проблем информатики РАН
sgrinchenko@ipiran.ru
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
О ДАРВИНИЗМЕ И НОМОГЕНЕЗЕ: КИБЕРНЕТИЧЕСКИЙ СИНТЕЗ презентация
Содержание
- 1. О ДАРВИНИЗМЕ И НОМОГЕНЕЗЕ: КИБЕРНЕТИЧЕСКИЙ СИНТЕЗ
- 2. План доклада 1. О соотношении теории эволюции
- 3. Кибернетическая (теоретико-управленческая) трактовка функционирования и эволюции живой
- 4. «В целом регулирующий механизм эволюции популяции [Шмальгаузен
- 5. Основной причиной весьма незначительного вклада кибернетики
- 6. К концу 60-х гг. ситуация в самой
- 7. Адаптивная поисковая оптимизация воспроизводит/моделирует активное непрерывное
- 8. Современный развитый информатико-кибернетический язык оперирует не только
- 9. «Механизмы случайного поиска, по-видимому, свойственны природе нашего
- 10. Предлагаемый поисково-оптимизационный подход предполагает рассмотрение биологических систем
- 11. Легко показать, что минимизация критерия
- 12. «Нынешняя структура есть следствие вчерашней кинетики» [А.М.Молчанов,1967].
- 13. где:
- 14. Введем понятие метаэволюции как процесса последовательного наращивания
- 15. Понятие «метаэволюция» необходимо выделить из традиционно использующегося
- 16. Живая природа сформировала на Земле, на 12-ти
- 21. Продолжительность каждой триады (согласно эмпирическим данным) –
- 23. В иерархии системы живого процесс поисковой оптимизации
- 24. Микроэволюция, протекающая внутри конкретного вида, является частью
- 25. Основными средствами реализации поисково-оптимизационного процесса адаптации/эволюции живого
- 26. Согласно энергическому критерию поисково-оптимизационного развития системы живого,
- 27. Реконструкция механизма биологической эволюции по Ч.Дарвину. Места
- 28. 1 – имманентная активность, т.е.
- 29. Предложенная концепция позволяет снять многолетнее недоразумение –
- 30. «Необходимо отметить, что в биологии термин "эволюция"
- 31. «Естественный отбор бесспорно остаётся движущим фактором эволюции,
- 32. Резюме 1. Моделируется иерархическая система живого, поведение
- 33. Гринченко С.Н. Системная память живого (как основа
Слайд 1О ДАРВИНИЗМЕ И НОМОГЕНЕЗЕ: КИБЕРНЕТИЧЕСКИЙ СИНТЕЗ
Гринченко Сергей Николаевич
д.т.н., проф., гл.н.с.
Институт

Слайд 2План доклада
1. О соотношении теории эволюции и теории управления (2 слайда).
2.
Современный кибернетический аппарат моделирования природных систем (5 слайдов).
3. Иерархическая модель системы живого (4 слайда).
4. Метаэволюция системы живого (9 слайдов).
5. Дарвинизм и номогенез в терминах поведения механизма адаптивной поисковой оптимизации энергетики в системе живого (10 слайдов).
3. Иерархическая модель системы живого (4 слайда).
4. Метаэволюция системы живого (9 слайдов).
5. Дарвинизм и номогенез в терминах поведения механизма адаптивной поисковой оптимизации энергетики в системе живого (10 слайдов).
.2. Современный кибернетический аппарат моделирования")
Слайд 3Кибернетическая (теоретико-управленческая) трактовка функционирования и эволюции живой природы была дана основоположниками
на заре кибернетики.
«Способность к авторегуляции жизненных проявлений – наиболее характерная особенность всех живых существ, причем регуляторные системы организмов – наиболее сложные, наиболее совершенные кибернетические устройства» [академик АН СССР И.И.Шмальгаузен (1884-1963). Кибернетические вопросы биологии. Новосибирск: Наука, 1968, С.21].
«Управление можно объявить характеристическим свойством жизни в широком смысле» [член-корр. АН СССР А.А.Ляпунов (1911-1973). Проблемы теоретической и прикладной кибернетики. М.: Наука, 1980, С.208].
«Способность к авторегуляции жизненных проявлений – наиболее характерная особенность всех живых существ, причем регуляторные системы организмов – наиболее сложные, наиболее совершенные кибернетические устройства» [академик АН СССР И.И.Шмальгаузен (1884-1963). Кибернетические вопросы биологии. Новосибирск: Наука, 1968, С.21].
«Управление можно объявить характеристическим свойством жизни в широком смысле» [член-корр. АН СССР А.А.Ляпунов (1911-1973). Проблемы теоретической и прикладной кибернетики. М.: Наука, 1980, С.208].
 трактовка функционирования и эволюции живой природы была дана основоположниками на заре кибернетики.«Способность к")
Слайд 4«В целом регулирующий механизм эволюции популяции [Шмальгаузен И.И. Регулирующие механизмы эволюции
// Зоол. ж., 1958, том 37, С.9] может быть представлен в виде следующего цикла:
Этот "кибернетический" цикл является лишь перефразировкой дарвиновского понимания эволюции» ([Шмальгаузен И.И. Кибернетические вопросы биологии. Новосибирск: Наука, 1968, С. 177]).

