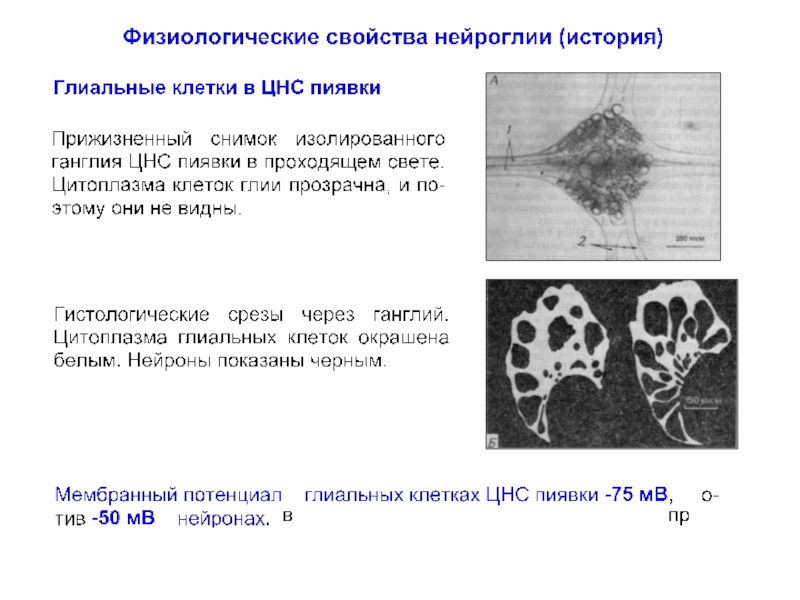
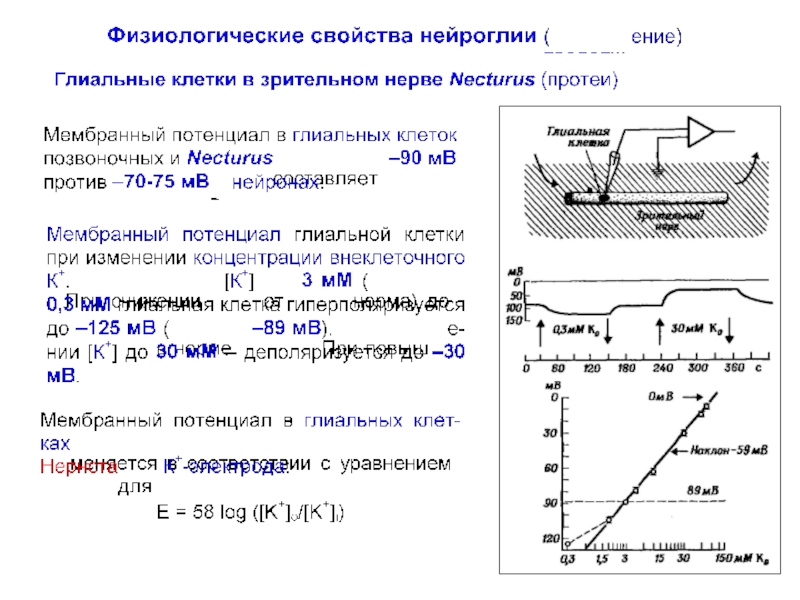
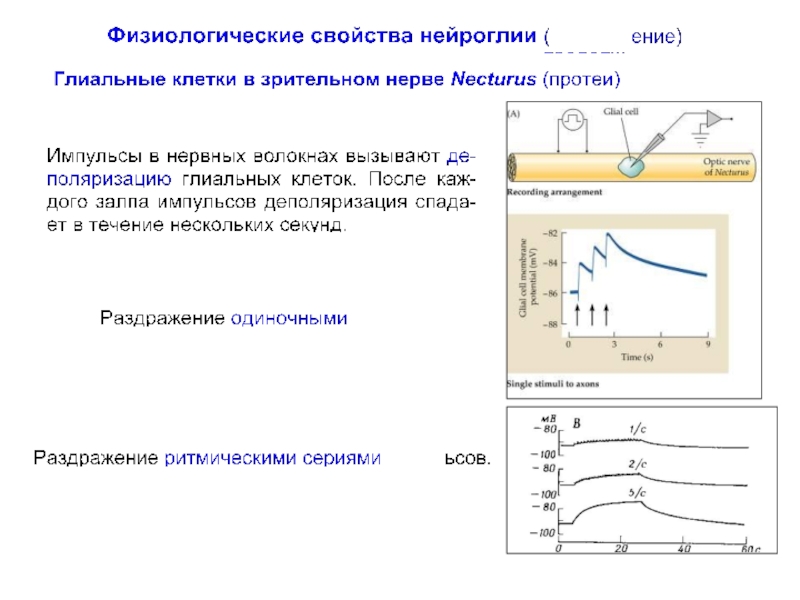
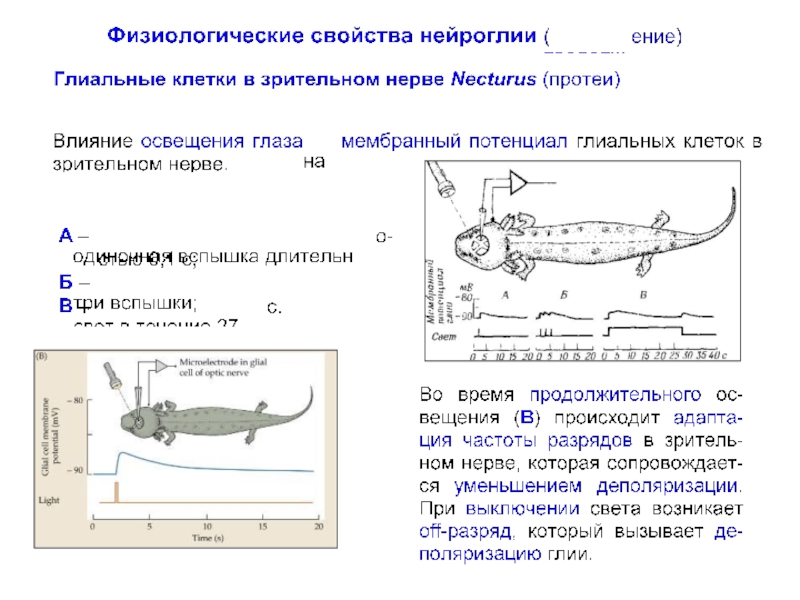
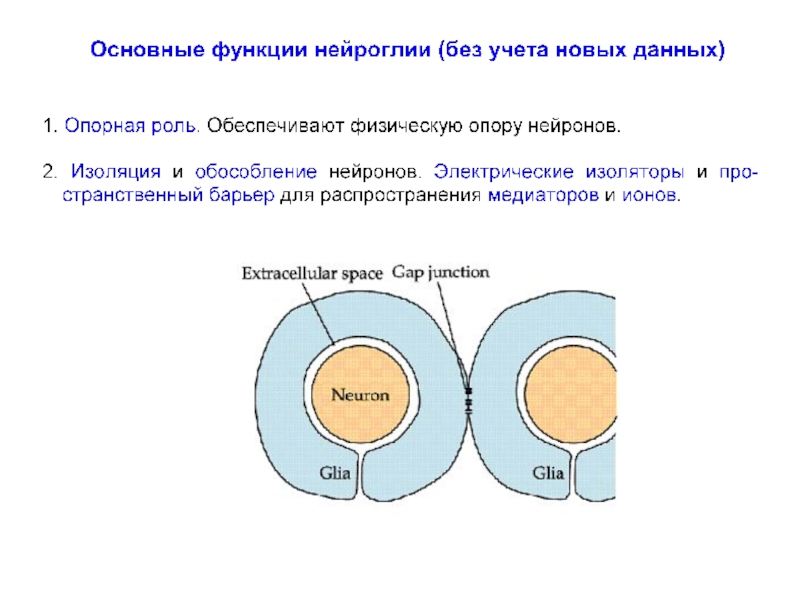
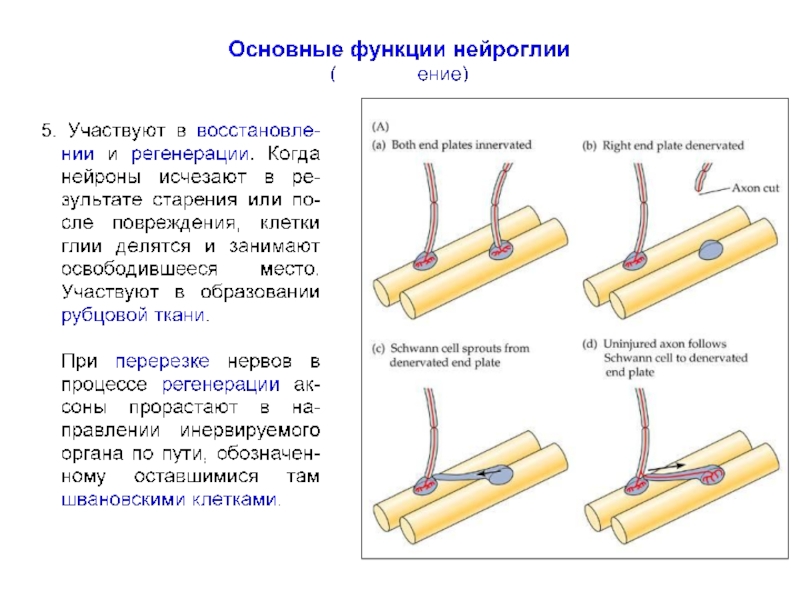
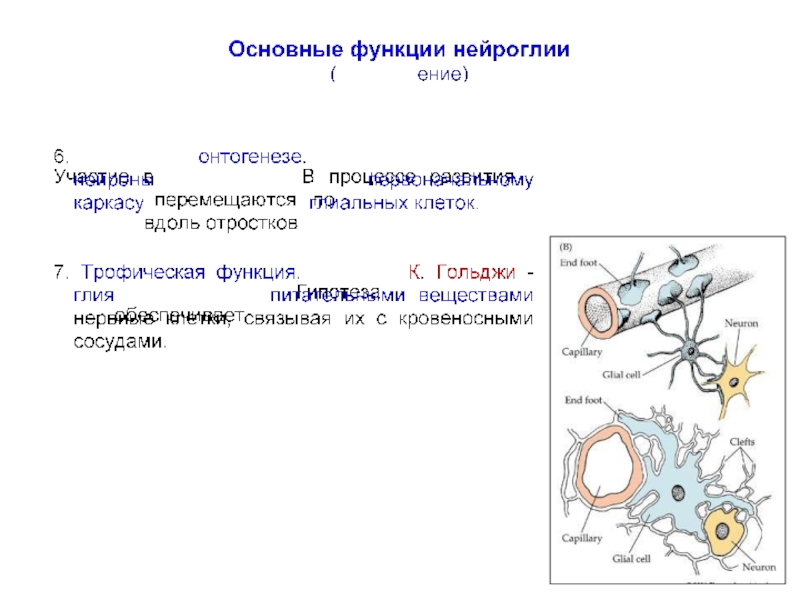
превышает число нейронов примерно в 10 раз, что составляет около половины объема мозга.
Глиальные клетки впервые описаны и так названы Р. Вирховым в 1859 г., который считал их своеобразной разновидностью соединительной ткани («нервным клеем»), расположенной между нервными клетками и «склеивающей» их между собой.
К. Гольджи отмечал, что глия отличается от обычной соединительной ткани как морфологически, так и химически и, кроме того, имеет иное эмбриональное происхождение.
В отличие от нервных клеток глиальные клетки способны делиться во взрослом организме и часто являются источником опухолей мозга и так называемых «глиальных рубцов» при регенерации пораженной нервной ткани.
По строению и свойствам глиальные клетки беспозвоночных (пиявка) и позвоночных (земноводные, млекопитающие) животных принципиально сходны.
