- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Биофизика мембранных процессов презентация
Содержание
- 1. Биофизика мембранных процессов
- 2. Биомембраны – это функциональные высокоорганизованные структуры
- 3. Функции биомембран (БМ) Барьерная – поддержание автономности
- 4. ИСТОРИЯ МЕМБРАНОЛОГИИ
- 5. Модель строения биологических мембран Даниэли-Доусона, или модель «сэндвича».
- 6. Жидкостно-мозаичная модель биомембран предложена Сингером и Николсоном
- 7. ЖИДКО-МОЗАИЧНАЯ МОДЕЛЬ
- 8. Ультраструктура биологической мембраны.
- 9. Жидкостно-мозаичная модель строения биомембран (БМ) 1.
- 10. Липидный бислой представлен преимущественно молекулами фосфолипидов состоящими
- 11. Недостатки жидкостно-мозаичной модели Не все белки свободно
- 12. Жидкостно-мозаичная модель строения мембран названа «последней классической»
- 13. Метаморфно-мозаичная модель биомембран
- 14. ХАРАКТЕР СВЯЗЕЙ МЕЖДУ КОМПОНЕНТАМИ КЛЕТОЧНЫХ МЕМБРАН гидрофобное
- 15. Мембранные липиды
- 16. Мембранные липиды представлены 3 классами: фосфолипиды, гликосфинголипиды,
- 21. Гликосфинголипиды – гликозильные производные церамида, делятся на
- 23. Стероиды – спирты со стерановым скелетом (холестерин, ситостерин)
- 24. ЛИПИДЫ КЛЕТОЧНЫХ МЕМБРАН структурная формула холестерина схема
- 25. Роль холестерина в биомембранах Регулирует правильную упаковку
- 27. Принципы организации липидного бислоя Амфифильность фосфолипидов. Полиморфизм
- 28. 1.Амфифильность фосфолипидов. Схема строения молекулы фосфолипида, фосфолипидного бислоя и биологической мембраны
- 30. Бислой образован амфифильными молекулами фосфолипидов и сфингомиелина.
- 31. Молекула фосфатидилэтаноламина
- 32. 2.Полиморфизм липидов – способность липидов образовывать различные
- 33. Полиморфизм липидов зависит от формы липида
- 34. Основные типы структурной организации водно-липидных систем 1/2
- 35. 3.Фазовые переходы мембранных липидов
- 36. Точка фазового перехода или температура плавления Это
- 37. В жидко-кристаллической фазе наблюдается конформационное плавление –
- 40. 4.Текучесть (микровязкость) липидной фазы биомембран регулирует мембранные
- 41. Текучесть БМ зависит от: Ненасыщенности мембранных липидов:
- 42. Мембранные липиды – мишень адаптации –
- 43. 5. Латеральная гетерогенность мембранных липидов.
- 44. Мембранные рафты – это
- 45. Перед тем, как снять осаду, нападающие сообщили
- 46. Выделяют 2 типа липидных рафтов: планарные и кавеолы Планарные рафты Кавеолы
- 47. Рафтовые неоднородности в мембране различного масштаба: а
- 48. Взаимодействие лейкоцитов с липидными рафтами
- 49. 6. Различные виды подвижности мембранных липидов 1.
- 50. 7. Трансбислойная асимметрия мембранных липидов – это
- 51. Трансмембранное перемещение фосфолипидов в двойном липидном слое:
- 52. Транслоказы фосфолипидов создают асимметричное распределение мембранных липидов.
- 53. Нарушение асимметрии липидного бислоя индуцирует апоптоз. Транслокация фосфатидилсерина во внешний монослой – рецепторы «съешь меня»
- 54. Выделяют 4 вида липидов по типу взаимодействия
- 55. Мембранные белки
- 56. 20-30% всех белков – мембранные. По своему
- 58. Топологическая классификация мембранных белков: Монотопические – являются
- 60. Выделяют 6 типов интегральных белков.
- 61. Основные типы трансмембранных α-спиральных белков
- 62. Типы связывания монотопических белков с мембраной посредством:
- 63. Типы связывания политопических белков с мембраной:
- 64. Функции мембранных белков Ферментативная Рецепторная Транспортная
- 65. ФУНКЦИИ МЕМБРАННЫХ БЕЛКОВ Рецепторная функция. лиганд активированный белковый рецептор активированная внутриклеточная киназа
- 66. ФУНКЦИИ МЕМБРАННЫХ БЕЛКОВ Транспортная функция. Простая и
- 67. ГЛИКОКАЛИКС слой углеводов цитоплазма ядро плазматическая мембрана Электронная микрофотография
- 68. ГЛИКОКАЛИКС бислой липидов слой углеводов трансмембранный гликопротеин Схема организации поверхностный гликопротеин гликолипид трансмембранный протеогликан углевод
- 69. Образование мембранных структур. Создание искусственных бислойных мембран
- 70. Искусственная плоская липидная мембрана ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 71. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Бислой на кончике пипетки.
- 72. Схема ванны Ленгмюра ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 73. Образование бислойной липидной мембраны. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 74. Липосомы. Образование и виды. ФГОУ ВПО
- 75. Способы включения различных веществ в липосомы. ФГОУ
- 76. Применение липосом. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 77. Липосомы-невидимки. ФГОУ ВПО ЮФУ каф. биохимии
- 78. Электрокинетические явления
- 79. Электрокинетические явления (ЭКЯ) - это
- 80. Дисперсные системы (ДС) – гетерогенные системы,
- 81. ЭКЯ характерны для систем, в которых
- 82. Электрофорез и электроосмос Открыты в 1807 г.
- 84. Потенциалы течения и седиментации Потенциалы
- 87. Все ЭКЯ
- 88. Механизм возникновения ζ-потенциала Диссоциация ионогенных групп мембраны,
- 89. 5. В итоге вокруг частицы или клетки
- 91. Разность потенциалов, которая возникает на
- 92. ζ-потенциал измеряется с помощью электрофореза. Формула Смолуховского
- 93. Электрофорез в агарозном геле с использованием бромистого этидия для
- 94. Полиморфные фазы и соответствующие молекулярные формы липидов:

Слайд 2
Биомембраны – это функциональные высокоорганизованные структуры клеток, ограничивающие цитоплазму и большинство

Слайд 3Функции биомембран (БМ)
Барьерная – поддержание автономности и компартментализации клетки
Матриксная – БМ-
- Внутренняя мембрана митохондрий содержит ЭТЦ и участвует в синтезе АТФ
- Мембраны тилакоидов в хлоропластах участвуют в фотосинтезе
- Мембраны ЭПР участвует в: а)синтезе белков, липидов; б)детоксикации ксенобиотиков
Транспортная
Рецепторная, участие в клеточной сигнализации
Межклеточная коммуникация (адгезия, щелевые контакты)
Электрическая возбудимость и проведение нервного импульса
Определяет гормональный и иммунный ответы
Участие в запуске апоптоза (митохондриальная и плазматическая мембраны)
Фоторецепторные мембраны участвуют в зрительном восприятии
Нарушение стабильности и структуры БМ – основа многих патологий (мембранная патология).
Барьерная – поддержание автономности и компартментализации клеткиМатриксная – БМ- матрица для протекания важнейших")


Слайд 6Жидкостно-мозаичная модель биомембран предложена Сингером и Николсоном в 1972 г. Мембрана

Слайд 7ЖИДКО-МОЗАИЧНАЯ МОДЕЛЬ
липиды
углеводы
гликопротеины
интегральные белки
полуинтегральные белки
поверхностные белки
цитоплазма клетки
внеклеточная жидкость
цитоскелет
гликолипиды


Слайд 9Жидкостно-мозаичная модель строения биомембран (БМ)
1. БМ различного происхождения имеют общий план
2. БМ - ансамбли липидов и белков, которые удерживаются вместе за счет нековалентных (слабых) взаимодействий, важнейшие из них - гидрофобные взаимодействия. Минорные компоненты мембран - гликолипиды и гликопротеины. Имеется мембранная вода.
3. Липиды образуют двойной липидный слой, который находится в жидко-кристаллическом состоянии.
4. Белки встроены в липидный матрикс подобно элементам мозаики и могут свободно перемещаться в мембране.
5. Липидам отводится пассивная роль, основная роль принадлежит белкам.
6. Наблюдается трансмембранная асимметрия.
1. БМ различного происхождения имеют общий план строения.2. БМ - ансамбли")
Слайд 10Липидный бислой представлен преимущественно молекулами фосфолипидов состоящими из двух длинных неполярных
В мембране гидрофобные цепи обращены внутрь бислоя, а гидрофильные головки – кнаружи.
 цепей жирных кислот")
Слайд 11Недостатки жидкостно-мозаичной модели
Не все белки свободно диффундируют в жидком липидном бислое.
Имеются
Наблюдается неоднородность мембраны за счет явления латеральной гетерогенности бислоя (рафты).
4. Мембранные липиды имеют не меньшее значение, чем мембранные белки.
Разрабатывается метаморфно-мозаичная и рафтовая модели БМ

Слайд 12Жидкостно-мозаичная модель строения мембран названа «последней классической» теорией потому что, с


Слайд 14ХАРАКТЕР СВЯЗЕЙ МЕЖДУ КОМПОНЕНТАМИ КЛЕТОЧНЫХ МЕМБРАН
гидрофобное взаимодействие
ионные (солевые) силы взаимодействия
ионные (солевые)
ионные (солевые) силы взаимодействия
ковалентные силы взаимодействия
 силы взаимодействияионные (солевые) силы взаимодействияионные (солевые) силы")

Слайд 16Мембранные липиды представлены 3 классами: фосфолипиды, гликосфинголипиды, стероиды
Глицерофосфолипиды – производные фосфа-тидной кислоты, к гидроксилу фосфорной кислоты которой с помощью простой эфирной связи присоединен радикал Х. (фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозитол).
Сфингофосфолипиды – производные церами-да (жирнокислотного эфира ненасыщенного аминоспирта сфингозина) и монофосфорных эфиров спиртов (сфингомиелин).

