Г.А.
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Иммунитет в защите и повреждении организма. (Лекция 6) презентация
Содержание
- 1. Иммунитет в защите и повреждении организма. (Лекция 6)
- 2. Часть1 Противоинфекционный иммунитет
- 3. ПРОТИВОИНФЕКЦИОННЫИ ИММУНИТЕТ – первая область
- 4. ФОРМИРОВАНИЕ ПРОТИВОИНФЕКЦИОННОГО ИММУНИТЕТА : По направленности
- 5. ПУТИ ПОСТУПЛЕНИЯ И РАСПРОСТРАНЕНИЯ ИНФЕКЦИОННЫХ АГЕНТОВ определяют:
- 6. КЛЮЧЕВОЙ ФАКТОР ОПРЕДЕЛЕНИЯ ТИПА ИММУННОГО ОТВЕТА Если
- 7. Локализация и эффективные пути иммунной защиты
- 8. Локализация и эффективные пути иммунной защиты
- 9. ДВЕ ТРИАДЫ ОБЪЕДИНЯЮЩИЕ ВРОЖДЕННЫЙ И ПРИОБРЕТЕННЫЙ ИММУНИТЕТ
- 10. ВРОЖДЕННЫЙ ИММУНИТЕТ ПРИ ВНЕКЛЕТОЧНЫХ ИНФЕКЦИЯХ
- 11. Индукция воспалительного ответа.
- 12. РАННИЙ ОТВЕТ ПРИ ВНЕКЛЕТОЧНЫХ ИНФЕКЦИЯХ
- 13. ФОРМИРОВАНИЕ ПОЛНОЦЕННОГО АДАПТИВНОГО ИММУННОГО ОТВЕТА ПРИ ВНЕКЛЕТОЧНЫХ
- 14. СХЕМА ИММУННОГО ОТВЕТА ПРИ ВНЕКЛЕТОЧНЫХ ИНФЕКЦИЯХ (антибактериальный иммунный ответ)
- 15. Второй этап: Th1-зависимый клеточный ответ. При внеклеточной
- 16. ОСОБЕННОСТИ ПАТОГЕНОВ МИКРООРГАНИЗМОВ. Патогенные свойства м/о во
- 17. ЭНДОТОКСИНЫ ЛИПОПОЛИСАХАРИДНОЙ ПРИРОДЫ ГРАМ(-): активируют комплемент, к
- 18. ПЕПТИДОГЛИКАНЫ БАКТЕРИАЛЬНОЙ СТЕНКИ - МУРАМИЛДИПЕПТИДЫ (ГРАМ +).
- 19. ПУТИ ЗАЩИТЫ МИКРООРГАНИЗМОВ ОТ ИММУННОГО ОТВЕТА 1.Вариабельность.
- 20. СРЕДИ ФАКТОРОВ, ПОЗВОЛЯЮЩИХ М/О ИЗБЕЖАТЬ ЗАЩИТНЫХ МЕХАНИЗМОВ,
- 21. ИММУННЫЙ ОТВЕТ ПРИ ВНУТРИКЛЕТОЧНОЙ ИНФЕКЦИИ Вирусы –
- 22. ВРОЖДЕННЫЙ ИММУНИТЕТ ПРИ ВНУТРИКЛЕТОЧНЫХ ИНФЕКЦИЯХ Первые часы
- 23. Индукция воспалительного ответа. На 2-4 сутки,
- 24. АДАПТИВНЫЙ ИММУННЫЙ ОТВЕТ ПРИ ВНУТРИКЛЕТОЧНЫХ ИНФЕКЦИЯХ Th1-
- 25. Противовирусный иммунитет
- 26. ГУМОРАЛЬНЫЙ АДАПТИВНЫЙ ИММУННЫЙ ОТВЕТ ПРИ ВНУТРИКЛЕТОЧНЫХ ИНФЕКЦИЯХ
- 27. СТРАТЕГИЯ ОБХОДА ВИРУСАМИ ИММУНОЛОГИЧЕСКОГО КОНТРОЛЯ 1.Вариабельность
- 28. 2.Модификация вирусами активности иммунной системы а)Блокада активации
- 29. 3.ОТКЛОНЕНИЯ ОТ «КЛАССИЧЕСКОГО» ВАРИАНТА ИММУННОГО ОТВЕТА ПРИ
- 30. 4.Неадекватная тактика иммунного ответа Th1-зависимые клеточные
- 31. 5.Переход вируса в латентную фазу.
- 32. 6.Непосредственное инфицирование ЛФ и МФ Некоторые вирусы,
- 33. 7.Перекрестная реактивность АГ микро- и макроорганизма При
- 34. Серологическое тестирование стадий инфекционного заболевания
- 36. ПРОТИВОПРОТОЗОЙНЫЙ ИММУНИТЕТ инвазия сопровождается формированием гуморального и
- 37. ИММУННАЯ ЗАЩИТА ПРОТИВ ГЕЛЬМИНТОВ компоненты
- 38. ОСОБЕННОСТИ ПРОТИВОГЛИСТНОГО ИММУНИТЕТА Ведущая роль - ЭОЗ,
- 39. Противогрибковый иммунитет Основой противогрибкового иммунитета является клеточный
- 40. Часть 2 АУТОИММУННЫЕ ПРОЦЕССЫ.
- 41. Аутоиммунные заболевания: - это заболевания, в патогенезе
- 42. Аутоиммунные заболевания: Эффекторные процессы аутоиммунных заболеваний реализуются
- 43. Естественные (нормальные) ауто-АТ вырабатываются в норме в
- 44. Естественные ауто -АТ, наряду с гормонами ,нейромедиаторами,
- 45. АУТОРЕАКТИВНЫЕ КЛОНЫ ЛИМФОЦИТОВ. В норме у каждого
- 46. ИММУНОЛОГИЧЕСКАЯ ТОЛЕРАНТНОСТЬ - это отсутствие активации
- 47. Факторы, обеспечивающие предрасположенность к аутоиммунным процессам Присутствие
- 48. Ассоциация аутоиммунных заболеваний с определенными антигенами МНС
- 49. МЕХАНИЗМЫ ЗАПУСКА АУТОИММУННОЙ ПАТОЛОГИИ. Нарушение функции регуляторных
- 50. МЕХАНИЗМЫ ЗАПУСКА АУТОИММУННОЙ ПАТОЛОГИИ. Микробные суперантигены вызывают
- 51. СОМАТИЧЕСКИЕ КЛЕТКИ СТАНОВЯТСЯ АПК В норме клетки
- 52. Нарушение изоляции «иммунологически привилегированных» органов и тканей:
- 53. АНТИГЕННАЯ МИМИКРИЯ. У бактерий имеются АГ-детерминанты, перекрестно
- 54. МИМИКРИЯ: МИКРОБНЫЕ И АУТО-АГ В РАЗВИТИЕ АЗ
- 55. МОДИФИКАЦИЯ СТРУКТУРЫ СОБСТВЕННЫХ БЕЛКОВ ОРГАНИЗМА.
- 56. Иммунологические механизмы аутоиммунных поражений гуморального типа накопление
- 57. Классификация аутоиммунных заболеваний В настоящее время нет единой классификации аутоиммунных заболеваний
- 58. Системные аутоиммунные заболевания - когда поражаются различные органы
- 59. Диффузный токсический зоб характеризуется триадой — гипертиреоидизм, зоб
- 60. Аутоиммунные заболевания суставов Ревматоидный артрит
- 61. Аутоиммунные заболевания крови Аутоиммунная гемолитическая анемия Аутоиммунная нейтропения Аутоиммунная тромбоцитопения Тромбоцитопеническая пурпура Аутоиммунная тромбоцитопения
- 62. Аутоиммунные заболевания кожи Витилиго Пемфигоид Изолированные
- 63. Аутоиммунные заболевания нервной системы Множественный или рассеянный
- 64. Классификация аутоиммунных заболеваний (по механизму иммунопатологического процесса)
- 65. Классификация аутоиммунных заболеваний (по механизму иммунопатологического процесса) - продолжение
- 66. Классификация аутоиммунных заболеваний (по механизму иммунопатологического процесса) - продолжение
- 67. Отличия между системными и органоспецифическими аутоиммунными заболеваниями.
- 68. Противоопухолевый иммунитет Часть 3 Онкопатология занимает
- 70. ОПУХОЛИ (незрелые, гетерологичные) состоят из умеренно- и
- 71. Опухоли способны возникать спонтанно, без видимых на
- 72. Противоопухолевые иммунные факторы Клеточные:
- 73. ИММУННЫЙ НАДЗОР ОПУХОЛЕВОГО РОСТА Концепция была предложена
- 74. Антигены, ассоциированные с опухолями
- 75. АНТИГЕНЫ, АССОЦИИРОВАННЫЕ С ОПУХОЛЯМИ раково-эмбриональный антиген
- 76. Противоопухолевый иммунитет основан на Тh1-зависимом клеточном
- 78. МЕХАНИЗМЫ ИЗБЕГАНИЯ ОПУХОЛЯМИ ИММУННОГО НАДЗОРА 1.
- 79. МЕХАНИЗМЫ ИЗБЕГАНИЯ ОПУХОЛЯМИ ИММУННОГО НАДЗОРА 3.Гены
- 80. МЕХАНИЗМЫ ИЗБЕГАНИЯ ОПУХОЛЯМИ ИММУННОГО НАДЗОРА 6.Противоопухолевые
- 81. Пробластомные факторы подавляют иммунитет 1. Супресcивные
- 82. ЦИТОКИНСИНТЕТИЧЕСКАЯ АКТИВНОСТЬ ОПУХОЛЕЙ. ДЕЙСТВИЕ ЦИТОКИНОВ НА ОПУХОЛИ.
- 85. Иммунодиагностика рака-МЕТОД ИФА основана на выявлении
- 86. Иммунотерапия больных с онкопатологией ведется по
- 87. ИММУНОТЕРАПИЯ ПРИ ЗЛОКАЧЕСТВЕННЫХ НОВООБРАЗОВАНИЯХ
- 90. Схема действия «мгновенного иммунитета». Выработавшиеся в ответ