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Нейроглия. Глиальные клетки презентация
Содержание
- 1. Нейроглия. Глиальные клетки
- 2. Типы глиальных клеток Глию подразделяют на микро-
- 3. Типы глиальных клеток: астроциты Астроциты занимают положение
- 4. Типы глиальных клеток: олигодендроциты Образуют миелиновую оболочку
- 5. Типы глиальных клеток: олигодендроциты Образуют миелиновую оболочку
- 6. Типы глиальных клеток: клетки радиальной глии Имеют
- 7. Типы глиальных клеток: клетки радиальной глии Radial
- 8. Типы глиальных клеток: клетки радиальной глии Имеют
- 9. Типы глиальных клеток: клетки радиальной глии В
- 10. Типы глиальных клеток: клетки радиальной глии В сетчатке глаза аналогами радиальной глии являются Мюллеровские клетки.
- 11. Типы глиальных клеток: микроглия Клетки микроглии представляют
- 12. Взаимодействие астроцитов и нейронов Classic concepts and
- 13. Взаимодействие астроцитов и нейронов Diffusion of neurotransmitter
- 14. Тройственный синапс = астроцит + пре- +
- 15. Взаимодействие астроцит-синапс-капилляр Interactions among astrocytes, synapses and
- 16. Взамиодействие астроцит-астроцит Interactions among astrocytes, synapses and
- 17. Регуляторная роль взаимодействия астроцит-нейрон Regulatory neuron–astrocyte interaction
- 18. Регуляторная роль взаимодействия астроцит-нейрон Regulatory neuron–astrocyte interaction
- 19. Регуляторная роль взаимодействия астроцит-нейрон Regulatory neuron–astrocyte interaction
- 20. Глиальные клетки генерируют спайки (?!) В мембранах
- 21. Глиальные клетки генерируют спайки (?!) NG2+-клетки подразделяются
- 22. Глиальные клетки генерируют спайки (?!) Методом пэтч-кламп
- 23. Глиальные клетки генерируют спайки (?!) NG2+-клетки с
- 24. Глиальные клетки генерируют спайки (?!) Káradóttir et
- 25. Глиальные клетки генерируют спайки (?!) В ответ
- 26. Активный транспорт и рецепторы в глиальных клетках
- 27. Участие глии в регуляции проведения возбуждения по
- 28. Участие глии в регуляции проведения возбуждения по
- 29. Деполяризация олигодендроцита улучшает проводимость в аксоне Schematic
- 30. Depolarization of oligodendrocytes in the rat hippocampus
Слайд 1Нейроглия
Нейроны мозга тесно окружены глиальными клетками, или глией. Количество глиальных клеток