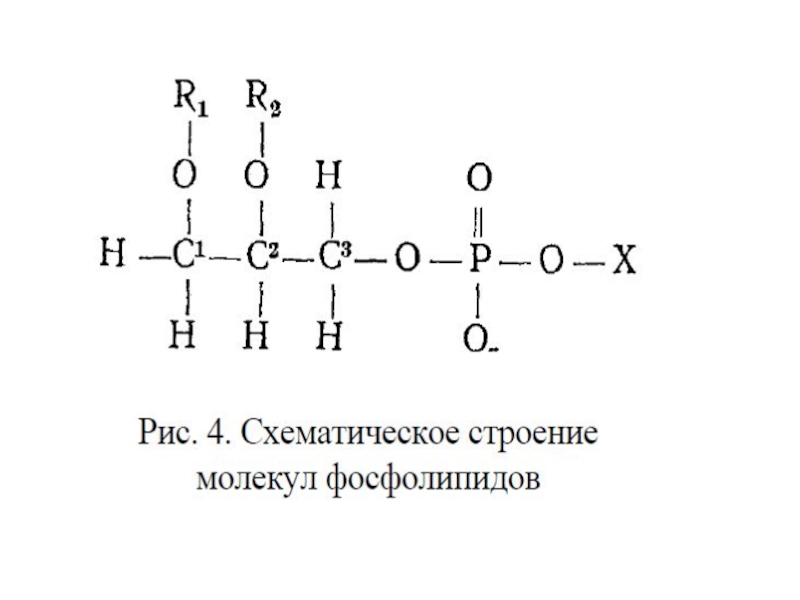

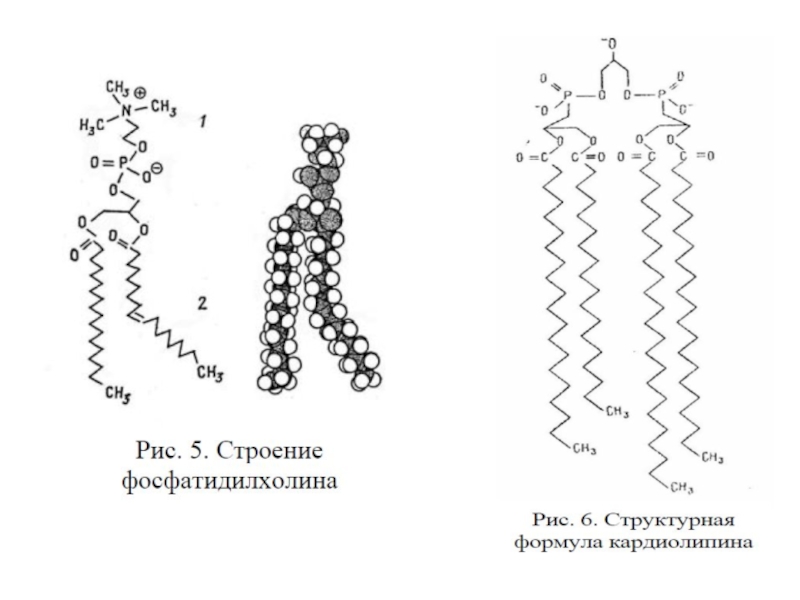
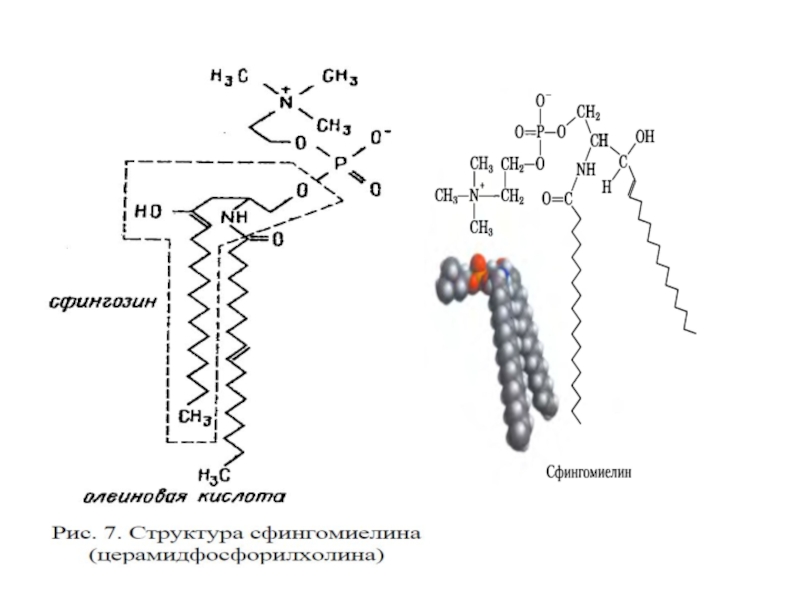
Слайд 21Гликосфинголипиды – гликозильные производные церамида, делятся на группы:
1.Цереброзиды – к ОН-группе
2. Ганглиозиды – к ОН-группе церамида присое-динен сложный, разветвленный олигосахарид, N-ацетилнейраминовая кислота.
3. Сульфатиды – сульфопроизводные гликолипидов

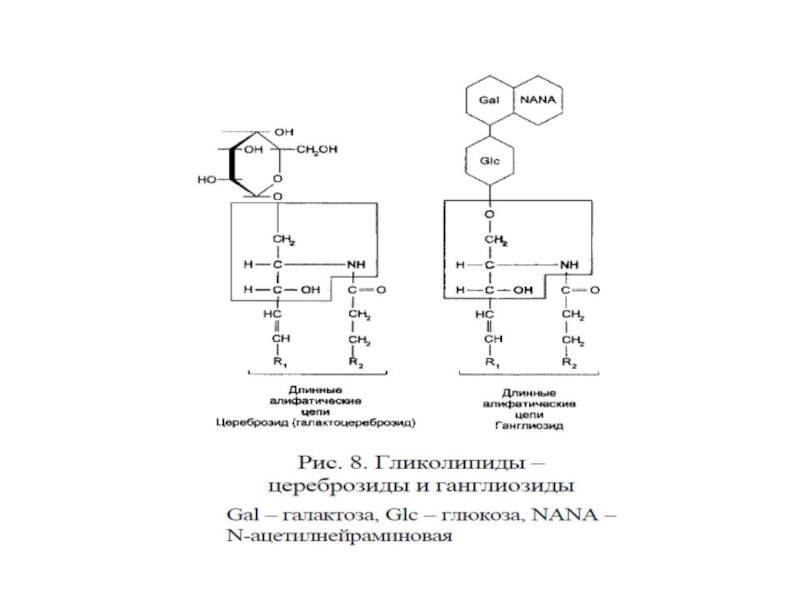
")
Слайд 24ЛИПИДЫ КЛЕТОЧНЫХ МЕМБРАН
структурная формула холестерина
схема строения холестерина
гидрофильная головка
гидрофобный хвост
жесткое
стероидное
кольцо

Слайд 25Роль холестерина в биомембранах
Регулирует правильную упаковку мембранных липидов:
-если мембрана слишком «жидкая» (текучая), то вызывает ее уплотнение.
2. Взаимная укладка ФЛ и Хол: неполярные цепи ФЛ вытянуты, а его полярная головка изогнута в виде трости. В образующуюся полость помещается Хол. Другой способ укладки – образование рафтов.
3. Локализован преимущественно в плазматических мембранах.