Слайд 3
ПРОТИВОИНФЕКЦИОННЫИ ИММУНИТЕТ
– первая область приложения иммунологических принципов
Особенности иммунного ответа определяются:
путями поступления
и распространения инфекционных агентов
локализацией инфекционных агентов вне- или внутри клеток
особенностями патогенов микроорганизмов
модификацией активности иммунной системы компонентами патогенов.
локализацией инфекционных агентов вне- или внутри клеток
особенностями патогенов микроорганизмов
модификацией активности иммунной системы компонентами патогенов.

Слайд 4ФОРМИРОВАНИЕ ПРОТИВОИНФЕКЦИОННОГО ИММУНИТЕТА :
По направленности выделяют:
антитоксический,
антибактериальный,
противовирусный,
антигрибковый,
противопаразитарный
ПО МЕХАНИЗМУ

Слайд 5ПУТИ ПОСТУПЛЕНИЯ И РАСПРОСТРАНЕНИЯ ИНФЕКЦИОННЫХ АГЕНТОВ
определяют:
первичную локализацию иммунного процесса
преобладающие механизмы иммунного
ответа
Первая линия обороны-иммунная система слизистых
( дыхательной, пищеварительной и мочеполовой систем).
Проникновение патогена в субэпителиальное пространство возможно:
при адгезии патогена на клетках эпителия.
через поврежденные барьеры, дефекты эпителия.
«первичный очаг» инфекции- место проникновения.

Слайд 6КЛЮЧЕВОЙ ФАКТОР ОПРЕДЕЛЕНИЯ ТИПА ИММУННОГО ОТВЕТА
Если инфекционный агент обладает тропностью к
конкретным типам клеток (харатерно для внутриклеточных агентов) – все дальнейшие процессы ограничиваются органами, в которые он поступил.
Если агент внеклеточный и не обладает тропностью – он имеет склонность к распространению.
Если агент внеклеточный и не обладает тропностью – он имеет склонность к распространению.



Слайд 9ДВЕ ТРИАДЫ ОБЪЕДИНЯЮЩИЕ ВРОЖДЕННЫЙ И ПРИОБРЕТЕННЫЙ ИММУНИТЕТ
Врожденный
Приобретенный
НГ
МФ
КОМПЛИМЕНТ
ЦИТОКИНЫ
В-Лф
АНТИТЕЛА
Т-ЛФ
ГУМАРАЛЬНЫЙ
ИММУНИТЕТ
КЛЕТОЧНЫЙ
ИММУНИТЕТ
ВНЕКЛЕТОЧНЫЕ
ОРГАНИЗМЫ
ВНУТРИКЛЕТОЧНЫЕ
ОРГАНИЗМЫ

Слайд 10
ВРОЖДЕННЫЙ ИММУНИТЕТ
ПРИ ВНЕКЛЕТОЧНЫХ ИНФЕКЦИЯХ
Фагоциты (НГ, МФ), комплемент, БОВ , местная
воспалительная реакция, в меньшей степени NK-клетки.
Фагоциты
в начале этапа их эффективность невелика:
число клеток в месте внедрения недостаточно, они не активированы, их возможности в отношении распознавания и разрушения микробов ограничены,
при наличии в организме естественных АТ к м/о, эффективность фагоцитоза значительно повышается.
Комплемент.
Активируется по альтернативному пути:
на поверхности микробных клеток отсутствуют DAF (мембранный белок (Decay–Accelerating Factor — фактор), ускоряющий деградацию –ингибитор комплемента), пропердин стабилизирует образующиеся комплексы СЗb—Вb.
Повышение количества фиксированных на поверхности микробной клетки фрагментов СЗb способствует распознаванию их фагоцитами.
Фагоциты
в начале этапа их эффективность невелика:
число клеток в месте внедрения недостаточно, они не активированы, их возможности в отношении распознавания и разрушения микробов ограничены,
при наличии в организме естественных АТ к м/о, эффективность фагоцитоза значительно повышается.
Комплемент.
Активируется по альтернативному пути:
на поверхности микробных клеток отсутствуют DAF (мембранный белок (Decay–Accelerating Factor — фактор), ускоряющий деградацию –ингибитор комплемента), пропердин стабилизирует образующиеся комплексы СЗb—Вb.
Повышение количества фиксированных на поверхности микробной клетки фрагментов СЗb способствует распознаванию их фагоцитами.
, комплемент, БОВ , местная воспалительная реакция,")
Слайд 11Индукция воспалительного ответа.
дегрануляция тучных клеток, выделение хемотаксических , провоспалительных факторов
активация НГ,МФ
факторами инфекционных агентов,
активация эндотелиальных клеток продуктами МФ, повышение проницаемости сосудов LTB4 и гистамином, экспрессия молекул адгезии и синтез цитокинов,
,
миграция в очаг поражения большого числа - НГ,МОН, NK, а затем и ЛФ
усиление синтеза гепатоцитами БОФ(СРБ, амилоида, МБЛ, др.),
активация мигрировавших клеток и выделение ИФНγ-1 и ИЛ-6, ФНОα и хемокинов
активация комплемента по классическому и лектиновому пути