Слайд 2Типы глиальных клеток
Глию подразделяют на микро- и макроглию, иногда к глиальным
клеткам относят и клетки эпендимы.
Макроглия включает:
- астроциты,
- олигодендроциты,
- Шванновские клетки (аналоги олигодендроцитов в периферических аксонах)
- и клетки радиальной глии.
Клетки микроглии представляют собой популяцию клеток-фагоцитов нервной системы. По строению и свойствам клетки они напоминают макрофаги крови, от которых они предположительно и происходят.
Клетки эпендимы выстилают поверхность желудочков мозга и считаются глиальными, поскольку являются источником глиальных клеток в процессах эмбриогенеза и прижизненного нейрогенеза.
Макроглия включает:
- астроциты,
- олигодендроциты,
- Шванновские клетки (аналоги олигодендроцитов в периферических аксонах)
- и клетки радиальной глии.
Клетки микроглии представляют собой популяцию клеток-фагоцитов нервной системы. По строению и свойствам клетки они напоминают макрофаги крови, от которых они предположительно и происходят.
Клетки эпендимы выстилают поверхность желудочков мозга и считаются глиальными, поскольку являются источником глиальных клеток в процессах эмбриогенеза и прижизненного нейрогенеза.

Слайд 3Типы глиальных клеток: астроциты
Астроциты занимают положение между нейронами и кровеносными капиллярами
и подразделяются на две группы.
Фиброзные астроциты содержат в цитоплазме много филаментов и локализованы преимущественно среди миелинизированных волокон.
Протоплазматические астроциты содержат меньше фиброзного материала и окружают тела нейронов, их дендриты и синаптические контакты.