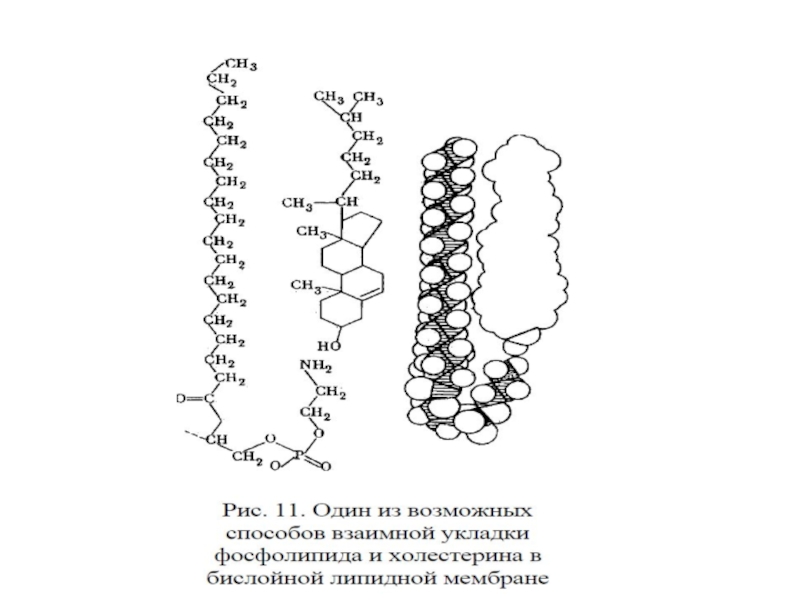
Слайд 27Принципы организации липидного бислоя
Амфифильность фосфолипидов.
Полиморфизм липидов – способность образо-вывать агрегаты различных
Фазовые переходы мембранных липидов.
Текучесть (микровязкость) биомембран.
Латеральная гетерогенность мембранных липидов.
Подвижность мембранных липидов.
Трансбислойная асимметрия липидов.

Слайд 281.Амфифильность фосфолипидов.
Схема строения молекулы фосфолипида, фосфолипидного бислоя и биологической мембраны

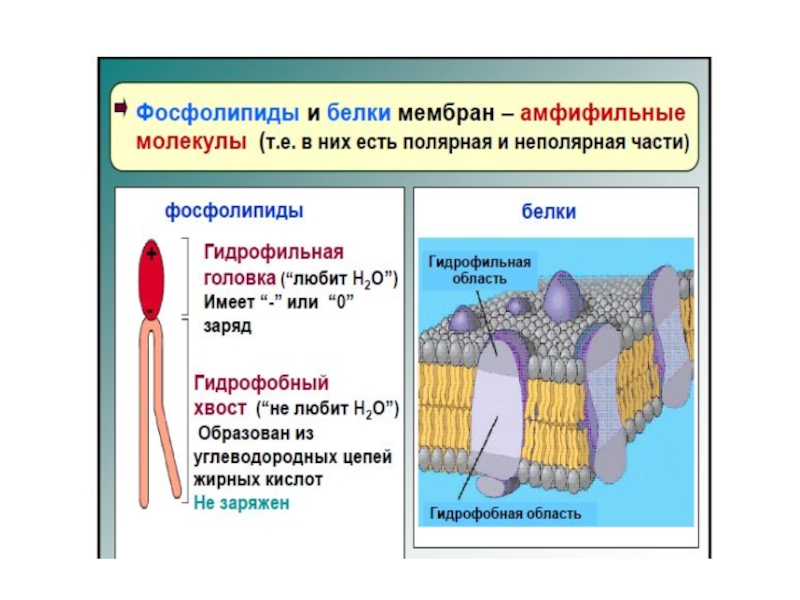
Слайд 30Бислой образован амфифильными молекулами фосфолипидов и сфингомиелина.
Амфифильными эти молекулы называются потому,
гидрофильной – полярной «головки» (1/4 объема), имеющей высокое сродство к воде;
гидрофобной (3/4 объема) – «хвостов», образованных неполярными углеводородными цепями жирных кислот, обладающих низким сродством к воде.


Слайд 322.Полиморфизм липидов – способность липидов образовывать различные структуры под влиянием факторов