Слайд 5
Основной причиной весьма незначительного вклада кибернетики в теорию биоэволюции к началу
60-х гг. является недостаточная разработанность к этому времени базиса самой кибернетики.
Впрочем, идея отрицательной обратной связи в контуре управления, реализующей устойчивость этого процесса, широко использовалась при моделировании биообъектов.
Этого нельзя сказать об идее положительной обратной связи, которая моделирует процессы взрывного развития. Взрыв и эволюционное развитие имеют мало общего, что и привело к выводу о недостаточности самого кибернетического подхода в данной предметной области.
Впрочем, идея отрицательной обратной связи в контуре управления, реализующей устойчивость этого процесса, широко использовалась при моделировании биообъектов.
Этого нельзя сказать об идее положительной обратной связи, которая моделирует процессы взрывного развития. Взрыв и эволюционное развитие имеют мало общего, что и привело к выводу о недостаточности самого кибернетического подхода в данной предметной области.

Слайд 6К концу 60-х гг. ситуация в самой кибернетике (теории управления) существенно
изменилась: в её рамках были развиты существенно более эффективные и универсальные механизмы самоуправления.
В том числе и те, которые опираются и на отрицательную, и на положительную обратную связь (попеременно).
Появился аппарат адаптивной поисковой оптимизации, включающий аппарат случайно-регулярного, или рандомизированного, поиска. Этот аппарат получил широкое распространение в области технической кибернетики, – и, кстати, в промышленности, – но, к сожалению, остался практически неизвестным в биокибернетике.
Появился аппарат адаптивной поисковой оптимизации, включающий аппарат случайно-регулярного, или рандомизированного, поиска. Этот аппарат получил широкое распространение в области технической кибернетики, – и, кстати, в промышленности, – но, к сожалению, остался практически неизвестным в биокибернетике.
 существенно изменилась: в её рамках")
Слайд 7
Адаптивная поисковая оптимизация воспроизводит/моделирует активное непрерывное стремление основных компонентов биосистемы к
наиболее энергетически предпочтительным для них состояниям.
Адаптивная поисковая оптимизация – процесс движения к цели.
Адаптивная поисковая оптимизация – процесс движения к цели.

Слайд 8Современный развитый информатико-кибернетический язык оперирует не только простейшими понятиями кибернетики («обратная
связь», «контур управления», «вход», «выход», «черный ящик» и т.п.), но и более агрегированными понятиями («поисковая оптимизация», «целевой критерий поисковой оптимизации», «алгоритм адаптивного случайно-регулярного поиска» и др.).

Слайд 9«Механизмы случайного поиска, по-видимому, свойственны природе нашего мира на всех уровнях
его проявления и организации. И, во всяком случае, могут служить удобной и конструктивной моделью этих процессов» – д.т.н., проф. Л.А.Растригин
[Случайный поиск. М.: Знание, 1979].
«По-видимому, всю историю развития жизни на Земле можно было бы изложить на языке многокритериальной оптимизации» – академик АН СССР/РАН Н.Н.Моисеев [Алгоритмы развития. М.: Наука,1987].
«По-видимому, всю историю развития жизни на Земле можно было бы изложить на языке многокритериальной оптимизации» – академик АН СССР/РАН Н.Н.Моисеев [Алгоритмы развития. М.: Наука,1987].