Слайд 4Типы глиальных клеток: олигодендроциты
Образуют миелиновую оболочку крупных аксонов нейронов ЦНС.
В нервах
и ганглиях вегетативной нервной системы аналогами олигодендроцитов являются Шванновские клетки, которые формируют миелиновую оболочку (My) вокруг крупных аксонов (Ax), характеризующихся высокой скорость проведения нервных импульсов.
75 nm

Слайд 5Типы глиальных клеток: олигодендроциты
Образуют миелиновую оболочку крупных аксонов нейронов ЦНС.
Олигодендроциты и
Шванновские клетки отличаются разным происхождением в эмбриогенезе: первые образуются из клеток-предшественниц, выстилающих мозговую трубку (эпендимы), а вторые - из нервного гребня.

Слайд 6Типы глиальных клеток: клетки радиальной глии
Имеют длинные отростки, которые образуют своеобразные
пути (тракты), вдоль которых развивающиеся в процессе нейрогенеза нервные клетки мигрируют к местам своего назначения.
А - на срезе развивающейся затылочной коры плода обезьяны радиальные волокна расположены вдоль путей миграции формирующихся нейронов от вентрикулярной зоны (внизу) к поверхностным слоям (вверху).
Б - клетки коры мигрируют к местам своего назначения с помощью специальных (ведущих) отростков, ориентированных вдоль волокон радиальной глии как своеобразных «направляющих» (проводников).
Клетки 1, 2, 3 – развивающейся нейроны на разных этапах миграции из вентрикулярной зоны в поверхностные слои. Несколько поперечных срезов через «мигрирующие» клетки (а-г) демонстрируют, что они «охватывают» ствол волокна радиальной глии (выглядит серым) всей своей поверхностью на протяжении пути «миграции».
, вдоль которых")
Слайд 7Типы глиальных клеток: клетки радиальной глии
Radial glia perform support and guidance
functions for migrating neurons. In early development, radial glia span the thickness of the expanding brain parenchyma. (Inset) Defined layers of the neural tube from the ventricular to the outer surface: VZ, ventricular zone; IZ, intermediate zone; CP, cortical plate; MZ, marginal zone. The radial process of the glial cell is indicated in blue, and a single attached migrating neuron is depicted at the right.

Слайд 8Типы глиальных клеток: клетки радиальной глии
Имеют длинные отростки, которые образуют своеобразные
пути (тракты), вдоль которых развивающиеся в процессе нейрогенеза нервные клетки мигрируют к местам своего назначения.
Фрагменты видеосъемки миграции нейрона гиппокампа вдоль волокна радиальной глиальной клетки in vitro.
, вдоль которых")
Слайд 9Типы глиальных клеток: клетки радиальной глии
В ЦНС аналогами радиальной глии являются
клетки Бергмана мозжечка (выделены красным).
Зеленые– протоплазматические астроциты.
Синие – фиброзные астроциты.
Зеленые– протоплазматические астроциты.
Синие – фиброзные астроциты.

Слайд 10Типы глиальных клеток: клетки радиальной глии
В сетчатке глаза аналогами радиальной глии
являются Мюллеровские клетки.

Слайд 11Типы глиальных клеток: микроглия
Клетки микроглии представляют собой популяцию клеток-фагоцитов нервной системы.
По строению и свойствам они напоминают макрофаги крови, от которых они предположительно и происходят.
Activation of microglial cells in a tissue section from human brain. Resting microglia in normal brain (A). Activated microglia in diseased cerebral cortex (B) have thicker processes and larger cell bodies. In regions of frank pathology (C) microglia transform into phagocytic macrophages, which can also develop from circulating monocytes that enter the brain. Arrow in B indicates rod cell. Sections stained with antibody to ferritin. Scale bar = 40 μm.

Слайд 12Взаимодействие астроцитов и нейронов
Classic concepts and new concepts of neuron–astrocyte interaction.
According
to classic concepts astrocytes are recognized as passive and supporting elements in the brain.
Since the discovery of neurotransmitter receptors on astrocytes and the release of transmitters from them, new concepts of neuron–astrocyte interaction as information processing elements have been established.
Since the discovery of neurotransmitter receptors on astrocytes and the release of transmitters from them, new concepts of neuron–astrocyte interaction as information processing elements have been established.

Слайд 13Взаимодействие астроцитов и нейронов
Diffusion of neurotransmitter in extracellular space (spill over).
Before disposal of the released neurotransmitters by specific transporters, they diffuse into the extracellular space and activate neurotransmitters receptors expressed on the astrocytes.
(2) Expression of many kinds of receptors. Astrocytes have been demonstrated to express receptors for neurotransmitters, such as glutamate, noradrenalin, serotonin, GABA and acetylcholine and also for trophic factors. Activation of these receptors induces the increase in [Ca2+]i.
(2) Expression of many kinds of receptors. Astrocytes have been demonstrated to express receptors for neurotransmitters, such as glutamate, noradrenalin, serotonin, GABA and acetylcholine and also for trophic factors. Activation of these receptors induces the increase in [Ca2+]i.
(3) Characteristic Ca2+ increase. The activation of astrocytes through receptors results in an oscillatory increase in [Ca2+]i.
(4) Neurotransmitter release. Activation of astrocytes sometimes causes release of neurotransmitters, such as glutamate and ATP. Released glutamate will activate neuronal cells.
(5) Inhibitory regulation by ATP released from astrocytes has been demonstrated to depress neuronal activities.
. Before disposal of the")
Слайд 14Тройственный синапс = астроцит + пре- + постсинаптический нейроны
Tripartite synapse
The structure
consisting of (1) pre-, (2) post-synaptic neurons and (3) astrocytes has been emphasized by the coining of the term “tripartite synapse”.
Information processing between pre- and post-synaptic neurons receives further modulation from astrocytes, which express neurotransmitter receptor and release neurotransmitters.
Information processing between pre- and post-synaptic neurons receives further modulation from astrocytes, which express neurotransmitter receptor and release neurotransmitters.
 pre-,")
Слайд 15Взаимодействие астроцит-синапс-капилляр
Interactions among astrocytes, synapses and vasculature
The [Ca2+]i increase in astrocytes
induced by neurotransmitter propagates intra- and inter-cellularly. The propagation may be promoted by two mechanisms:
(1) One is diffusion of IP3 inside the cell and also through the gap junctions formed between astrocytes.
(2) The other is the response mediated by released ATP and its receptor (purinergic receptor).
(1) One is diffusion of IP3 inside the cell and also through the gap junctions formed between astrocytes.
(2) The other is the response mediated by released ATP and its receptor (purinergic receptor).