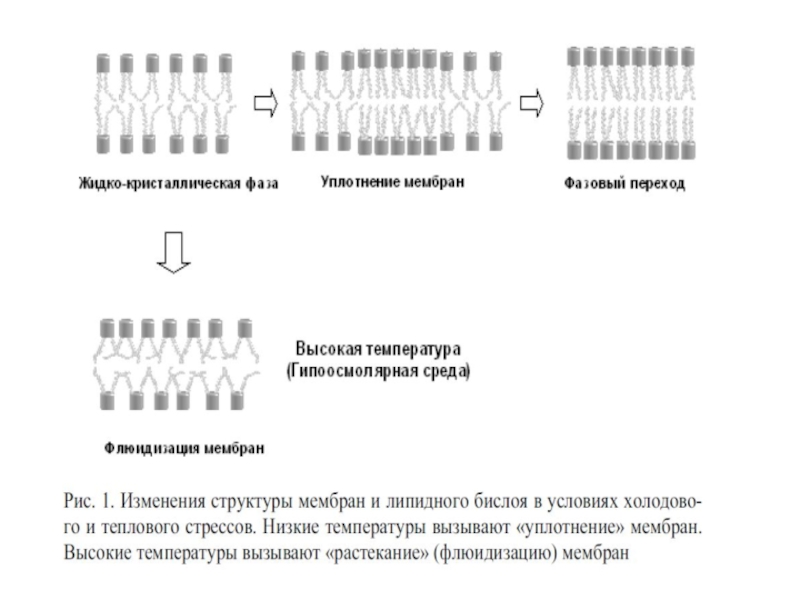
Слайд 353.Фазовые переходы мембранных липидов
Мембранные липиды в бислое
- твердо-кристаллической So (solid ordered) (гель): толщина бислоя максимальна, площадь на молекулу липида минимальна;
- жидко-кристаллической L (liquid), которая делится на 2 фазы (толщина бислоя минимальна, площадь на молекулу липида максимальна):
а) жидкая упорядоченная Lo (liquid ordered): высокая упорядоченность жирнокислотных ацилов, но высокая латеральная подвижность липидных доменов (мезофаза);
б) жидкая неупорядоченная Ld (liquid disordered): липидные хвосты максимально неупорядочены.

Слайд 36Точка фазового перехода или температура плавления
Это такая температура, при которой 50%
У теплокровных животных и человека точка фазового перехода лежит в области отрицательных температур

Слайд 37В жидко-кристаллической фазе наблюдается конформационное плавление – транс-конформация ацилов жирных кислот

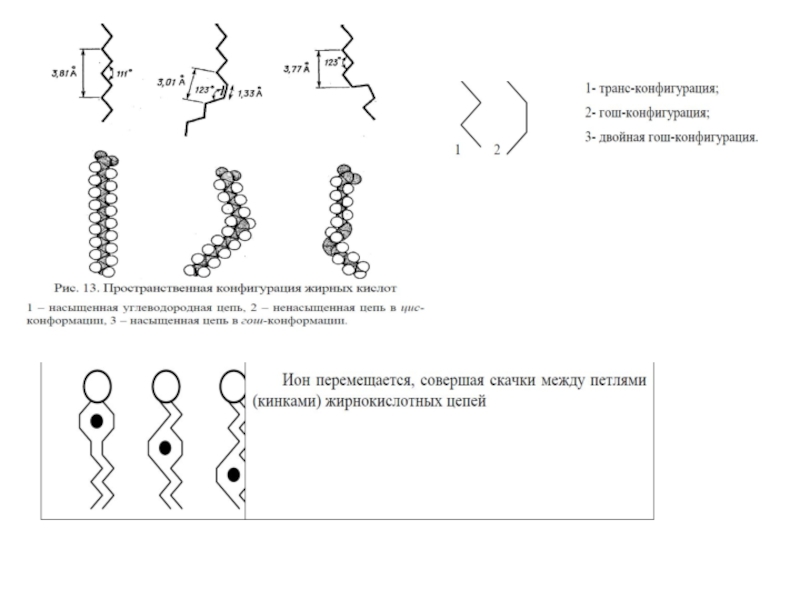
Слайд 404.Текучесть (микровязкость) липидной фазы биомембран регулирует мембранные процессы.
В физиологических условиях липиды
Степень текучести мембран зависит от:
- длины жирнокислотных цепей и степени их ненасыщенности;
- соотношения фосфолипиды/стерины;
- взаимодействия между ФЛ и белками;
- физических факторов (to , р, рН, потенциала)
 липидной фазы биомембран регулирует мембранные процессы.В физиологических условиях липиды мембран имеют жидко-кристаллическую структуру,")
Слайд 41Текучесть БМ зависит от:
Ненасыщенности мембранных липидов:
увеличение количества двойных связей
повышает текучесть
2. Длины ЖК-цепи: удлинение ацильной цепи уменьшает текучесть мембраны.
3. Температуры среды: увеличение температуры повышает текучесть мембраны

Слайд 42
Мембранные липиды – мишень адаптации – повышение ненасыщенности жирнокислотных цепей мембранных

Слайд 435. Латеральная гетерогенность мембранных липидов. Латеральная гетерогенность мембраны – это свойство

Слайд 44 Мембранные рафты – это маленькие (10-200 нм), гетерогенные
Биологическая роль рафтов
1.Группировка и сортировка белков для выполнения функций.
2.Деление клеток.
3.Везикулярный транспорт.
4.Способствуют проникновению вирусов в клетку и выходу из клетки.
5.Способноствуют проникновению токсинов в клетку.
, гетерогенные липидные домены, обогащенные холестеролом, сфинголипидами,")
Слайд 45Перед тем, как снять осаду, нападающие сообщили троянцам, что построенный ими
Рафты наряду с важной биологической ролью в определенных условиях становятся «троянским конем» для клетки


Слайд 47Рафтовые неоднородности в мембране различного масштаба: а – нанокластер, б –


Слайд 496. Различные виды подвижности мембранных липидов
1. Изменение ориентации полярных голов.
2. Латеральная
3. Сегментарная подвижность – колебания ацильных цепей.
4. Образование кинков и их миграция вдоль ацильных цепей.
5. Ротационная подвижность.
6. Флип-флоп переходы.
7. Выход из бислоя.

Слайд 507. Трансбислойная асимметрия мембранных липидов – это различное распределение липидов между
Во внешнем монослое локализуются холинсодержащие ФЛ – фосфатидилхолин, сфингомиелин, а также стероид холестерол.
Во внутреннем монослое локализуются аминосодержащие ФЛ – фосфатидилэтаноламин, фосфатидилсерин.
Во внешнем монослое – 1/3 двойных связей, во внутреннем – 2/3 двойных связей.
Известны 2 механизма, обеспечивающие асимметрию липидного бислоя:
- различия конформационных размеров полярных «головок» липидов;
- транслокация с помощью белков-переносчиков.