Слайд 12РАННИЙ ОТВЕТ ПРИ ВНЕКЛЕТОЧНЫХ ИНФЕКЦИЯХ
Осуществляются в первые 4
суток, локально.
участники: Ig, вырабатываемые B1-лф(CD5+), тимуснезависимые В-л и γδ+Т-л.
на общие РАМР разных бактерий вырабатываются низкоафинные АТ- Ig классов A, M, G2 ( В-л, без помощи Т-л).
Тимуснезависимый ответ В-л осуществляется в региональных ЛУ и групповых лимфатических фолликулах.
из-за неучастия Т-клеток:
отсутствует переключение изотипов
низкий аффинитет АТ ограничивает их защитную активность
отсутствует формирование иммунологической памяти.
IgA блокирует прикрепление бактерий к эпителиальным клеткам, но не способны активировать комплемент.
γδ+-Т-клетки в слизистых распознают ограниченный круг АГ (стрессорные белки микобактерий , пораженных клеток хозяина), путем связывания рецептором TCR без участия АПК. Механизм действия γδ+-клеток состоит в цитолизе инфицированных клеток
участники: Ig, вырабатываемые B1-лф(CD5+), тимуснезависимые В-л и γδ+Т-л.
на общие РАМР разных бактерий вырабатываются низкоафинные АТ- Ig классов A, M, G2 ( В-л, без помощи Т-л).
Тимуснезависимый ответ В-л осуществляется в региональных ЛУ и групповых лимфатических фолликулах.
из-за неучастия Т-клеток:
отсутствует переключение изотипов
низкий аффинитет АТ ограничивает их защитную активность
отсутствует формирование иммунологической памяти.
IgA блокирует прикрепление бактерий к эпителиальным клеткам, но не способны активировать комплемент.
γδ+-Т-клетки в слизистых распознают ограниченный круг АГ (стрессорные белки микобактерий , пораженных клеток хозяина), путем связывания рецептором TCR без участия АПК. Механизм действия γδ+-клеток состоит в цитолизе инфицированных клеток

Слайд 13ФОРМИРОВАНИЕ ПОЛНОЦЕННОГО АДАПТИВНОГО ИММУННОГО ОТВЕТА ПРИ ВНЕКЛЕТОЧНЫХ ИНФЕКЦИЯХ.
Первый этап: Гуморальный иммунный
ответ (не менее 4 дней).
Перемещение клеток между первичным инфекционным очагом и региональным лимфоидным органом.
В слизистых оболочках АПК – ДК, В-л,( не классические ТК) поэтому выбор ИО «решается» в пользу гуморального Th2-зaвисимого.
образуются IgG1 и IgG3-АТв региональных ЛУ и особенно в селезенке,
Перемещение клеток между первичным инфекционным очагом и региональным лимфоидным органом.
В слизистых оболочках АПК – ДК, В-л,( не классические ТК) поэтому выбор ИО «решается» в пользу гуморального Th2-зaвисимого.
образуются IgG1 и IgG3-АТв региональных ЛУ и особенно в селезенке,
Направленность действия АТ:
блокируют прикрепление бактерий- связывая компоненты фимбрий и капсул, липотейхоевые кислоты
инициируют комплемент зависимые реакции . АТ против липопротеинов капсул грам(-) бактерий и приводят к нарушению их целостности.
подавляют размножение- блокируют рецепторы для поглощения питательных веществ.
восстанавливают фагоцитоз связывая бактериальные иммунорепелленты.
препятствуют повреждению тканей. АТ против токсинов и гиалуронидазы
опсонизируя клетки-мишени Ig вызывают активацию NK-клеток и МФ, комплемента, нейтрализацию экзотоксинов.

")
Слайд 15Второй этап: Th1-зависимый клеточный ответ.
При внеклеточной инфекции – макрофагальный.
на 8-9 сутки
в процесс вовлекаются αβTCR-Т-лф при контакте с АПК в региональных ЛУ.
Th1- выделяют ИФН-γ, который, активируя МФ, в очаге поражения, способствует уничтожению фагоцитированных и цитолизу инфицированных клеток.
патоген первоначально перемещается лимфогенным путем.
Регионарный ЛУ- место формирования первичного (регионального) ИО и барьер на пути распространения инфекции.
Преодоление этого барьера приводит:
к гематогенному распространению инфекции, септицемии и генерализации распространения микроорганизма,
формированию вторичных очагов в тканях,
расселению активированных иммунных клеток по организму, системной активации лимфоидной ткани, вовлечению в ИО селезенки и других лимфоидных образований;
иногда к ареактивности вследствие интоксикации и избытка АГ.
Th1- выделяют ИФН-γ, который, активируя МФ, в очаге поражения, способствует уничтожению фагоцитированных и цитолизу инфицированных клеток.
патоген первоначально перемещается лимфогенным путем.
Регионарный ЛУ- место формирования первичного (регионального) ИО и барьер на пути распространения инфекции.
Преодоление этого барьера приводит:
к гематогенному распространению инфекции, септицемии и генерализации распространения микроорганизма,
формированию вторичных очагов в тканях,
расселению активированных иммунных клеток по организму, системной активации лимфоидной ткани, вовлечению в ИО селезенки и других лимфоидных образований;
иногда к ареактивности вследствие интоксикации и избытка АГ.

Слайд 16ОСОБЕННОСТИ ПАТОГЕНОВ МИКРООРГАНИЗМОВ.
Патогенные свойства м/о во многом зависят от типа оболочки.
бактерии можно поделить на:
Грамположительные
Грамотрицательные
Микобактерии
Спирохеты
В клеточной стенке бактерий присутствует пептидогликановый слой
у грам(-) бактерий он дополнительно покрыт внешним слоем, содержащим липополисахариды.
Поверхностные молекулы патогенов – выступают в роли «образов патогенности» - РАМР и распознаются ТLR рецепторами