Слайд 16Взамиодействие астроцит-астроцит
Interactions among astrocytes, synapses and vasculature
ATP released through the gap
junction hemichannel diffuses to adjacent astrocytes and activates their ATP receptors.
Functional molecules for regulating the astrocytes, such as purine receptor and aquaporin-4 are expressed mainly on the endfoot of astrocytes, which makes tight contact with a blood vessel.
Since synaptic activities give and receive information between astrocytes, the size of the blood vessel may also be regulated depending upon the neuronal activities. The structure consisting of astrocyte, blood vessel and neuron will provide dynamic regulation of information processing in the brain. The gap junction hemichannel will participate in the release not only of ATP but also of some other transmitters and of trophic factors.
Functional molecules for regulating the astrocytes, such as purine receptor and aquaporin-4 are expressed mainly on the endfoot of astrocytes, which makes tight contact with a blood vessel.
Since synaptic activities give and receive information between astrocytes, the size of the blood vessel may also be regulated depending upon the neuronal activities. The structure consisting of astrocyte, blood vessel and neuron will provide dynamic regulation of information processing in the brain. The gap junction hemichannel will participate in the release not only of ATP but also of some other transmitters and of trophic factors.

Слайд 17Регуляторная роль взаимодействия астроцит-нейрон
Regulatory neuron–astrocyte interaction
(1) Synaptogenesis
Astrocytes play important roles in
synaptogenesis during development. Apoprotein E (cholesterol complexed with apolipoprotein E-containing lipoproteins) released from astrocytes has been shown to be a factor facilitating synapse formation.
Direct contact of astrocytes and neuronal cells through integrin receptor induces drastic synaptogenesis, which is mediated by protein kinase C (PKC) activation.
Direct contact of astrocytes and neuronal cells through integrin receptor induces drastic synaptogenesis, which is mediated by protein kinase C (PKC) activation.
 SynaptogenesisAstrocytes play important roles in synaptogenesis during development. Apoprotein")
Слайд 18Регуляторная роль взаимодействия астроцит-нейрон
Regulatory neuron–astrocyte interaction
(2) Activation of N-methyl-D-aspartic acid (NMDA)
receptor by D-serine
D-Serine is produced from L-serine only inside astrocytes by a specific enzyme, serine racemase. The amino acid is an effective co-activator (binds with glycine site) for NMDA receptor, a key receptor for synaptic plasticity.
D-Serine is produced from L-serine only inside astrocytes by a specific enzyme, serine racemase. The amino acid is an effective co-activator (binds with glycine site) for NMDA receptor, a key receptor for synaptic plasticity.
 Activation of N-methyl-D-aspartic acid (NMDA) receptor by D-serineD-Serine is")
Слайд 19Регуляторная роль взаимодействия астроцит-нейрон
Regulatory neuron–astrocyte interaction
(3) Regulation of neuron–astrocyte interaction by
Ca2+-permeable AMPA receptors
Ca2+-permeable AMPA-type glutamate receptors (without GluR2 subunit) expressed on the Bergman glia are indispensable for proper structural and functional regulation of the Bergmann glia and glutamatergic synapses.
Ca2+-permeable AMPA-type glutamate receptors (without GluR2 subunit) expressed on the Bergman glia are indispensable for proper structural and functional regulation of the Bergmann glia and glutamatergic synapses.
(4) Ectopic release of glutamate from neuron to astrocyte
The ascending fiber terminal releases glutamate and directly activates Ca2+ permeable AMPA receptor expressed on the Bergman cell.
 Regulation of neuron–astrocyte interaction by Ca2+-permeable AMPA receptors Ca2+-permeable")
Слайд 20Глиальные клетки генерируют спайки (?!)
В мембранах астроцитов и Шванновских клеток найдены
K+- и Cl--каналы, проводящие токи утечки, а также потенциал-зависимые Na+- и Ca2+-каналы. Долгое время считалось, что, несмотря на наличие потенциал-зависимых каналов, активация мембран глиальных клеток не приводит к генерации ПД. Однако публикации 2008 г. изменили традиционные представления о физиологических свойствах глии.