Слайд 51Трансмембранное перемещение фосфолипидов в двойном липидном слое:
Некатализируемый флип-флоп из одного монослоя
Катализируемый перенос фосфолипидов в мембране:
1.Флиппазы (АТФ-азы Р-типа) переносят аминофосфо-липиды (ФС, ФЭА) от внешнего к внутреннему монослою;
2. Флоппазы (АВС-транспортеры) переносят фосфолипиды от цитозольного монослоя к внешнему монослою (используют АТФ);
3.Скрамблазы выравнивают фосфолипидный состав двух монослоев, нарушают асимметрию (не используют АТФ, активируются Са2+).

Слайд 52Транслоказы фосфолипидов создают асимметричное распределение мембранных липидов. АТФ-зависимые специфические переносчики –

Слайд 53Нарушение асимметрии липидного бислоя индуцирует апоптоз. Транслокация фосфатидилсерина во внешний монослой

Слайд 54Выделяют 4 вида липидов по типу взаимодействия с белками
1.Аннулярные
2. «Неаннулярные липиды» - прочно связаны с белком и играют роль кофакторов
3. Липиды общего липидного бислоя или свободного объема (часто находятся в жидко-неупорядоченном состоянии )
4.Рафтовые липиды – локализованы в рафтах и находятся в жидко-упорядоченном состоянии


Слайд 5620-30% всех белков – мембранные. По своему расположению относительно липидного бислоя
1) Периферические -
непрочно связаны с поверхностью мембраны и обычно находятся вне липидного бислоя.
2) Интегральные -
3) Амфитропные - белки с переменной гидрофобностью, могут находиться в цитозоле и бислое.
очень прочно связаны с мембраной и делятся на:
1.собственно интегральные белки – полностью погружены в липидный бислой; 2.полуинтегральные белки -частично погружены в бислой.

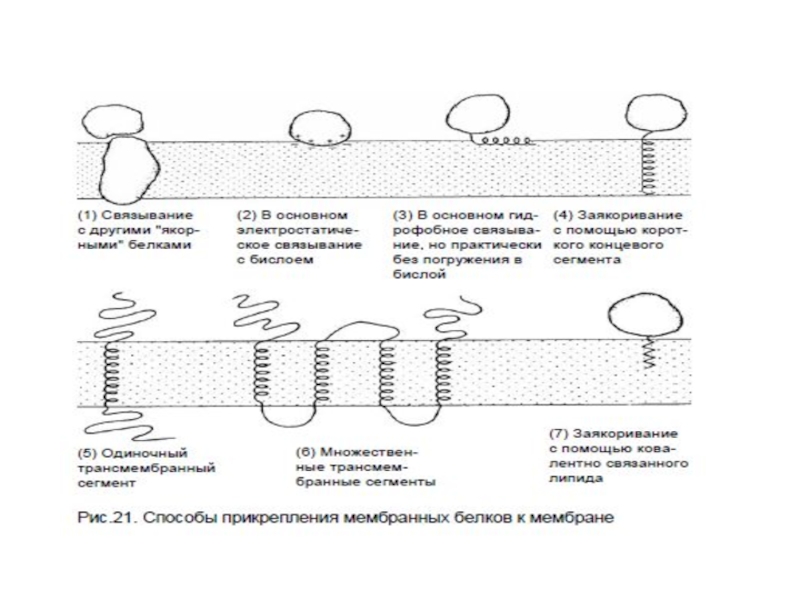
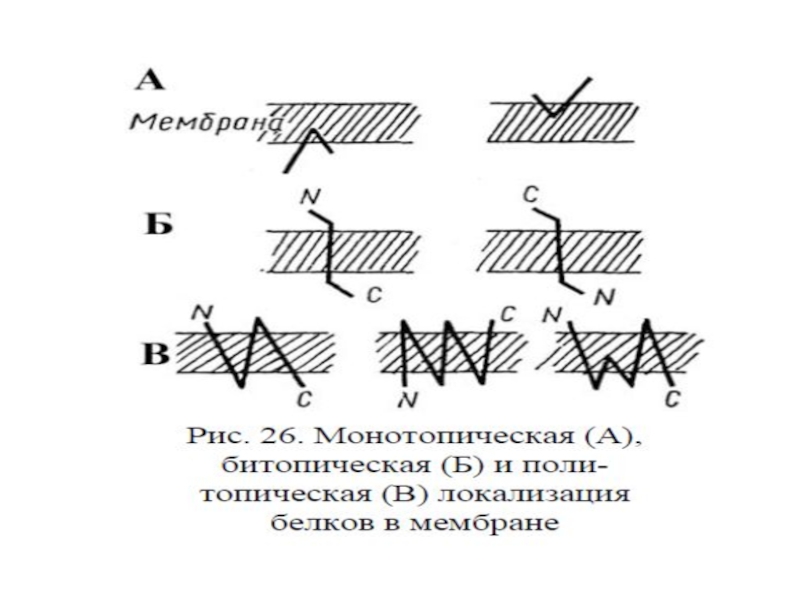
Слайд 58Топологическая классификация мембранных белков:
Монотопические – являются периферическими белками и не пронизывают
2. Битопические – пронизывают мембрану один раз (гликофорин)
3. Политопические - пронизывают мембрану два и более раз (рецепторы, сопряженные с G-белками; Na, K- АТФ-аза; АТФ-синтаза и др.)
2. Битопические")
Слайд 60Выделяют 6 типов интегральных белков.
I тип – один трансмембранный
II тип – одна трансмембранная спираль, N-конец внутри
Ш тип – много трансмембранных спиралей IV тип – трансмембранные домены нескольких полипеп-тидов соединяются с образованием канала через мембрану V тип – белки удерживаются бислоем с помощью ковалентно связанных липидов VI тип – содержат трансмембранные спирали и липидные ГФИ-якори
(ГФИ – гликозилфосфатидилинозитол)