Слайд 17ЭНДОТОКСИНЫ ЛИПОПОЛИСАХАРИДНОЙ ПРИРОДЫ ГРАМ(-):
активируют комплемент, к которому у грам(-) бактерий нет
резистентности.
вызывают образование фибрина, что нарушает подвижность бактерий.
распознают CD14, экспрессируемые НГ, ДК, МФ и В-л.
При этом увеличивается:
эффективность презентации АГ, активируется кооперация Т- и В-л.
продукция провоспалительных – ИЛ-1, ФНО-α, ИЛ-6, хемокинов
В малых дозах липопротеин работает как ко-стимулятор - адъювант и полезен для раннего ответа
Высокая его концентрация может привести к поликлональной стимуляции.
Иммунный ответ при этом:
«распыляется» на синтез АТ разнообразной специфичности (поликлональная стимуляция конкурирует с клоноспецифическим ответом),
чреват опасностью развития аутоиммунного процесса.
поликлональная стимуляция – важный фактор антимикробной защиты, поскольку проявляется в первые 4 суток (до того, как сформируется полноценный адаптивный ответ).
вызывают образование фибрина, что нарушает подвижность бактерий.
распознают CD14, экспрессируемые НГ, ДК, МФ и В-л.
При этом увеличивается:
эффективность презентации АГ, активируется кооперация Т- и В-л.
продукция провоспалительных – ИЛ-1, ФНО-α, ИЛ-6, хемокинов
В малых дозах липопротеин работает как ко-стимулятор - адъювант и полезен для раннего ответа
Высокая его концентрация может привести к поликлональной стимуляции.
Иммунный ответ при этом:
«распыляется» на синтез АТ разнообразной специфичности (поликлональная стимуляция конкурирует с клоноспецифическим ответом),
чреват опасностью развития аутоиммунного процесса.
поликлональная стимуляция – важный фактор антимикробной защиты, поскольку проявляется в первые 4 суток (до того, как сформируется полноценный адаптивный ответ).
:активируют комплемент, к которому у грам(-) бактерий нет резистентности.вызывают образование фибрина, что")
Слайд 18ПЕПТИДОГЛИКАНЫ БАКТЕРИАЛЬНОЙ СТЕНКИ - МУРАМИЛДИПЕПТИДЫ (ГРАМ +).
Активно влияют на МФ:
индуцируют выработку провоспалительных ИЛ и повышают их бактерицидную активность. Активированные МФ стимулируют иммунные реакции, реализуемые Т-, В- и NK-клетками, возможно прямое действие на Т-л.
Пептидогликаны слабо активируют комплемент.
Мурамилдипептиды лишены токсического компонента, препараты на их основе используют в клинической практике в качестве имунномодуляторов (Ликопид,гликопин).
Полисахариды с повторяющимися эпитопами- являются тимуснезависимыми АГ( например, оболочки пневмококков). Однако некоторые бактериальные полисахариды способны индуцировать анергию В-л.
Гликолипиды оболочки микобактерий обладают адьювантным эффектом, стимулируют фагоцитоз.
Пептидогликаны слабо активируют комплемент.
Мурамилдипептиды лишены токсического компонента, препараты на их основе используют в клинической практике в качестве имунномодуляторов (Ликопид,гликопин).
Полисахариды с повторяющимися эпитопами- являются тимуснезависимыми АГ( например, оболочки пневмококков). Однако некоторые бактериальные полисахариды способны индуцировать анергию В-л.
Гликолипиды оболочки микобактерий обладают адьювантным эффектом, стимулируют фагоцитоз.
. Активно влияют на МФ: индуцируют выработку провоспалительных ИЛ")
Слайд 19ПУТИ ЗАЩИТЫ МИКРООРГАНИЗМОВ ОТ ИММУННОГО ОТВЕТА
1.Вариабельность.
За счет мутаций внутри вида возникают
различные серологические варианты (типы и субтипы). При инфицировании формируется иммунитет только к линии возбудителя, но не ко всем его разновидностям. Характерна для стрептококка, м/о кишечной группы.
Если специфичность АГ изменяется в инфицированном организме – происходит обострение заболевания после временного ослабления симптоматики.
2.Способность избегать разрушающего действия комплемента
Особенности капсулы грам(+) бактерии.
С3b связывается в отдалении от чувствительной к комплементу оболочки.
E.Coli, Salmonella, Pseudomonas способны «сбрасывать» комплекс С5b-С9,
Конфигурация наружной поверхности препятствует контакту фагоцита с С3b.
Бактерии выделяют «белки-ловушки», связывающие комплемент.
Если специфичность АГ изменяется в инфицированном организме – происходит обострение заболевания после временного ослабления симптоматики.
2.Способность избегать разрушающего действия комплемента
Особенности капсулы грам(+) бактерии.
С3b связывается в отдалении от чувствительной к комплементу оболочки.
E.Coli, Salmonella, Pseudomonas способны «сбрасывать» комплекс С5b-С9,
Конфигурация наружной поверхности препятствует контакту фагоцита с С3b.
Бактерии выделяют «белки-ловушки», связывающие комплемент.

Слайд 20СРЕДИ ФАКТОРОВ, ПОЗВОЛЯЮЩИХ М/О ИЗБЕЖАТЬ ЗАЩИТНЫХ МЕХАНИЗМОВ, ВЕЩЕСТВА, ИЗМЕНЯЮЩИЕ НАПРАВЛЕНИЕ ИО,
И СОВСЕМ НЕМНОГО ЧИСТЫХ ИММУНОСУПРЕССАНТОВ.
3.Компоненты микроорганизмов влияют на фагоцитоз.
подавляют хемотаксис (репелленты)
препятствуют связыванию с фагоцитом ( гиалуроновая кислота грам(+), белки шипов нейссерий, фимбрий стрептококков)
препятствуют слиянию фагосом с лизосомами, могут размножаться внутриклеточно, блокируют способность фагоцита активироваться ИФН (Микобактерии)
способны инактивировать свободные радикалы (Мycobacterium leprae).
4.Белки бактерий могут связывать иммуноглобулины.
Белок-А клеточной стенки Staphylococcus aureus связываясь с Fc-фрагментом к IgG, препятствует взаимодействию IgG с эффекторными клетками.
5.Экзотоксины (суперантигены ) вызвают поликлональную гиперактивацию Т-л, а затем их массовую гибель.

Слайд 21ИММУННЫЙ ОТВЕТ ПРИ ВНУТРИКЛЕТОЧНОЙ ИНФЕКЦИИ
Вирусы – облигатные внутриклеточные паразиты, использующие для
воспроизводства ресурсы клетки хозяина.
Вирусы прикрепляются к клеткам, связываясь со специфическими рецепторами, что определяет тропизм вируса к определенным тканям.
Вирусы прикрепляются к клеткам, связываясь со специфическими рецепторами, что определяет тропизм вируса к определенным тканям.
Пути проникновения вирусов в клетки.

Слайд 22ВРОЖДЕННЫЙ ИММУНИТЕТ ПРИ ВНУТРИКЛЕТОЧНЫХ ИНФЕКЦИЯХ
Первые часы после внедрения (внеклеточная фаза) защиту
обеспечивают фагоциты и NK-клетки.
Их мало , эффективность невелика, повышается при наличии в организме естественных АТ к вирусам.
Некоторые вирусы могут активировать комплемент, но он не играет роли в противовирусной защите.
Клетки, инфицированные вирусом, синтезируют ИФНα и ИФНβ
Интерфероны делают соседние клетки устойчивыми к вирусу:
активируя протеинкиназу, которая блокирует синтез вирусного белка,
активируя эндонуклеазу, которая способна разрушить вирусную РНК.
Через 2-е суток появляются активные NK-клетки.
При герпесвирусной (в частности цитамегаловирусной) инфекции это главный эффекторный механизм, так указанные вирусы подавляют экспрессию MHC-1.
Их мало , эффективность невелика, повышается при наличии в организме естественных АТ к вирусам.
Некоторые вирусы могут активировать комплемент, но он не играет роли в противовирусной защите.
Клетки, инфицированные вирусом, синтезируют ИФНα и ИФНβ
Интерфероны делают соседние клетки устойчивыми к вирусу:
активируя протеинкиназу, которая блокирует синтез вирусного белка,
активируя эндонуклеазу, которая способна разрушить вирусную РНК.
Через 2-е суток появляются активные NK-клетки.
При герпесвирусной (в частности цитамегаловирусной) инфекции это главный эффекторный механизм, так указанные вирусы подавляют экспрессию MHC-1.
 защиту обеспечивают фагоциты и NK-клетки.")
Слайд 23
Индукция воспалительного ответа.
На 2-4 сутки, запускается воспалительный ответ.
При этом происходит:
активация
эндотелиальных клеток продуктами МФ,
повышение проницаемости сосудов,
экспрессия молекул адгезии и синтез цитокинов,
миграция в очаг поражения большого числа — НГ, МОН, а затем и ЛФ,
активация мигрировавших клеток и выделение ИЛ-1 и ИЛ-6, ФНОα, хемокинов.
усиление синтеза гепатоцитами БОФ(С-реактивного белка, сывороточного амилоида, маннозосвязывающего белка),
выработка инфицированными клетками ИФН α и β͢ => активация NK-клеток.
повышение проницаемости сосудов,
экспрессия молекул адгезии и синтез цитокинов,
миграция в очаг поражения большого числа — НГ, МОН, а затем и ЛФ,
активация мигрировавших клеток и выделение ИЛ-1 и ИЛ-6, ФНОα, хемокинов.
усиление синтеза гепатоцитами БОФ(С-реактивного белка, сывороточного амилоида, маннозосвязывающего белка),
выработка инфицированными клетками ИФН α и β͢ => активация NK-клеток.