Слайд 10Предлагаемый поисково-оптимизационный подход предполагает рассмотрение биологических систем как иерархических, активных, адаптивных
и целенаправленных. При этом важнейшие их цели имеют энергетический характер:
где: – поток энергии, затрачиваемой на обеспечение специфической функциональной активности биообъекта; – поток дополнительной энергии, вырабатываемой во время переходных процессов в биообъекте и затрачиваемой на перестройку (оптимизацию) его внутренней структуры.
где: – поток энергии, затрачиваемой на обеспечение специфической функциональной активности биообъекта; – поток дополнительной энергии, вырабатываемой во время переходных процессов в биообъекте и затрачиваемой на перестройку (оптимизацию) его внутренней структуры.

Слайд 11Легко показать, что минимизация критерия эквивалентна максимизации «к.п.д.»
системы:
Подобный «энергетический» подход – применительно к организменному и биогеоценотическому ярусам в иерархии живого – активно развивают д.т.н., проф. М.А.Ханин, д. б. н., проф. Н.С.Печуркин, академик РАН А.С.Бугаев и др.
Подобный «энергетический» подход – применительно к организменному и биогеоценотическому ярусам в иерархии живого – активно развивают д.т.н., проф. М.А.Ханин, д. б. н., проф. Н.С.Печуркин, академик РАН А.С.Бугаев и др.



Слайд 14Введем понятие метаэволюции как процесса последовательного наращивания числа уровней/ярусов в иерархической
природной системе.
Этот термин является фактически обобщением предложенного В.Ф.Турчиным термина «метасистемный переход» [Турчин В.Ф. Феномен науки. Кибернетический подход к эволюции. М.: ЭТС, 2000 (1977)] – поскольку метаэволюция подразумевает последовательность таких метасистемных переходов.
Этот термин является фактически обобщением предложенного В.Ф.Турчиным термина «метасистемный переход» [Турчин В.Ф. Феномен науки. Кибернетический подход к эволюции. М.: ЭТС, 2000 (1977)] – поскольку метаэволюция подразумевает последовательность таких метасистемных переходов.

Слайд 15Понятие «метаэволюция» необходимо выделить из традиционно использующегося понятия «эволюция», которое предлагается относить
лишь к приспособительным процессам, происходящим на уже сформировавшихся «верхних» ярусах в системной иерархии.
Метаэволюция же приспособительным процессом не является, и развивается по своим собственным законам (частью специфическим, частью общим с законами метаэволюции неживой природы и метаэволюции социально-технологической природы).
Метаэволюция же приспособительным процессом не является, и развивается по своим собственным законам (частью специфическим, частью общим с законами метаэволюции неживой природы и метаэволюции социально-технологической природы).

Слайд 16Живая природа сформировала на Земле, на 12-ти предшествующих и 13-ой текущей
метафазах своей метаэволюции, различные формы своей системной организации.
Метафазы метаэволюции живого составляют триады, с нарастанием их сложности и эффективности.
Метафазы метаэволюции живого составляют триады, с нарастанием их сложности и эффективности.