Слайд 62Типы связывания монотопических белков с мембраной посредством:
1 – амфипатической α-спирали,
2 – гидрофобной петли,
3 – ковалентно-присоединенного остатка жирной кислоты,
4 – электростатического взаимодействия (прямого или металл-опосредованного)

Слайд 63Типы связывания политопических белков с мембраной:
1- связывание посредством единичной α-спирали,
2 – посредством множественных α-спиралей,
3 – посредством складчатой β-структуры

Слайд 64Функции мембранных белков
Ферментативная
Рецепторная
Транспортная
- образуют каналы
Межклеточная коммуникация – белки межклеточной адгезии
Структурная

Слайд 65ФУНКЦИИ МЕМБРАННЫХ БЕЛКОВ
Рецепторная функция.
лиганд
активированный
белковый рецептор
активированная внутриклеточная киназа

Слайд 66ФУНКЦИИ МЕМБРАННЫХ БЕЛКОВ
Транспортная функция. Простая и облегченная диффузия. Активный транспорт с


Слайд 68ГЛИКОКАЛИКС
бислой
липидов
слой
углеводов
трансмембранный гликопротеин
Схема организации
поверхностный гликопротеин
гликолипид
трансмембранный протеогликан
углевод

Слайд 69Образование мембранных структур.
Создание искусственных бислойных мембран позволяет моделировать и исследовать свойства
Сферические бислойные мембраны
Плоские бислойные мембраны
Электроды
Тефлоновый стакан
Круглое отверстие в стенке тефлонового стакана





Слайд 74Липосомы. Образование и виды.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Стремление фосфолипидов
Различные виды липосом:
мультиламеллярные везикулы (МЛВ).
большие моноламеллярные везикулы (БМВ).
олиголамеллярные везикулы (ОЛВ).
олиговезикулярные липосомы (ОВЛ).
малые моноламеллярные везикулы (ММВ).
дискомы - дискоообразные липосомы.
тубулярные трубчатые везикулы

Слайд 75Способы включения различных веществ в липосомы.
ФГОУ ВПО ЮФУ каф. биохимии и
Водорастворимые вещества включаются во
внутренний водный объём липосом. Наличие
в бислое достаточно протяжённой
углеводородной области позволяет вводить в него гидрофобные молекулы. На поверхности бислоя можно адсорбировать различные вещества, а также химически связывать их с липидами или другими компонентами мембраны.


Слайд 77Липосомы-невидимки.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Полиэтиленгликоль с молекулярной массой 1500-5000


Слайд 79 Электрокинетические явления (ЭКЯ) - это движение фаз дисперсной системы
 - это движение фаз дисперсной системы под влиянием наложенного на нее")
Слайд 80Дисперсные системы (ДС) –
гетерогенные системы, состоящие из 2 фаз: дисперсионной
Характерное свойство ДС – наличие межфазной поверхности, свойства которой влияют на ДС.
Дисперсионная среда крови – плазма, дисперсные фазы – форменные элементы крови (эритроциты, лейкоциты, тромбоциты)
 – гетерогенные системы, состоящие из 2 фаз: дисперсионной среды – растворителя, и")
Слайд 81 ЭКЯ характерны для систем, в которых на границе раздела фаз
Открыто 4 вида ЭКЯ:
электрофорез потенциалы оседания (седиментации)
электроосмос потенциалы течения

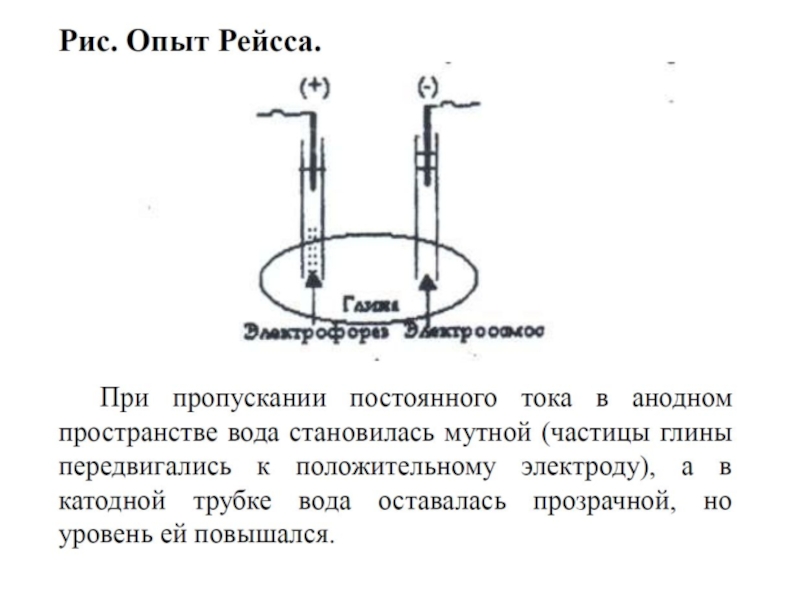
Слайд 82Электрофорез и электроосмос
Открыты в 1807 г. проф. Ф.Ф.Рейссом.
2.
3. Электроосмос – движение дисперсионной среды в постоянном электрическом поле к противоположно заряженному электроду.
4. Рейсс проводил опыт: в сырую глину помещались 2 трубки, в которые наливалась вода и опускались электроды. При наличии капиллярно-пористой перегородки (в опыте Рейсса – это влажная глина) под действием постоянного электрического поля передвигаются дисперсионная среда (электроосмос) и дисперсная фаза (электрофорез).