Слайд 24АДАПТИВНЫЙ ИММУННЫЙ ОТВЕТ ПРИ ВНУТРИКЛЕТОЧНЫХ ИНФЕКЦИЯХ Th1- ЗАВИСИМЫЙ КЛЕТОЧНЫЙ
Начинает формироваться в
первые дни, пик достигается через 3-4 недели.
протекает с преобладанием цитотоксического варианта:
Th1 синтезируют ИЛ-2,ИФН-γ активируя CD8+ -клетки (ТЦЛ).
CD8+Т-лф покидают ЛУ и мигрируют в воспаленные участки.
В очаге Т-киллеры убивают инфицированные клетки, несущие на своей поверхности вирусные АГ, ассоциированные с MHC-1.
Th1-клетки выделяют ИФН-γ, который:
активирует NK-клеток и МФ, для цитолиза инфицированных клеток уничтожения фагоцитированных клеток (основной механизм защиты от ВПГ1-типа),
препятствует инфицированию клеток,
ингибирует размножение вируса в клетке,
усиливает экспрессию MHC-1 и MHC-2 типа.
протекает с преобладанием цитотоксического варианта:
Th1 синтезируют ИЛ-2,ИФН-γ активируя CD8+ -клетки (ТЦЛ).
CD8+Т-лф покидают ЛУ и мигрируют в воспаленные участки.
В очаге Т-киллеры убивают инфицированные клетки, несущие на своей поверхности вирусные АГ, ассоциированные с MHC-1.
Th1-клетки выделяют ИФН-γ, который:
активирует NK-клеток и МФ, для цитолиза инфицированных клеток уничтожения фагоцитированных клеток (основной механизм защиты от ВПГ1-типа),
препятствует инфицированию клеток,
ингибирует размножение вируса в клетке,
усиливает экспрессию MHC-1 и MHC-2 типа.


Слайд 26ГУМОРАЛЬНЫЙ АДАПТИВНЫЙ ИММУННЫЙ ОТВЕТ ПРИ ВНУТРИКЛЕТОЧНЫХ ИНФЕКЦИЯХ
АТ при вирусной инфекции вырабатываются
на белки и гликопротеины оболочки (тимусзависимые АГ).
развивается медленно, но вследствие участия Тh обеспечивается:
высокий аффинитет АТ (за счет усиления мутагенеза),
формирование иммунологической памяти.
Функции специфических АТ при вирусной инфекции:
главное препятствие внеклеточного распространения вируса,
участие в реакциях антителозависимой цитотоксичности (АЗКЦ) против инфицированных клеток,
активации комплемента для разрушения инфицированных клеток.
развивается медленно, но вследствие участия Тh обеспечивается:
высокий аффинитет АТ (за счет усиления мутагенеза),
формирование иммунологической памяти.
Функции специфических АТ при вирусной инфекции:
главное препятствие внеклеточного распространения вируса,
участие в реакциях антителозависимой цитотоксичности (АЗКЦ) против инфицированных клеток,
активации комплемента для разрушения инфицированных клеток.

Слайд 27СТРАТЕГИЯ ОБХОДА ВИРУСАМИ ИММУНОЛОГИЧЕСКОГО КОНТРОЛЯ
1.Вариабельность - делает невозможным формирование защитного иммунитета
и очень затрудняет вакцинопрофилактику.
Типы вариабельности:
Антигенный дрейф (drift). Вследствие серии точечных мутаций накапливаются изменения специфичности АГ. Сформировавшийся протективный механизм срабатывает лишь частично.
Антигенный сдвиг (shift) – следствие обмена генетическим материалом между разными линиями. Изменения специфичности АГ происходят скачкообразно. Сформировавшийся иммунитет не распространяется на измененные варианты.
Пример: Вариабельность гемагглютинина вируса гриппа А
Время, требующееся для изменения типа АГ у вирусов гриппа:
для гемагглютинина — от одной эпидемии к следующей,
для нейраминидазы — через 2—3 эпидемии;
При этом спустя некоторое время (десятки лет, 3—5 эпидемий) наблюдается возвращение ранее уже зарегистрированного серотипа гемагглютинина.
Типы вариабельности:
Антигенный дрейф (drift). Вследствие серии точечных мутаций накапливаются изменения специфичности АГ. Сформировавшийся протективный механизм срабатывает лишь частично.
Антигенный сдвиг (shift) – следствие обмена генетическим материалом между разными линиями. Изменения специфичности АГ происходят скачкообразно. Сформировавшийся иммунитет не распространяется на измененные варианты.
Пример: Вариабельность гемагглютинина вируса гриппа А
Время, требующееся для изменения типа АГ у вирусов гриппа:
для гемагглютинина — от одной эпидемии к следующей,
для нейраминидазы — через 2—3 эпидемии;
При этом спустя некоторое время (десятки лет, 3—5 эпидемий) наблюдается возвращение ранее уже зарегистрированного серотипа гемагглютинина.

