- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Управление памятью (лекция 5) презентация
Содержание
- 1. Управление памятью (лекция 5)
- 2. Перенос памяти - опыты Мак-Коннелла на планариях
- 3. Перенос памяти Схема, иллюстрирующая передачу планариям-реципиентам условных
- 4. Павловское «проторение пути» («Электрическая» теория памяти)
- 5. Механизм Хебба В данной книге постулат Хебба
- 6. Синапсы Хебба Согласно Хеббу, следы памяти могут
- 7. Анатомия гиппокампа Сети гиппокампа: Гиппокамп образует
- 8. Циркуляция в гиппокампе Весь гиппокамп можно рассматривать
- 9. Гиппокампальное представительство физического пространства. Клетки места
- 10. Последовательное поведение клеток места воспроизводится как во
- 11. Роль тэта-ритма Тэта-ритм - это очень точный
- 12. Прецессия фазы тэта-ритма Это выделение разных этапов
- 13. Рабочий цикл с переменой направления передачи Во
- 14. Запоминание по механизму Хебба У грызунов гиппокампальные
- 15. Вычислительная роль движущихся волн Из [Ermentrout GB,
- 16. Разности фаз тэта-волн вдоль оси гиппокампа a-d.
- 17. Топография интраламинарного фазового профиля a-c. Серые кружки
- 18. Протез гиппокампа у крыс. Модель MIMO
- 19. Рекуррентный интерфейс (R-BCI) A -
- 20. Опыты со скотофобином, бродячие и домашние
- 21. Голографическая гипотеза памяти
- 22. Оптическая голограмма Голография создана Деннисом Габором в 1947 г (Нобелевская премия по физике 1971 г)
- 23. Физический принцип голографии Рассеянные объектом волны характеризуются
- 24. Дивергенция и конвергенция в нервной сети
- 25. Нейрональная голограмма
- 26. Спасибо за внимание

Слайд 2Перенос памяти - опыты Мак-Коннелла на планариях
(«химическая» теория памяти)
Американский зоопсихолог Джеймс
Американский зоопсихолог Джеймс Мак-Коннел лизучал поведение")
Слайд 3Перенос памяти
Схема, иллюстрирующая передачу планариям-реципиентам условных рефлексов,
выработанных на свет или
Условные рефлексы вырабатывали у планарий (разделив их на две группы) «хитрым» способом. Каждый из подпороговых раздражителей - вибрацию или освещение - сочетали с электрическим воздействием. Ответ в группах на оба условных раздражителя был одинаковым по форме, но «световые» планарии реагировали только на свет, а «вибрационные» - только на вибрацию. Так образовались две группы обученных планарий. Они и стали жертвами каннибализма - ими покормили планарий-реципиентов. Каждую такую группу в свою очередь поделили пополам и подвергли испытанию на один из условных раздражителей. Итак, половину группы планарий, поевших «световых» доноров, тестировали на свет, а другую половину - на вибрацию. Точно так же поступили с реципиентами «вибрационных» доноров. Вопреки ожиданиям, эффект у реципиентов оказался специфическим, т.е. они отвечали большим количеством реакций только на раздражитель, сигнальный для поеденных ими доноров

")
Слайд 5Механизм Хебба
В данной книге постулат Хебба звучит следующим образом: «When an
Разберем данное утверждение и выделим основные следствия, которые могут быть выведены из представленной формулировки:
Причинно-следственная связь. Основной смысл постулата Хебба заключается в том, что если изначально наблюдается причинно-следственная связь между активациями пре- и постсинаптического нейрона, то эта связь имеет тенденцию к усилению (про обратный закон Хебб в данной формулировке ничего не говорит).
Местоположение изменений. Хебб указывает, что это усиление связанности происходит либо за счет изменения в проводимости синапса (процесс роста), либо за счет изменения метаболических особенностей самих клеток.
Канадский нейропсихолог Дональд Хебб шел к окончательной формулировке своего «нейрофизиологического постулата» достаточно долго, публикуя различные его версии в ранних статьях. Однако, окончательный вид он приобрел в 1949 в самом значимом труде Хебба – «The Organization of Behavior: A NEUROPSYCHOLOGICAL THEORY».