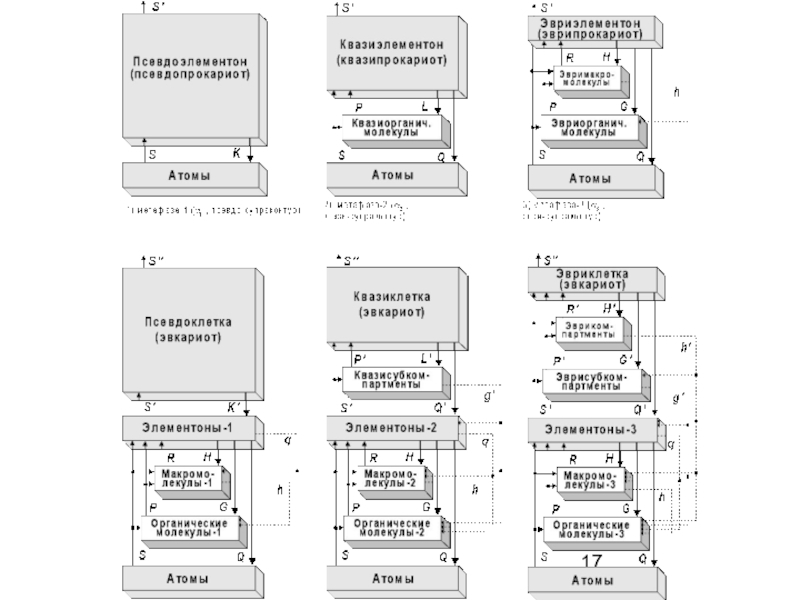
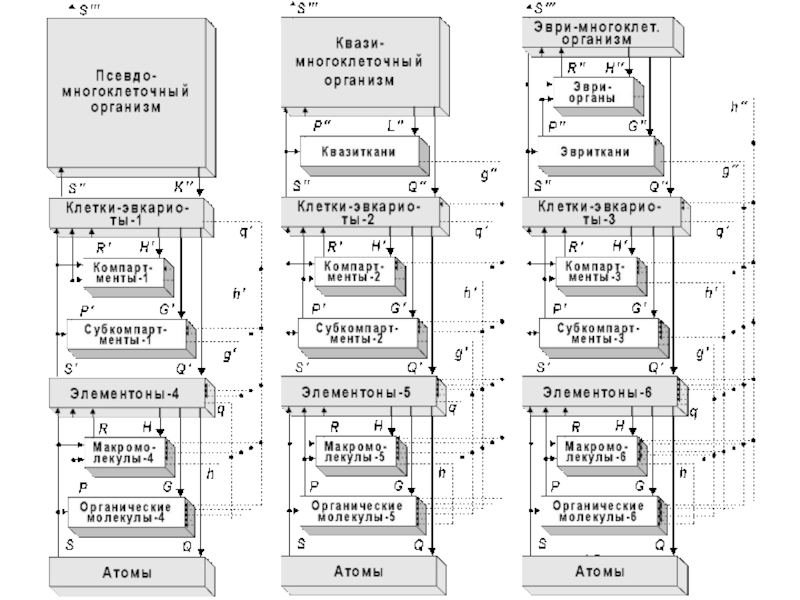
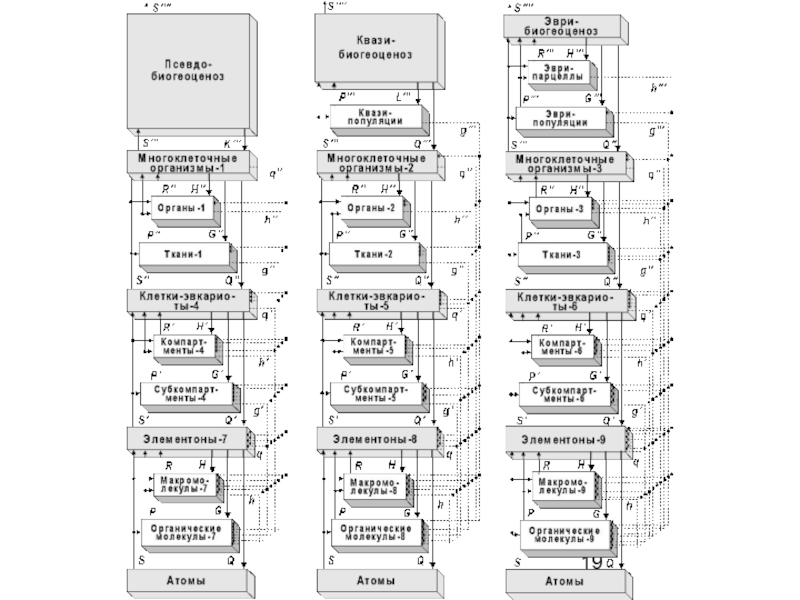
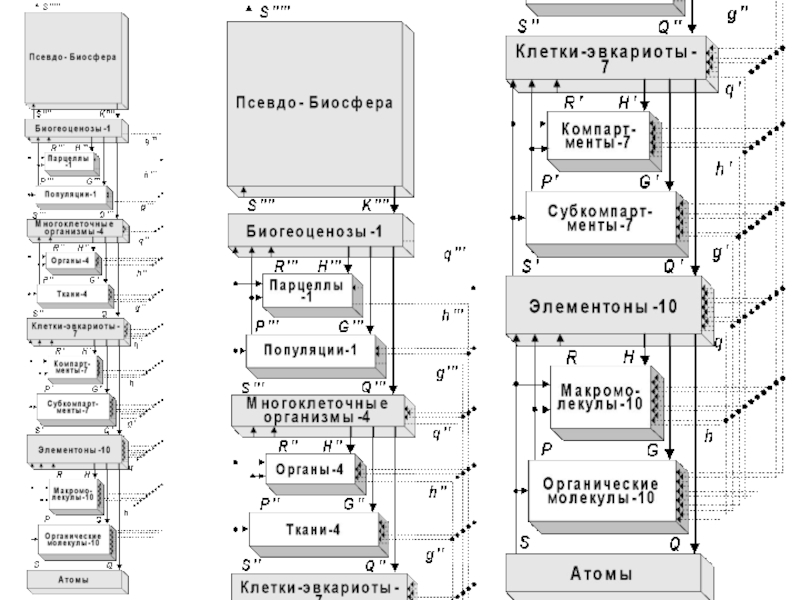
Слайд 21Продолжительность каждой триады (согласно эмпирическим данным) – около 1,01 млрд. лет.
Тенденция усложнения структур, находящихся иерархически «внутри» вновь возникающих, постоянна.
Важно, что все старые иерархические структуры с появлением новых продолжают «параллельно» существовать, коэволюционируя с вновь возникающими.
 – около 1,01 млрд. лет. Тенденция усложнения структур, находящихся")
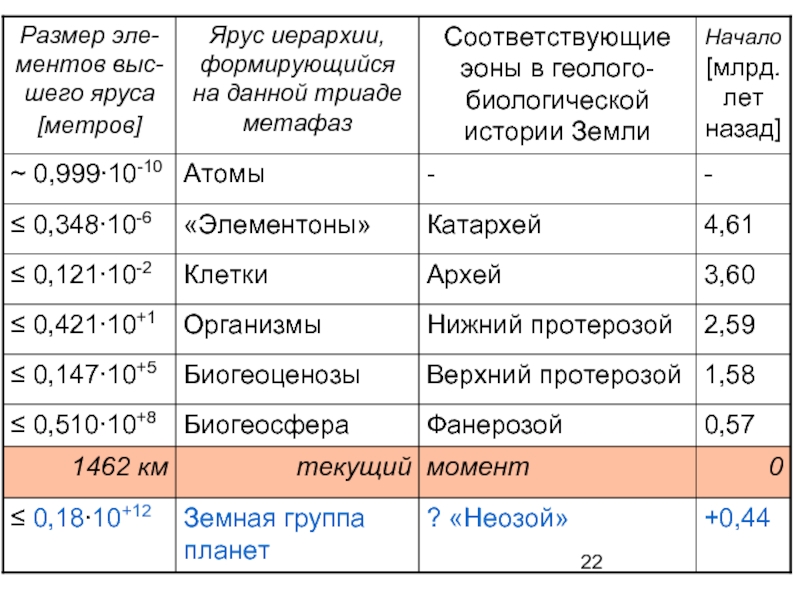
Слайд 23В иерархии системы живого процесс поисковой оптимизации энергетики происходит сразу в
пяти контурах, иерархически вложенных друг в друга и образующих в результате этот целостный процесс.
Поисковую активность проявляют представители всех ярусов в иерархии, а в качестве целезадающих выступают биообъекты, относящиеся к «основным уровням биологической интеграции». Это: 1) «элементоны» (прокариотические ячейки и ультраструктурные внутриклеточные элементы эвкариот), 2) одноклеточные эвкариоты (сложные клетки), 3) многоклеточные организмы, 4) биогеоценозы и 5) Биогеосфера в целом.
Процессы поисковой оптимизации в контурах ярусов 1-3 обычно называют адаптацией (соответственно клеток или организмов). Процессы в контурах ярусов 4-5 целесообразно называть биогеоценотической эволюцией и Биогеосферной эволюцией. Все процессы в контурах ярусов 1-5 происходят одновременно и параллельно, но каждый в своих типичных пространствах и типичных темпах времени!
Поисковую активность проявляют представители всех ярусов в иерархии, а в качестве целезадающих выступают биообъекты, относящиеся к «основным уровням биологической интеграции». Это: 1) «элементоны» (прокариотические ячейки и ультраструктурные внутриклеточные элементы эвкариот), 2) одноклеточные эвкариоты (сложные клетки), 3) многоклеточные организмы, 4) биогеоценозы и 5) Биогеосфера в целом.
Процессы поисковой оптимизации в контурах ярусов 1-3 обычно называют адаптацией (соответственно клеток или организмов). Процессы в контурах ярусов 4-5 целесообразно называть биогеоценотической эволюцией и Биогеосферной эволюцией. Все процессы в контурах ярусов 1-5 происходят одновременно и параллельно, но каждый в своих типичных пространствах и типичных темпах времени!