Слайд 282.Модификация вирусами активности иммунной системы
а)Блокада активации комплемента, связывание АТ.
Гликопротеин ЦМВ,ВПГ, связывает
Fc-фрагмент IgG тем самым:
связываются противовирусные АТ, не происходит антителозависимый цитолиз
блокируется активация комплемента (не развивается МАК-зависимый цитолиз )
б)Торможение экспрессии МНС-1. ЦМВ, аденовирусы, блокируя экспрессию МНС-1, подавляют распознавание инфицированной клетки CD8+ лимфоцитами.
в)Синтез вирусами факторов взаимодействующих с цитокинами.
блокада эффекта ИФН( ВЭБ, аденовирусы)
вирус Миксомы вырабатывает гомолог рецептора ИФНγ, связывающий ИФН и препятствующий реализации его функции,
белок, секретируемый ВЭБ, гомологичен ИЛ-10 и воспроизводит его эффекты (подавление активности Th1 и переключение и.о. на гуморальный).
Вирусная двуспиральная РНК - мощный интерфероноген, повышает активность МФ и NK. Синтетические аналоги вирусной двуспиральной РНК (поли-I/поли-С, поли-А/поли-U) широко используются в качестве иммуномодуляторов.
связываются противовирусные АТ, не происходит антителозависимый цитолиз
блокируется активация комплемента (не развивается МАК-зависимый цитолиз )
б)Торможение экспрессии МНС-1. ЦМВ, аденовирусы, блокируя экспрессию МНС-1, подавляют распознавание инфицированной клетки CD8+ лимфоцитами.
в)Синтез вирусами факторов взаимодействующих с цитокинами.
блокада эффекта ИФН( ВЭБ, аденовирусы)
вирус Миксомы вырабатывает гомолог рецептора ИФНγ, связывающий ИФН и препятствующий реализации его функции,
белок, секретируемый ВЭБ, гомологичен ИЛ-10 и воспроизводит его эффекты (подавление активности Th1 и переключение и.о. на гуморальный).
Вирусная двуспиральная РНК - мощный интерфероноген, повышает активность МФ и NK. Синтетические аналоги вирусной двуспиральной РНК (поли-I/поли-С, поли-А/поли-U) широко используются в качестве иммуномодуляторов.
Блокада активации комплемента, связывание АТ.Гликопротеин ЦМВ,ВПГ, связывает Fc-фрагмент IgG тем самым:связываются")
Слайд 293.ОТКЛОНЕНИЯ ОТ «КЛАССИЧЕСКОГО» ВАРИАНТА ИММУННОГО ОТВЕТА ПРИ ИНФЕКЦИЯХ
Особенности патогенов или
ИС могут приводить:
к повреждению собственных тканей
длительному персистированию возбудителя
хронизации процесса.
Повреждение собственных тканей
При персистировании инфекции избыток количества АГ относительно количества АТ приводит к образованию слабо растворимых иммунных комплексов (ЦИК) – в результате иммунокомплексные заболевания.(Вирус гепатита В, вирус лимфоцитарного хориоменингита)
Клетки ЦНС, инфицированные вирусом лимфоцитарного хориоменингита, поражаются собственными цитотоксическими лимфоцитами.
быстрое и обильное высвобождение цитокинов индуцируемое суперантигенами, вызывает потенциально смертельное повреждение – эндотоксический шок. Чаще развивается при инфицировании грам(-) бактериями и стафилококком.
к повреждению собственных тканей
длительному персистированию возбудителя
хронизации процесса.
Повреждение собственных тканей
При персистировании инфекции избыток количества АГ относительно количества АТ приводит к образованию слабо растворимых иммунных комплексов (ЦИК) – в результате иммунокомплексные заболевания.(Вирус гепатита В, вирус лимфоцитарного хориоменингита)
Клетки ЦНС, инфицированные вирусом лимфоцитарного хориоменингита, поражаются собственными цитотоксическими лимфоцитами.
быстрое и обильное высвобождение цитокинов индуцируемое суперантигенами, вызывает потенциально смертельное повреждение – эндотоксический шок. Чаще развивается при инфицировании грам(-) бактериями и стафилококком.

Слайд 304.Неадекватная тактика иммунного ответа
Th1-зависимые клеточные механизмы
эффективны при внутриклеточных инфекциях: туберкулезе,
листериозе, лейшманиозе, проказе, столбняке, вирусных инфекциях (грипп, ВИЧ), кандидозе.
Пример: при инфицировании М. Leprae если ответ развивается
по Тh1- типу – развивается туберкулоидная форма лепры, имеющая благоприятный прогноз
По Тh2- пути – проказа проявляется в лепроматозной форме, не поддающейся излечению
Th2-зависимый гуморальный путь эффективен при внеклеточных инфекциях: бореллиозе, гельминтозах, нейссериями, стафилококками, микоплазмами, кишечными палочками, частично вирусных инфекциях.
Пример: при инфицировании Leishmania maior ответ развивается
по Тh1-зависимому типу – полная защита
по Тh2-зависимому пути – болезнь со смертельным исходом,.
Механизмы формирования неадекватного ответа неясны
за редкими исключениями:
Вирус Эпштейна-Барр продуцирует фактор BCR F-1 с активностью подобной ИЛ-10, направляющий ответ по Th2-пути неадекватному для защиты от вирусной инфекции.

Слайд 315.Переход вируса в латентную фазу.
При этом происходит:
Прекращение репликации вируса и
формирования новых вирусных частиц=>>
вирусный АГ перестает экспрессироваться на поверхности клеток.
Патоген становится недосягаемым для иммунных факторов.
Иммунный процесс затухает до следующей фазы активации вируса.
Вирус гепатита В и Эпштейна-Барр, вируса ветряной оспы, опоясывающего лишая, простого герпеса персистируют с постоянным или периодическим выделением активного вируса.
Прионы особый класс инфекционных агентов- белки с аномальной третичной структурой и не содержащих нуклеиновых кислот. Прионы увеличивают свою численность, используя функции живых клеток (в этом схожи с вирусами).
Прион -способен катализировать превращение гомологичного ему нормального клеточного белка в себе подобный (болезнь Скрейпи и Крейцфельда-Якоба), персистируют, не вызывая иммунного ответа.
вирусный АГ перестает экспрессироваться на поверхности клеток.
Патоген становится недосягаемым для иммунных факторов.
Иммунный процесс затухает до следующей фазы активации вируса.
Вирус гепатита В и Эпштейна-Барр, вируса ветряной оспы, опоясывающего лишая, простого герпеса персистируют с постоянным или периодическим выделением активного вируса.
Прионы особый класс инфекционных агентов- белки с аномальной третичной структурой и не содержащих нуклеиновых кислот. Прионы увеличивают свою численность, используя функции живых клеток (в этом схожи с вирусами).
Прион -способен катализировать превращение гомологичного ему нормального клеточного белка в себе подобный (болезнь Скрейпи и Крейцфельда-Якоба), персистируют, не вызывая иммунного ответа.

Слайд 326.Непосредственное инфицирование ЛФ и МФ
Некоторые вирусы, поражая ИКК, приводят к иммунным
дефектам.
Инфекции, при которых вирус размножается в пролиферирующих лимфоцитах, к АГ вируса формируется иммунологическая толерантность.
Инфекции, при которых вирус размножается в пролиферирующих лимфоцитах, к АГ вируса формируется иммунологическая толерантность.
Формирование гранулемы
Гранулемы (при туберкулезе, проказе) – скопления активированных МФ, содержащих неубитые и размножающиеся микобактерии, окруженные активированными Т-л. Туберкулезные вирусные и некоторые другие инфекционные агенты могут вызывать повреждения мембран с последующей их «неправильной» репарацией, в результате происходит слияние клеток – формируется симпласт. Развитие гранулемы, как правило, служит показателем неэффективности иммунной защиты.

Слайд 337.Перекрестная реактивность АГ микро- и макроорганизма
При инфицировании стрептококком типа А появляются
АТ против β-D-N-ацетилглюкозамина – эпитопа, общего для стрептококка и клеток человека.
АТ реагируют с тканями хозяина, экспрессирующими этот полисахарид – миокардом, клапанами сердца, синовиальной оболочкой суставов, эпителиальными клетками – развитие ревматизма.
При инфицировании Treponema pallidum появляются АТ к кардиолипину, патогенетически значимые для формирования волчаночного синдрома.
При инфицировании Treponema cruzis имеет место перекрестная реакция с белками ЦНС. Вследствие развития аутоагрессии возникает поздняя нервная симптоматика.
АТ реагируют с тканями хозяина, экспрессирующими этот полисахарид – миокардом, клапанами сердца, синовиальной оболочкой суставов, эпителиальными клетками – развитие ревматизма.
При инфицировании Treponema pallidum появляются АТ к кардиолипину, патогенетически значимые для формирования волчаночного синдрома.
При инфицировании Treponema cruzis имеет место перекрестная реакция с белками ЦНС. Вследствие развития аутоагрессии возникает поздняя нервная симптоматика.