Слайд 6Синапсы Хебба
Согласно Хеббу, следы памяти могут формироваться путем модификации синапсов. На
Сначала (I), когда активен нижний (темный) синапс, происходит возбуждение третьего нейрона, которое не может быть вызвано через слишком слабый верхний (светлый) синапс.
Однако при одновременном воздействии обоих синапсов (II) третий нейрон тоже возбуждается и в нем происходит ряд последовательных биохимических процессов, которые усиливают прежде слабый верхний синапс.
В результате (III) верхний синапс приобретает способность сам по себе вызывать реакцию третьего нейрона.
Совокупное возбуждение.
Хебб не случайно два раза (в начале и в конце формулировки) обращает наше внимание на то, что рассматриваемый пресинаптический нейрон является лишь одним из нейронов, которые принимают участие в возбуждении постсинаптического нейрона. Это утверждение, которое вполне понятно нейрофизиологам, для математиков является достаточно трудным. Такой формулировкой он указывает, что возбуждение постсинаптического нейрона не может быть осуществлено только за счет одного пресинаптического (спайк – это деполяризация мембраны нейрона, а разряд одного пресинатического нейрона никогда не может привести к деполяризации постсинаптического нейрона). В моделях искусственных нейронных сетей этот факт практически всегда нарушается, и к чему приводит такое несоответствие мы рассмотрим ниже

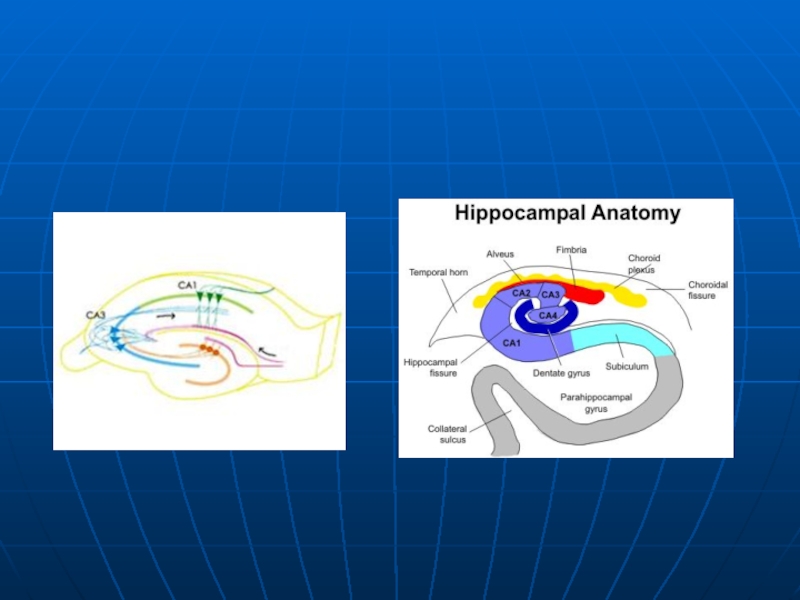
Слайд 7Анатомия гиппокампа
Сети гиппокампа:
Гиппокамп образует главным образом однонаправленные сети, с входом
СА3 нейроны также получают поддержку со стороны DG по мшистым волокнам (MF). Они посылают аксоны к пирамидам поля CA1 с помощью пучка Шаффера (SC), а также к клеткам CA1 в контралатеральном гиппокампе через ассоциативный коммисуральный пучок (AC).
CA1 нейронов также получают прямые входы от перфорантного пути и посылают аксоны к подлежащую ткань (Sb). Эти нейроны в свою очередь, посылают аксон назад (основной выход гиппокампа) в ЕС, образуя петлю.

Слайд 8Циркуляция в гиппокампе
Весь гиппокамп можно рассматривать как одну гигантскую корковую колонку.
Нижний слой зернистых клеток (gc) расходятся ортогонально как входная информация для второго слоя нейронов CA3b/с. Последние проецируются главным образом на CA1, но в значительной степени также возвратно на популяцию CA3a/б. Мшистые клетки на входе (не показаны), которые обеспечивают возбудительную обратную связь для большого числа зернистых клеток, также могут быть представлены как отдельный слой.
Сложная возбуждительная глутамат-ергическая петля в гиппокампе и смежных структурах.
Длинная петля, соединяющая слой 2 энторинальной коры (EC), зернистые клетки (gc), CA3, CA1, и подлежащую ткань (S), возвращается в слой 5 энторинальной коры и дополняется несколькими «срезками» и смежными петлями.
Самый короткий цикл между энторинальной корой и гиппокампом - это путь от слоя 3 энторинальной коры в CA1 и обратно в слой 5 энторинальной коры.
Возбуждающие трафик в несколько петель находится под контролем большой семьи интернейронов, чьи соединения не являются петлеобразными: MC, мшистые клетки на входе; миндалины; ретикулярное ядро таламуса; PFC префронтальная кора, передняя поясная кора.

