сахар клетки – это сахароза.
Он синтезируется из Ф-6-Ф, который образуется из ФГА и ФДА в цитоплазме, вернее в её слое, непосредственно прилегающем к наружной мембране хлоропласта. ФГА и ФДА, по сравнению с другими сахарами С3-цикла (пентозами и гексозами) легче транспортируются через мембраны хлоропластов.
 восстановления СО2 до полисахаридов локализован в строме хлоропластов. Там же локализован и")

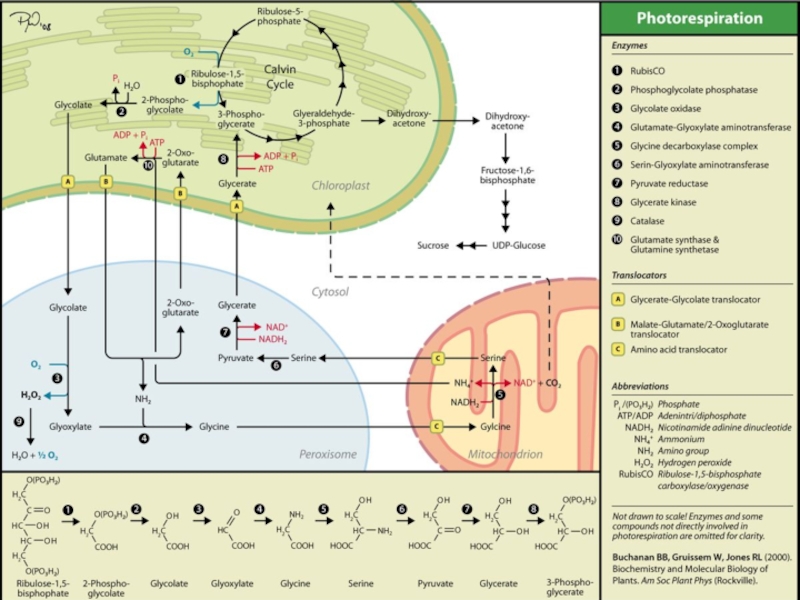
Слайд 25С2-цикл или Фотодыхание
(поглощается кислород, поэтому называется «дыханием»)
Фотодыхание – это
(РДФ-карбоксилаза – «Рубиско») присоединяет к рибулозо-1,5-дифосфату (РДФ) кислород, а не СО2.
Т.е. этот фермент ведет себя как оксигеназа
и катализирует первую реакцию фотодыхания.
Фотодыхание снижает эффективность фотосинтеза у С3-растений (до 25%). Наиболее часто наблюдается при засухе, когда устьица закрыты. Также возрастает с ростом температуры. Часто приводит к потере не только углерода, но и азота в виде иона аммония.
Рубиско имеет намного большее сродство к СО2, чем к О2, однако, концентрация растворенного СО2 при интенсивном фотосинтезе может становиться очень низкой (СО2 и так совсем немного в атмосфере – 0,039% на объем, в отличие от кислорода).
 Фотодыхание – это процесс, в котором рибулозодифосфаткарбоксилаза")
Слайд 26С2-цикл или Фотодыхание
1. Начальная (оксигеназная) реакция;
фермент – «Рубиско»
рибулозо-1,5-дифосфат + О2
3-фосфоглицерат (ФГК) + фосфогликолат + 2Н+
ФГК возвращается обратно в цикл Кальвина, а фосфогликолат метаболизируется с участием пероксисом и митохондрий.
«- рат» на конце слова означает остаток кислоты и часто используется для обозначения кислоты – например, фосфоглицерат и фосфоглицериновая кислоты – одно и тоже.
ФГК
Фос-
фогли-
колат
 реакция;фермент – «Рубиско»рибулозо-1,5-дифосфат + О2 → 3-фосфоглицерат (ФГК) +")
Слайд 27С2-цикл или Фотодыхание
2. Дефосфорилирование фосфокликолата;
фермент – фосфогликолат фосфатаза
фосфогликолат + Н2О
Транспортная реакция А: Гликолат переносится через 2 мембраны хлоропласта и одиночную мембрану пероксисомы внутрь пероксисомы;
фермент – кликолат-глицерат транслокатор
3. Окисление гликолата до клиоксилата растворенным кислородом; фермент – гликолат оксидаза
гликолат + О2 → глиоксилат + Н2О2
4. Переаминирование (аминирование глиоксилата за счет дезаминирования глутамата); фермент – глутамат-глиоксилат аминотрансфераза.
глиоксилат + глутамат → глицин + 2-оксоглутарат

Слайд 28С2-цикл или Фотодыхание
Транспортная реакция Б-1:
глицин переносится через одиночную мембрану
фермент – транслокатор аминокислот
5. Декарбоксилирование глицина;
фермент – глицин-декарбоксилазный комплекс
глицин + НАДН2 → серин + НАД+ + СО2 + NH4+
Транспортная реакция Б-2:
серин переносится обратно в пероксисому, проходя через 2 мембраны митохондрии и одиночную мембрану пероксисомы;
фермент – транслокатор аминокислот
6. Дезаминирование серина (он превращается в пируват);
фермент – серин-глиоксилат аминотрансфераза
серин → пируват + NH3 (-NH2, NН4+)

Слайд 29С2-цикл или Фотодыхание
7. Восстановление пирувата (до глицерата);
фермент – пируват-редуктаза
пируват +
Транспортная реакция А-2:
глицерат переносится обратно в хлоропласт, проходя через одиночную мембрану пероксисомы и 2 мембраны хлоропласта;
фермент – транслокатор аминокислот
8. Фосфорилирование глицерата;
ферменты – глицерат-киназа
глицерат + АТФ → 3-фосфоглицерат + АДФ
;фермент – пируват-редуктазапируват + НАДН2 → глицерат +")
Слайд 30С2-цикл или Фотодыхание
9. Детоксификация перекиси водорода;
фермент – каталаза
2 Н2О2 →
Транспортная реакция В:
2-оксоглутарат переносится обратно в хлоропласт, проходя через одиночную мембрану пероксисомы и 2 мембраны хлоропласта;
фермент – малата-глутамат/2-оксо-глутарат транслокатор
10. Фосфорилирование оксоглутарата;
фермент – глутамат синтаза и глутамин-синтета
оксоглутарат + АТФ → глутамат + АДФ

Слайд 32Цикл Хетча – Слэка – Карпилова (С4-цикл)
Цикл Кальвина – основной, но
Так, советский ученый Ю. Карпилов и австралийские ученые М. Хетч и К. Слэк выявили, что у некоторых растений, главным образом тропических и субтропических, таких как кукуруза, сахарный тростник, сорго и другие, основная часть меченного углерода (14СО2) уже после нескольких секунд фотосинтеза обнаруживается не в фосфоглицериновой кислоте, а в щавелево-уксусной (ЩУК), яблочной (ЯК) и аспарагиновой (АК) кислотах.
Цикл Кальвина – основной, но не единственный путь восстановления")
Слайд 33В этих кислотах можно обнаружить в первые секунды до 90 %
Так как эти органические кислоты содержат 4 атома углерода, то такие растения начали называть С4-растениями в отличие от С3-растений, в которых радиоуглеродная метка появляется прежде всего в ФГК.
Это открытие положило начало серии исследований, в результате которых подробно был изучен химизм превращения углерода в фотосинтезе в С4-растениях.