Слайд 34Серологическое тестирование стадий инфекционного заболевания
2-3-х кратное снижение специфических IgG, IgA в процессе лечения –
эффективность терапии!!
эффективность терапии!!
!!!

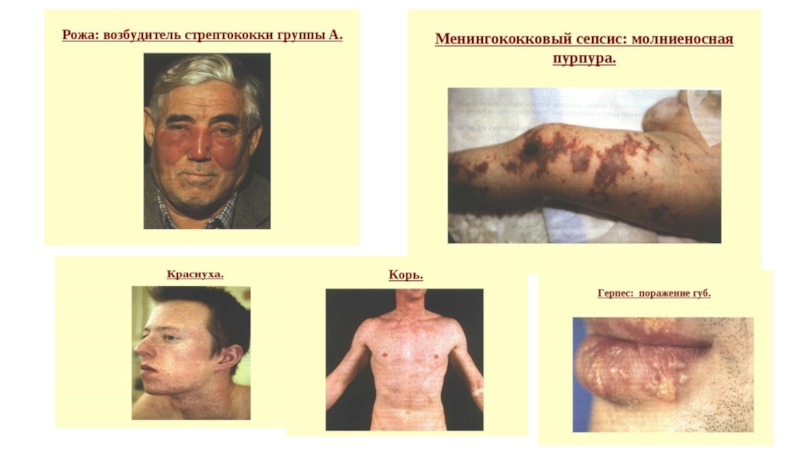
Слайд 36ПРОТИВОПРОТОЗОЙНЫЙ ИММУНИТЕТ
инвазия сопровождается формированием гуморального и клеточного иммунитета.
Характер противопаразитарного иммунитета
определяется биологическими особенностями паразита.
Антитела (IgМ, IgG) против простейших действуют на внеклеточные формы паразитов:
активируют АЗКЦ с участием МФ,
в случае внутриклеточного паразитирования участвуют – NK,NKTи γδT лимфоциты.
Паразитарные инвазии сопровождаются аллергизацией организма по механизму ГЗТ.
Многие паразиты обладают высокой АГ-изменчивостью, что позволяет им избегать действия факторов иммунитета.
Такие паразиты (малярийный плазмодий) как бы «ускользает» от ранее образовавшихся АТ
Фагоцитоз часто незавершенный (при лейшманиозах)
Выявление ГЗТ используют при диагностике токсоплазмоза, лейшманиоза и др. протозойных инфекций
Антитела (IgМ, IgG) против простейших действуют на внеклеточные формы паразитов:
активируют АЗКЦ с участием МФ,
в случае внутриклеточного паразитирования участвуют – NK,NKTи γδT лимфоциты.
Паразитарные инвазии сопровождаются аллергизацией организма по механизму ГЗТ.
Многие паразиты обладают высокой АГ-изменчивостью, что позволяет им избегать действия факторов иммунитета.
Такие паразиты (малярийный плазмодий) как бы «ускользает» от ранее образовавшихся АТ
Фагоцитоз часто незавершенный (при лейшманиозах)
Выявление ГЗТ используют при диагностике токсоплазмоза, лейшманиоза и др. протозойных инфекций

Слайд 37ИММУННАЯ ЗАЩИТА ПРОТИВ ГЕЛЬМИНТОВ
компоненты гельминтов (PAMP) распознаются РРR рецепторами клеток врожденного
иммунитета (МФ, ДК, энтероцитами слизистых оболочек кишечника).
лизофосфатидилсерин (распознается TLR2), липопротеины, обогащенные фосфорилхолином (распознаются TLR4), гликаны- распознаются лектиновыми рецепторами
активация ДК приводит к дифференцировке Th0 в Th2.
Th2-ответ- преобладающая форма протективного иммунного ответа против гельминтов.
ИЛ-4, ИЛ-13, ИЛ-5 обеспечивают переключение изотипов антител на IgE,
привлекают эозинофилы и базофилы.
IgE – взаимодействуют с сенсебилизированными тучными клетками, стимулируют выброс факторов с антигельминтной активностью и хемоатрактантами для ЭОЗ.
лизофосфатидилсерин (распознается TLR2), липопротеины, обогащенные фосфорилхолином (распознаются TLR4), гликаны- распознаются лектиновыми рецепторами
активация ДК приводит к дифференцировке Th0 в Th2.
Th2-ответ- преобладающая форма протективного иммунного ответа против гельминтов.
ИЛ-4, ИЛ-13, ИЛ-5 обеспечивают переключение изотипов антител на IgE,
привлекают эозинофилы и базофилы.
IgE – взаимодействуют с сенсебилизированными тучными клетками, стимулируют выброс факторов с антигельминтной активностью и хемоатрактантами для ЭОЗ.
 распознаются РРR рецепторами клеток врожденного иммунитета (МФ, ДК,")
Слайд 38ОСОБЕННОСТИ ПРОТИВОГЛИСТНОГО ИММУНИТЕТА
Ведущая роль - ЭОЗ, которые осуществляют АЗКЦ.
ЭОЗ распознают
паразитов, отмеченных специфическими IgE или IgA.
ЭОЗ и БАЗ образуют вал вокруг гельминтов и выделеляют молекулы и вещества обладающие антигельминтными свойствами:
белок ЭОЗ- МВР (главный щелочной белок)
белок ЭОЗ-ЕСР (эозинофильный катионный белок)
АГ гельминта, связываясь с рецепторами ТК слизистой оболочки, вызывают их дегрануляцию.
Экскретированные биологически активные соединения вызывают интенсивную перистальтику, удаляющую паразита или его останки из просвета кишки.
ЭОЗ и ТК синтезируют цитокины и липидные медиаторы, потенцирующие воспалительную реакцию в месте внедрения гельминта.
Глистная инвазия сопровождается аллергизацией в основном по механизму ГЗТ.
Против гельминтов формируется относительно слабая и кратковременная иммунологическая память.
ЭОЗ и БАЗ образуют вал вокруг гельминтов и выделеляют молекулы и вещества обладающие антигельминтными свойствами:
белок ЭОЗ- МВР (главный щелочной белок)
белок ЭОЗ-ЕСР (эозинофильный катионный белок)
АГ гельминта, связываясь с рецепторами ТК слизистой оболочки, вызывают их дегрануляцию.
Экскретированные биологически активные соединения вызывают интенсивную перистальтику, удаляющую паразита или его останки из просвета кишки.
ЭОЗ и ТК синтезируют цитокины и липидные медиаторы, потенцирующие воспалительную реакцию в месте внедрения гельминта.
Глистная инвазия сопровождается аллергизацией в основном по механизму ГЗТ.
Против гельминтов формируется относительно слабая и кратковременная иммунологическая память.
гибель гельминтов

Слайд 39Противогрибковый иммунитет
Основой противогрибкового иммунитета является клеточный иммунитет:
фагоцитоз→ эпителиоидная гранулематозная реакция→
ГЗТ → Аллергия →IgE-антитела против антигенов грибов → ГЗТ и ГНТ.
Антитела (IgM, IgG) при микозах выявляются в низких титрах.
Факторы противогрибковой неспецифической резистентности кожи, слизистых оболочек рта и слюны
Микозы, особенно, оппортунистические, часто развиваются после длительной антибиотикотерапии и при ИДС
Антитела (IgM, IgG) при микозах выявляются в низких титрах.
Факторы противогрибковой неспецифической резистентности кожи, слизистых оболочек рта и слюны
Микозы, особенно, оппортунистические, часто развиваются после длительной антибиотикотерапии и при ИДС


Слайд 41Аутоиммунные заболевания:
- это заболевания, в патогенезе которых ведущую роль играют аутореактивные
лимфоциты, которые распознают АГ собственного организма как чужеродные и запускают патологические процессы, характеризующиеся деструкцией -
клеток-мишеней
тканей-мишеней
нарушением их функций (как снижением, так и усилением)
развитием хронического воспаления.
клеток-мишеней
тканей-мишеней
нарушением их функций (как снижением, так и усилением)
развитием хронического воспаления.
Аутоиммунные заболевания поражают 5-7 % населения Земли.