В мозжечке млекопитающих как у развивающихся, так и взрослых животных идентифицированы (методами гистохимии) и исследованы (методом пэтч-кламп) глиальные клетки, содержащие протеогликан NG2+. В разные годы эти клетки считались предшественниками олигодендроцитов, «полидендроцитами» и «синаптоцитами».
В мозжечке млекопитающих как у развивающихся, так и взрослых животных идентифицированы (методами гистохимии) и исследованы (методом пэтч-кламп) глиальные клетки, содержащие протеогликан NG2+. В разные годы эти клетки считались предшественниками олигодендроцитов, «полидендроцитами» и «синаптоцитами».
В мембранах астроцитов и Шванновских клеток найдены K+- и Cl--каналы, проводящие")
Слайд 21Глиальные клетки генерируют спайки (?!)
NG2+-клетки подразделяются на два отдельных класса:
(1) с
типичными глиальными свойствами без синаптических контактов, без лиганд-зависимых каналов и с минимальными возбуждающими мембранными токами (no INa cells),
(2) с такими же, как и у нейронов синаптическими входами, выраженными глутамат-зависимыми синаптическими токами и существенными потенциал-зависимыми Na+-токами (INa cells).
(2) с такими же, как и у нейронов синаптическими входами, выраженными глутамат-зависимыми синаптическими токами и существенными потенциал-зависимыми Na+-токами (INa cells).
NG2+-клетки подразделяются на два отдельных класса:(1) с типичными глиальными свойствами без")
Слайд 22Глиальные клетки генерируют спайки (?!)
Методом пэтч-кламп в мембранах INa cells зарегистрированы
потенциал-зависимые Na+-токи, обуславливающие генерацию истинных ПД (a). На (b) ответы no INa cells.
Так же как и в нейронах, эти токи демонстрируют зависимость от мембранного потенциала и обратимо блокируются ТТХ (с).
Так же как и в нейронах, эти токи демонстрируют зависимость от мембранного потенциала и обратимо блокируются ТТХ (с).
Методом пэтч-кламп в мембранах INa cells зарегистрированы потенциал-зависимые Na+-токи, обуславливающие генерацию")
Слайд 23Глиальные клетки генерируют спайки (?!)
NG2+-клетки с нейронным фенотипом получают обычные возбуждающие
синаптические глутаматные входы от немиелинизированных аксонов в белом веществе, а также возбуждающие и тормозные входы в сером веществе.
NG2+-клетки с нейронным фенотипом получают обычные возбуждающие синаптические глутаматные входы от")
Слайд 24Глиальные клетки генерируют спайки (?!)
Káradóttir et al. (2008) report that some
NG2+ glia can be induced to fire action potentials by excitatory synaptic input (highlighted by red starbursts) formed by myelinated (top) or unmyelinated (bottom) axons.
The physiological role of NG2+ cell excitability is not yet clear, but, as suggested by the authors, may influence the transition of NG2+ cells into myelinating oligodendrocytes. These NG2+ cells have processes that wrap nodes of Ranvier, and they might therefore be well positioned for a role in optimizing nodal spacing or stability (?).
The physiological role of NG2+ cell excitability is not yet clear, but, as suggested by the authors, may influence the transition of NG2+ cells into myelinating oligodendrocytes. These NG2+ cells have processes that wrap nodes of Ranvier, and they might therefore be well positioned for a role in optimizing nodal spacing or stability (?).
Káradóttir et al. (2008) report that some NG2+ glia can be")
Слайд 25Глиальные клетки генерируют спайки (?!)
В ответ на деполяризвцию INa cells генерируют
истинные ПД (a). На (с) отсутствие ПД у no INa cells.
Так же как и в нейронах ПД обратимо блокируются ТТХ (b).
Так же как и в нейронах ПД обратимо блокируются ТТХ (b).
Спонтанные ПД
В ответ на деполяризвцию INa cells генерируют истинные ПД (a). На")
Слайд 26Активный транспорт и рецепторы в глиальных клетках
В мембранах глиальных клеток обнаружены
системы активного транспорта ионов, необходимые для обеспечения возбуждающей функции:
ионные помпы для трансмембранного транспорта Na+, K+ и H+,
а также Na+-зависимый Cl-/HCO3--обменник.
В мембранах астроцитов, олигодендроцитов и Шванновских клеток также представлены
- транспортеры для глутамата и ГАМК
- рецепторы для некоторых медиаторов - glutamate, noradrenalin, serotonin, GABA and acetylcholine and also for trophic factors.
ионные помпы для трансмембранного транспорта Na+, K+ и H+,
а также Na+-зависимый Cl-/HCO3--обменник.
В мембранах астроцитов, олигодендроцитов и Шванновских клеток также представлены
- транспортеры для глутамата и ГАМК
- рецепторы для некоторых медиаторов - glutamate, noradrenalin, serotonin, GABA and acetylcholine and also for trophic factors.