Слайд 24Микроэволюция, протекающая внутри конкретного вида, является частью биогеоценотической эволюции (с характерным
ареалом – радиусом условного круга – порядка 15 км), в рамках которой этот вид существует и развивается.
Макроэволюция, протекающая на надвидовом уровне, является частью Биогеосферной эволюции, в которой выделяются характерные размеры (порядка 222 км, 3370 км и 51 тыс. км), к которым тяготеют ареалы существования биомов, природных зон и живого на Земле как целого.
Макроэволюция, протекающая на надвидовом уровне, является частью Биогеосферной эволюции, в которой выделяются характерные размеры (порядка 222 км, 3370 км и 51 тыс. км), к которым тяготеют ареалы существования биомов, природных зон и живого на Земле как целого.

Слайд 25Основными средствами реализации поисково-оптимизационного процесса адаптации/эволюции живого являются:
1. селекция направленная (задающая
тенденцию изменения приспособительного поведения субъектов S в направлении экстремума общегруппового энергетического целевого критерия Q),
2. селекция канализирующая (задающая предпочтительные варианты приспособительного поведения субъектов P в составе группы согласно общегрупповому целевому критерию G),
3. селекция ограничивающая (запрещающая нежелательные варианты приспособительного поведения субъектов R в составе группы согласно общегрупповому целевому критерию H),
4. селекция посредством внешнего дополнения W (никак не связанная с проявлениями активности субъектов S, P, R в составе группы – например, катастрофические влияния),
5. канализируемость структурная (посредством параметрических влияний системной памяти М на все перечисленные 4 типа селекции).
2. селекция канализирующая (задающая предпочтительные варианты приспособительного поведения субъектов P в составе группы согласно общегрупповому целевому критерию G),
3. селекция ограничивающая (запрещающая нежелательные варианты приспособительного поведения субъектов R в составе группы согласно общегрупповому целевому критерию H),
4. селекция посредством внешнего дополнения W (никак не связанная с проявлениями активности субъектов S, P, R в составе группы – например, катастрофические влияния),
5. канализируемость структурная (посредством параметрических влияний системной памяти М на все перечисленные 4 типа селекции).

Слайд 26Согласно энергическому критерию поисково-оптимизационного развития системы живого, выживают биообъекты не «наиболее
приспособленные к каким-то конкретным свойствам внешней среды», а наиболее приспособленные энергетически, т.е. интегрально к внешней и внутренней среде биообъекта.

Слайд 27Реконструкция механизма биологической эволюции по Ч.Дарвину. Места для третьей важнейшей составляющей поискового
оптимизационного механизма – системной памяти – в рамках описываемой дарвинистской схемы нет.

Слайд 28
1 – имманентная активность, т.е. «изменчивость», проявляемая генами;
2 –
процесс онтогенеза, отражающий «наследственность»;
3 – проявление специфического поведения организмов в рамках соответствующих популяций;
4 – селектор получаемого на базе конкурентной борьбы между особями результата, определяемый целевой функцией (в данном случае только ограничениями типа равенств и типа неравенств), т.е. «естественный отбор» на базе критерия «приспособленности»;
5 – процесс сообщения отрицательного результата селекции на организменный уровень;
6 – процедура «изъятия» организма из популяции (его гибель или, как минимум, лишение возможности дать новое потомство);
7 – «внешнее дополнение», т.е. внешнее неспецифическое воздействие на организм (например, катастрофического характера: пожар, наводнение, извержение вулкана, резкое похолодание и т.п.), также приводящее к его устранению из популяции.
3 – проявление специфического поведения организмов в рамках соответствующих популяций;
4 – селектор получаемого на базе конкурентной борьбы между особями результата, определяемый целевой функцией (в данном случае только ограничениями типа равенств и типа неравенств), т.е. «естественный отбор» на базе критерия «приспособленности»;
5 – процесс сообщения отрицательного результата селекции на организменный уровень;
6 – процедура «изъятия» организма из популяции (его гибель или, как минимум, лишение возможности дать новое потомство);
7 – «внешнее дополнение», т.е. внешнее неспецифическое воздействие на организм (например, катастрофического характера: пожар, наводнение, извержение вулкана, резкое похолодание и т.п.), также приводящее к его устранению из популяции.