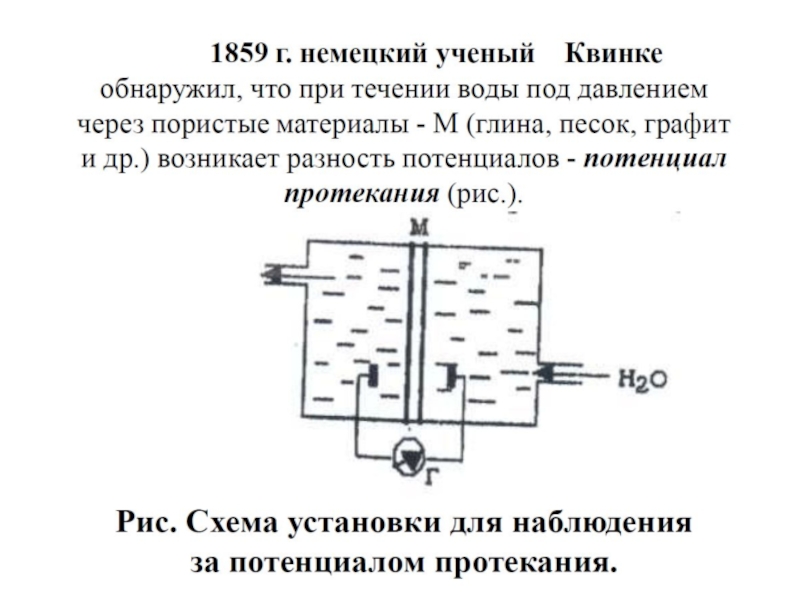
Слайд 84Потенциалы течения и седиментации
Потенциалы течения (Uf ) (Квинке,1859) –
Потенциалы седиментации (Us) (Дорн, 1878) – возникают в дисперсной системе, когда частицы твердой фазы, несущие заряд, осаждаются под действием силы тяжести. В результате между твердой и жидкой фазами возникнет разность потенциалов. Это явление противоположно электрофорезу.
 (Квинке,1859) – возникают в результате движения дисперсионной")

Слайд 87 Все ЭКЯ связаны с возникновением разности
Причина появления ζ- потенциала – возникновение поверхностного заряда на границе раздела двух фаз в результате асимметричного распределения ионов.

Слайд 88Механизм возникновения ζ-потенциала
Диссоциация ионогенных групп мембраны, частицы.
Адсорбция ионов и полярных молекул
В итоге ионы одного знака прочно связаны с дисперсной фазой и формируют потенциалообразующий слой, а ионы противоположного знака находятся в дисперсионной среде, образуя адсорбционный слой.
Эти два слоя образуют двойной электрический слой (ДЭС), который формируется под действием 2 противоположно действующих сил – электростатического притяжения и теплового движения ионов.

Слайд 895. В итоге вокруг частицы или клетки образуется ионная атмосфера с
φ = φ0 е-рх , где
φ0 - полный электрохимический потенциал
φ - электрохимический потенциал на расстоянии х
р – коэффициент, характеризующий свойства среды
Дебаевская длина - расстояние х, на котором φ падает в е раз (φ=0,37 φ0)
6. Толщина ДЭС находится в обратной зависимости от ионной силы раствора.

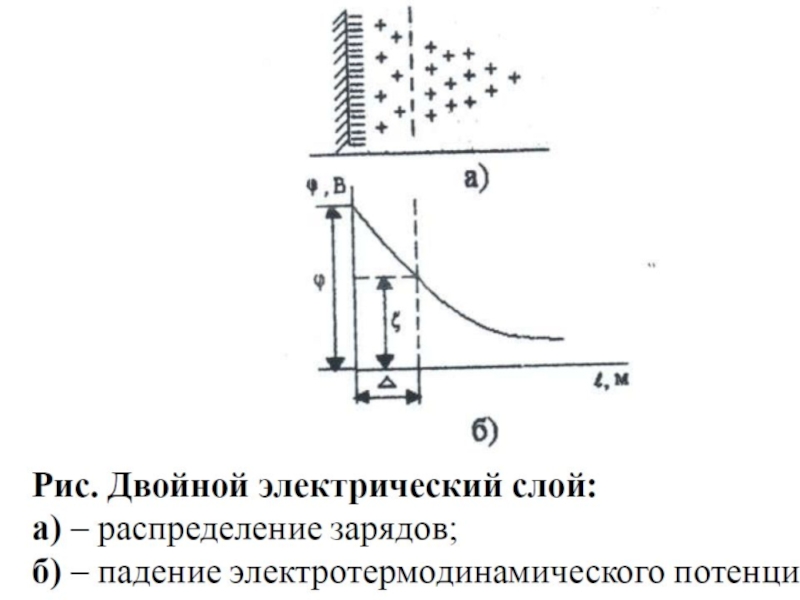
Слайд 91 Разность потенциалов, которая возникает на границе между тончайшей пленкой
ζ-потенциалом.
Его величина находится в прямой зависимости от толщины ДЭС и в обратной зависимости от ионной силы раствора (дисперсионной среды).
 -")
Слайд 92ζ-потенциал измеряется с помощью электрофореза.
Формула Смолуховского для определения ζ-потенциала:
V - скорость движения частицы в электрическом поле
E - напряженность электрического поля
ζ - дзета-потенциал
ε - диэлектрическая постоянная среды
η - вязкость среды
ζ = 4πηV/Eε

Слайд 93Электрофорез в агарозном геле с использованием бромистого этидия для визуализации результатов в ультрафиолете (слева).
. Вторая слева дорожка — маркер")
Слайд 94Полиморфные фазы и соответствующие молекулярные формы липидов:
а – плоский бислой и