Слайд 27Участие глии в регуляции проведения возбуждения
по аксонам
Нейроноподобные NG2+-клетки, получающие возбуждающие
и тормозные синаптические входы от аксонов и генерирующие настоящие ПД участвуют в регуляции проведения возбуждения по аксонам.
NG2+-клетки направляют свои отростки к перехватам Ранвье миелинизированных аксонов. Такие возбудительные посылки увеличивают эффективность проведения возбуждения через перехваты Ранвье.
NG2+-клетки направляют свои отростки к перехватам Ранвье миелинизированных аксонов. Такие возбудительные посылки увеличивают эффективность проведения возбуждения через перехваты Ранвье.
С одной стороны NG2+-клетки получают синаптические входы от аксонов и тем самым детектируют различные нарушения в прохождении ПД по этим аксонам.
С другой стороны они структурно взаимодействуют с перехватами («окутывают» их).

Слайд 28Участие глии в регуляции проведения возбуждения
по аксонам
Предполагается, что эти клетки
при взаимодействии с близлежащими олигодендроцитами и аксонами могут укорачивать межперехватные участки, вызывать увеличение диаметра аксона или повышать плотность Na+-каналов в мембране перехватов. Действительно в окончаниях NG2+-клеток, контактирующих с перехватами, присутствует миелин-гликопротеин олигодендроцитов, и у трансгенных мышей, у которых отсутствует ген, кодирующий этот белок, отмечается нарушение роста аксональной мембраны в перехватах.
Такие функциональные особенности NG2+-клеток указывают на их возможную регуляторную роль в ремиелинизации аксонов после различных мозговых нарушений (параличей, апоплексии мозга и травм спинного мозга), приведших к демиелинизации проводящих путей.
Такие функциональные особенности NG2+-клеток указывают на их возможную регуляторную роль в ремиелинизации аксонов после различных мозговых нарушений (параличей, апоплексии мозга и травм спинного мозга), приведших к демиелинизации проводящих путей.

Слайд 29Деполяризация олигодендроцита улучшает проводимость в аксоне
Schematic diagram of experiments by Yamazaki
et al. (Neuron Glia Biology. 2007. 3: 325-344) revealing that depolarization of oligodendrocytes can decrease the latency of action potentials in axons in the rat hippocampus. Theta burst stimulation of axons depolarized the oligodendrocytes by activating glutamate and potassium channels in the oligodendrocyte. Paired cell recordings between oligodendrocytes (Roligo) and pyramidal neurons (Rpyr) were used to monitor and regulate voltage responses in oligodendrocytes. Action potentials were stimulated by antidromic stimulation (S) for measurements of action potential conduction velocity. Conduction velocity increased in axons myelinated by the oligodendrocyte that was depolarized.

Слайд 30Depolarization of oligodendrocytes in the rat hippocampus can reduce action potential
latency through the axons ensheathed by the cell.
A, Axon (triangles) and oligodendrocyte filled with biocytin in paired whole-cell recordings.
B, Depolarization of the oligodendrocyte reduced the spike latency in axons ensheathed by the oligodendrocyte within seconds, and the latency returned to baseline after restoring normal resting potential in the oligodendrocyte. Reprinted from Yamazaki and others (2008), with permission
A, Axon (triangles) and oligodendrocyte filled with biocytin in paired whole-cell recordings.
B, Depolarization of the oligodendrocyte reduced the spike latency in axons ensheathed by the oligodendrocyte within seconds, and the latency returned to baseline after restoring normal resting potential in the oligodendrocyte. Reprinted from Yamazaki and others (2008), with permission
Деполяризация олигодендроцита улучшает проводимость в аксоне