Слайд 29Предложенная концепция позволяет снять многолетнее недоразумение – рассмотрение дарвинистских и номогенетических
теорий эволюции живого как альтернативных, объединяя их модифицированные версии в рамках единой схемы иерархической поисковой оптимизации живой природы.
Это даёт возможность с единых позиций объяснить эмпирические факты, часть из которых противоречит дарвинизму, а часть - номогенезу.
Это даёт возможность с единых позиций объяснить эмпирические факты, часть из которых противоречит дарвинизму, а часть - номогенезу.

Слайд 30«Необходимо отметить, что в биологии термин "эволюция" применяется в различных смыслах.
Связь возникающих при этом проблем с языком описания процесса осознаётся редко, и большей частью они считаются содержательными; в результате возникают длительные, ожесточенные, но совершенно бесплодные дискуссии о том, какая модель эволюции лучше соответствует действительности (...) Корректная же теория, описывающая поведение морфологических структур во времени, по-видимому, должна быть существенно номогенетической (изменения частей подчиняются законам целого), и дискуссия
между сторонниками номогенеза и дарвинистического подхода основана на непонимании того, что эти концепции дополнительны, а не взаимоисключающи» [д.б.н. Жерихин Владимир Васильевич, Раутиан Александр Сергеевич. Филоценогенез и эволюционные кризисы, 1997]
.

Слайд 31«Естественный отбор бесспорно остаётся движущим фактором эволюции, но не единственным. Дарвиновская
и “недарвиновская” эволюции взаимно не противоречивы» [член-корр. РАН Н.Н.Воронцов. Развитие эволюционных идей в биологии. М.: МГУ, Прогресс-Традиция, 1999, С.606].
«Селектогенез не только не отрицает закономерность и направленность эволюции, но и вскрывает биологические причины этих явлений.» [д.б.н., проф. А.С.Северцов. Теория эволюции. М.: ВЛАДОС, 2005, С.351].
«Селектогенез не только не отрицает закономерность и направленность эволюции, но и вскрывает биологические причины этих явлений.» [д.б.н., проф. А.С.Северцов. Теория эволюции. М.: ВЛАДОС, 2005, С.351].

Слайд 32Резюме
1. Моделируется иерархическая система живого, поведение части переменных которой интерпретируется как
процесс эволюции.
2. Основными переменными иерархического механизма системы живой природы являются: а) поисковая приспо-собительная активность его элементов; б) целевой критерий энергетического характера; в) системная память – память структур о прошлом приспособительном поведении.
3. Разделяются понятия «метаэволюция» (процесс последовательного наращивания числа уровней/ярусов в иерархии) и «эволюция» (процессы приспособительного поведения на «верхних» ярусах в системе живого).
4. Дарвинистские и номогенетические теории эволюции живого – их модифицированные версии – объединяются, как непротиворечивые, в рамках единой схемы иерархической поисковой оптимизации живой природы.
2. Основными переменными иерархического механизма системы живой природы являются: а) поисковая приспо-собительная активность его элементов; б) целевой критерий энергетического характера; в) системная память – память структур о прошлом приспособительном поведении.
3. Разделяются понятия «метаэволюция» (процесс последовательного наращивания числа уровней/ярусов в иерархии) и «эволюция» (процессы приспособительного поведения на «верхних» ярусах в системе живого).
4. Дарвинистские и номогенетические теории эволюции живого – их модифицированные версии – объединяются, как непротиворечивые, в рамках единой схемы иерархической поисковой оптимизации живой природы.

Слайд 33Гринченко С.Н. Системная память живого (как основа его метаэволюции и периодической
структуры). М.: ИПИРАН, “Мир”, 2004, 512 с.
Литература
. М.: ИПИРАН, “Мир”,")