Слайд 9 Гиппокампальное представительство физического пространства. Клетки места
a. Участки зоны СА1
b. Карта зоны CA1, с точками, показывающими все клетки с полями, картирующими позицию крысы (цвета соответствуют полям). Это клетки места. Трэк пробежки – экспериментальная парадигма. Расширительно «клетки места» – это «клетки элемента обстановки» или «клетки жизненного обстоятельства».
Серые панели - мгновенные снимки тэта-фазы на зоне CA1 в случаях синфазных колебаний (sync) и бегущих волн (wave).
c. Паттерн активности в CA1 если тэта-колебания синфазны. Каждая панель аналогична случаю b, но окрашены только клетки, активные во время каждого временного окна. В каждый момент отдельная тэта-фаза окрашивает всю зону CA1, и, следовательно, клетки, отражающие свой сегмент физического пространства, совместно все активированы.
d. Если тэта-колебания движутся волной, то в каждый момент времени распорядок локальных тэта-фаз картирован по гиппокампу и, следовательно, совместно активированы группы клеток, связанные с сегментами пространства ABCD.

Слайд 10Последовательное поведение клеток места воспроизводится как во время движения в прямом,
Воспроизведение в прямой последовательности (левая вставка, красная рамка) происходит перед обходом окружающей среды и обратное воспроизведение – после (правая вставка, синяя рамка).LFP в зоне CA1 показан сверху и скорость животного показана снизу. Перепечатано из Diba и Ки Buzsaki (2007)
Роль острых волн
(высокоамплитудные LFP, «sharp waves» SPW, или вспышки гамма-ритма)

Слайд 11Роль тэта-ритма
Тэта-ритм - это очень точный способ задания временного хода активности
Гиппокамп является основной структурой, участвующей в формировании памяти, поэтому информация должна достичь гиппокампа во время фазового кодирования. Активное поведение сопровождается гиппокампальными тэта-колебаниями, управляемыми генераторами вне гиппокампа, которые находятся в медиальной перегородке и в энторинальной коре, и взаимодействуют с автономными тэта-осцилляторами в гиппокампе. Большинство клеток гиппокампа вовлечены в тэта-ритм, то есть, они разряжаются преимущественно на определенной фазе тета-волны. Тэта- колебания также возникают в медиальной височной доле и других структурах, тесно связанных с гиппокампом, и регулирующих обмен информацией с гиппокампом
Префронтальная кора (PFC) играет особую роль при обмене, связанном с памятью, между гиппокампом и другими регионами во время активного поведения. Электрофизиологические исследования также показали связь между гиппокампом и PFC при контролируемых поведенческих задачах.
Тем не менее, активность, так или иначе связанная с гиппокампом, распространяется на гораздо большую область неокортекса: в тэта-колебания вовлечены также много синапсов от нейронов гиппокампа, расположенные в сенсорных и ассоциативных областях теменной коры.
Тэта-колебания служат метрономом для передачи информации из неокортекса в гиппокамп и обратно. В неокортексе более быстрые локальные гамма-колебания (30-140 Гц) связываются с предпочтительной фазой тэта-цикла.

Слайд 12Прецессия фазы тэта-ритма
Это выделение разных этапов обработки сигнала вдоль пути «ЭК-гиппокамп»
Это подтверждается привязкой к разным тета-фазам всплесков гамма-LFP, соответствующих афферентам, сходящимся в поле гиппокампа CA1 из поля СА3 и ЭК. ЭК передаёт текущую сенсорную информацию непосредственно в зону CA1, в то время как зона CA3, в динамике которой преобладают обратные связи, может инициировать поиск следа памяти, и затем отправить его в CA1.
Действительно, наличие гамма-всплесков на участке CA3-CA1, и их разделение на определенной тэта-фазе, коррелирует с обучением и эффективностью принятия решений в задаче на рабочую память.
.
Прецессия тета-фазы в CA1 и CA3 (наблюдается также в ЭК, слой 2) постепенно сдвигает разряды клеток места в начало тэта-цикла по мере того, как животное пересекает участок поля. Таким образом, в каждом участке «разрядная» фаза с сдвигается от фазы, сопоставимой с «разрядной» фазой входной структуры, на фазу, в которой входы, наоборот, минимальны, и активность может возникнуть лишь в результате внутренней динамической обработки в гиппокампе предыдущих входных посылок, которые пришли гораздо раньше.
Наклонные стрелки – фазовый сдвиг (отставание) станций друг от друга
Горизонтальные красные стрелочки – прецессия тэта-фазы

Слайд 13Рабочий цикл с переменой направления передачи
Во время кодирования при бодрствовании, а
Медленные колебания создают «рабочий цикл» обмена информацией между структурами с изменениями направления передачи в зависимости от фазы волны.
Это гипотеза «источник-приемник». В рабочем цикле текущий источник и приёмник информации смещаются так, чтобы передача источника пришлась на "восприимчивую" фазу приемника.
Упрощенная схема взаимодействия гиппокампа и коры во время медленных сонных волн.
Клетки неокортекса и ЭК активны в течение UP-фазы, но относительно замолкают во время DOWN-фазы. Неокортекс стимулирует гиппокамп (красные стрелки), хотя нейроны CA3 тоже активны во время DOWN-фазы (автономно без корковых влияний держится след прошлого паттерна), тогда как CA1 работают только в течение UP-фазы.
Активность от CA3 в конце концов прорывается в виде острых волн во время следующей UP-фазы, которые облегчают действие энторинальных входов, т.е. острые волны временно влияют на разряды в CA1, ЭК и неокортексе (синие стрелки). Это и есть сравнение с предыдущим паттерном (синие и красные стрелки упираются друг в друга).
Отклик гиппокампа является максимальным во время острых волн, и последовательность активаций может составлять несколько ритмических разрядов (чёрные стрелки).
Во время DOWN-фаз автономная обработка может посеять семя активационной структуры и породить (наверху -?) острые волны и отклики во время UP-фазы. Некоторые структуры гиппокампа не включены для простоты.

Слайд 14Запоминание по механизму Хебба
У грызунов гиппокампальные «клетки места» активны, когда животное
Таким образом, в пределах каждого тэта-цикла клетки места разряжаются в последовательности, которая повторяет ближайшую предыдущую траекторию (путь животного), а по времени достаточно быстро друг за другом, чтобы вызвать зависящую от времени спайка синаптическую пластичность и, следовательно, помогают хранить эту информацию (запомнить траекторию по механизму Хебба).
Прецессия тэта-фазы повсеместна в гиппокампе, а также наблюдается в ЭК. В сочетании с фазовыми сдвигами между различными структурами, описанными выше, прецессия фазы создает сложные паттерны совместной импульсации через структуры, значение которых должно быть ещё исследовано.
Группы нейронов в зоне CA1, повторно и синхронно активируемых в очень узкое окно времени (около 30 мс, т.е. примерно гамма-цикл) у впадины тета-волны, возможно, соответствуют Хеббиановским ансамблям.

Слайд 15Вычислительная роль движущихся волн
Из [Ermentrout GB, Kleinfeld D, 2001]: Волны обычно
Есть две возможные вычислительные роли волн, основанные на их появлении исключительно как последовательности колебаний в сети с преимущественно короткими синаптическими связями.
Во-первых, периодическая деинактивация повышает чувствительность нейронов к изменениям на входе (афферентации) в другие моменты. Движение волны обеспечивает, что только часть сенсорного поля выключается во время каждого периода. При этом некоторая фракция нейронов максимально чувствительна к изменениям на входе в любой момент времени.
Во-вторых, движущиеся волны могут служить для пометки мгновенной черты в потоке стимулов с уникальной фазой. Так сенсорная активность в определённой пространственной локализации связывается с уникальным значением фазы.
Развилка «волны или синфазность». Перекидка с движущихся волн к синхронности, или почти синхронности может быть важным вычислительным средством. Когерентная активность среди большого числа нейронов могла бы быть средством усиления или ослабления синаптических связей по механизму Хебба. По последним данным (2001), что такая пластичность возникает только, если пре- и постсинаптические спайки появляются в пределах узкого временного окна (приблизительно менее 20 мс) и синаптические изменения критически зависят от взаимного графика пре- и постсинаптической активации. Таким образом, перекидка с движущихся волн к почти синфазности, которая возникает лишь в присутствии стимуляции, может затем служить для запуска синаптической пластичности.

Слайд 16Разности фаз тэта-волн вдоль оси гиппокампа
a-d. Корональный MRI-срез по брегме -
e. Внизу - трёхмерная модель правого гиппокампа со слоем CA1 пирамидных клеток (красный) и тетродами (чёрные). Красная стрелка показывает рострально, чёрная стрелка – латерально. Вверху - вид сверху тетродов, показанных цветными кружками.
f. LPFs, записанные от области stratum oriens слоя CA1 с использованием блока, показанного на e, во время тэта-колебаний. Сигналы от каждого ряда блока перечислены от медиального к латеральному и сложены от рострального к каудальному по цветным кружкам.
g. LFPs, отфильтрованные в полосе 4-10 Гц из интервала, помеченного горизонтальной чертой
, -4.3 (b),")
Слайд 17Топография интраламинарного фазового профиля
a-c. Серые кружки отмечают точки блока, где записывались
d. Контуры изображают общий вид сверху на мозг того, что на a-c. Серые стрелки показывают направление и скорость распространения волны.

Слайд 18Протез гиппокампа у крыс. Модель MIMO – «Много входов / много
Два рычага (показаны зелёным и красным), нажимая которые животное получало вознаграждение, позволили записать режим гиппокампа в процессе обучения.
Животное запоминает, какой именно рычаг приносит ожидаемый результат, благодаря взаимодействию отделов гиппокампа CA1 и CA3.
В процессе обучения мозг трансформирует кратковременные воспоминания в долговременную память. Однако стоит медикаментозными способами отключить гиппокамп, как эта связь (между видами памяти) пропадает.
MIMO-паттерны, определяемые в реальном времени, были задействованы в соответствии с поведенческой парадигмой замкнутой петли. Результаты показали, что модель была в состоянии предсказать успешное выполнение текущей попытки. Таким образом, паттерны, полученные из MIMO-модели, доставляемые как электрическая стимуляция к тем же электродам, при нормальных условиях тестирования улучшают выполнение и, что важнее, делают возможным восстановление, когда доставляются животному с ансамблевой гиппокампальной активностью, «скомпрометированной» фармакологической блокадой синаптической передачи.

Слайд 19Рекуррентный интерфейс (R-BCI)
A - схема передачи сигнала в рекуррентном интерфейсе
B. Плата нейрочипа с контуром, электродный коннектор и батарейка в круглой капсуле, имплантируемой в голову обезьяны [Jackson et al., 2007].
D. Предлагаемая полностью имплантируемая микросистема (University of Michigan).
C. Действия с рекуррентным BCI, ведущие к стойким изменениям в связях [Jackson et al. 2006a].
Вверху. Внутричерепная стимуляция 3-х сайтов в моторной коре с вызываемыми у обезьяны в состоянии покоя тремя разными мышечными ответами (в центре) и разными изометрическими вращениями запястья (справа). Стрелки справа показывают среднее из 200 траекторий вращения.
В середине. Обучение, включающее 2-дневную триггерную микростимуляцию сайта Nstim на каждый спайк, записанный сайтом Nrec во время свободного поведения или сна.
Внизу. После обучения выходной эффект, вызванный сайтом Nrec изменился в сторону ответа, ранее присущего только сайту Nstim. Этот эффект длился неделю.
 A - схема передачи сигнала в рекуррентном интерфейсе (R-BCI) по Mavoori et")
Слайд 20
Опыты со скотофобином, бродячие и домашние собаки;
Опыты, о которых говорит К.Анохин
.")

Слайд 22Оптическая голограмма
Голография создана Деннисом Габором в 1947 г (Нобелевская премия по
")
Слайд 23Физический принцип голографии
Рассеянные объектом волны характеризуются амплитудой и фазой. Регистрация амплитуды
Интерференция возникает, когда в некоторой области пространства складываются несколько волн, частоты которых близки, или совпадают.
При записи голограммы складываются две волны: опорная волна идёт от специального источника, а другая отражается от объекта записи (объектная волна). В этой же области размещают фотопластинкуПри записи голограммы складываются две волны: опорная волна идёт от специального источника, а другая отражается от объекта записи (объектная волна). В этой же области размещают фотопластинку (или иной регистрирующий материал), и на ней возникает сложная картина полос потемнения, которые соответствуют распределению энергии (картине интерференции) в этой области.
Если теперь эту пластинку осветить волной, близкой к опорной, то она преобразует эту волну в волну, близкую к объектной. Таким образом, мы будем видеть (с той или иной степенью точности) такой же свет, какой отражался бы от объекта записи.