Слайд 42Аутоиммунные заболевания:
Эффекторные процессы аутоиммунных заболеваний реализуются через гуморальные (ауто-АТ) и/или клеточные
(аутореактивные клоны лимфоцитов) иммунные реакции.
Гиперпродукция провоспалительных цитокинов сопровождает практически все аутоиммунные заболевания.
 и/или клеточные (аутореактивные клоны лимфоцитов) иммунные")
Слайд 43Естественные (нормальные) ауто-АТ
вырабатываются в норме в небольших количествах
синтез ауто-АТ начинается
во внутриутробном периоде – раньше синтеза АТ к чужеродным АГ.
CD5+-В1-лф - вырабатывают «естественные» АТ- IgM ,не индуцированные иммунизацией, без Тh поддержки, в том числе к некоторым ауто-АГ.
присутствуют нормальные ауто-АТ, реагирующие с ДНК, компонентами цитоскелета, миелином, сывороточным альбумином, тиреоглобулином, цитохромом, коллагеном, трансферрином, с компонентами клеток сердца, почек, эритроцитами, фибробластами, а также органоспецифические IgG-ауто-АТ. С возрастом титр этих АТ нарастает.
CD5+-В1-лф - вырабатывают «естественные» АТ- IgM ,не индуцированные иммунизацией, без Тh поддержки, в том числе к некоторым ауто-АГ.
присутствуют нормальные ауто-АТ, реагирующие с ДНК, компонентами цитоскелета, миелином, сывороточным альбумином, тиреоглобулином, цитохромом, коллагеном, трансферрином, с компонентами клеток сердца, почек, эритроцитами, фибробластами, а также органоспецифические IgG-ауто-АТ. С возрастом титр этих АТ нарастает.
В норме АТ обновляются ежедневно из них:
70-80% - АТ к инфекционным АГ
20-30% - ауто-АТ
Если продукция ауто-АТ к ауто-АГ выходит за физиологические пределы и изменения сохраняются месяцами- это может стать причиной развития аутоиммунных заболеваний (АИЗ).
 ауто-АТвырабатываются в норме в небольших количествах синтез ауто-АТ начинается во внутриутробном периоде –")
Слайд 44Естественные ауто -АТ, наряду с гормонами ,нейромедиаторами, регулируют множество физиологических функций.
Способствуют
освобождению организма от отмирающих клеток и других отживших свой срок биологических структур.
Взаимодействуя со специализированными ауто-АГ: рецепторами, гормонами, ферментативными комплексами естественные АТ служат:
стимуляторами роста и функции щитовидной железы, коры надпочечников,
агентами апоптоза,
регуляторами дифференцировки, стимуляторами регенерации,
являются факторами естественной радиопротекции.
Эффекты ауто-АТ не замещают и не превосходят действие нормальных эндокринных или нервных регуляторов (гормонов, медиаторов и прочих биоактивных веществ), а только дополняют их.
Взаимодействуя со специализированными ауто-АГ: рецепторами, гормонами, ферментативными комплексами естественные АТ служат:
стимуляторами роста и функции щитовидной железы, коры надпочечников,
агентами апоптоза,
регуляторами дифференцировки, стимуляторами регенерации,
являются факторами естественной радиопротекции.
Эффекты ауто-АТ не замещают и не превосходят действие нормальных эндокринных или нервных регуляторов (гормонов, медиаторов и прочих биоактивных веществ), а только дополняют их.

Слайд 45АУТОРЕАКТИВНЫЕ КЛОНЫ ЛИМФОЦИТОВ.
В норме у каждого здорового организма в периферических лимфоидных
тканях есть и T-л, и B-л с АГ-распознающими рецепторами для «своего»,
т.е. манифестация аутоиммунных заболеваний не является результатом возникновения аномальных аутореактивных клонов лимфоцитов - они всегда присутствуют.
В норме аутореактивные клоны находятся в состоянии иммунологической толерантности, и срыв этого состояния приводит к запуску аутоиммунного процесса, переходящего в заболевание.
т.е. манифестация аутоиммунных заболеваний не является результатом возникновения аномальных аутореактивных клонов лимфоцитов - они всегда присутствуют.
В норме аутореактивные клоны находятся в состоянии иммунологической толерантности, и срыв этого состояния приводит к запуску аутоиммунного процесса, переходящего в заболевание.
Чаще всего манифестация аутоиммунного деструктивного процесса инициируется патогенным внешним фактором.

Слайд 46ИММУНОЛОГИЧЕСКАЯ ТОЛЕРАНТНОСТЬ -
это отсутствие активации лимфоцитов (и, следовательно продукции ими
эффекторных молекул) при наличии специфического АГ.
Толерантность Лф (как отсутствие ответа на АГ) нужна только по отношению к АГ собственных неповрежденных тканей.
Толерантность Лф (как отсутствие ответа на АГ) нужна только по отношению к АГ собственных неповрежденных тканей.
Супрессия
Толерантность
Продуктивная активация клона ЛФ начинается, реализуется, затем подавляется
Продуктивной активации антигенспецифического клона ЛФ не происходит
Механизмы:
апоптоз
подавление внутриклеточного метаболизма сигналами с ингибирующих рецепторов
 при наличии")
Слайд 47Факторы, обеспечивающие предрасположенность к аутоиммунным процессам
Присутствие ауто-АТ или аутоспецифических клеток недостаточно
для развития аутоиммунного процесса.
Наследственный фактор. У ближайших родственников больных, даже в случаях отсутствия заболевания, обнаруживаются повышенный уровень ауто-АТ. Семейные заболевания чаще органоспецифические, наследуется не только предрасположенность, но и мишень (орган).
Генетическая предрасположенность сцеплена с генами МНС.
Пример: предрасположенность или резистентность к развитию инсулинзависимого диабета определяет различие по 1 остатку в позиции 57 молекулы HLA-DQ (резистентность обусловлена присутствием остатка аспарагиновой кислоты, предрасположенность – остатков валина, серина или аланина).
Среди негенетических факторов - пол (у женщин развиваются чаще) и возраст (с возрастом увеличивается вероятность развития заболевания).
Наследственный фактор. У ближайших родственников больных, даже в случаях отсутствия заболевания, обнаруживаются повышенный уровень ауто-АТ. Семейные заболевания чаще органоспецифические, наследуется не только предрасположенность, но и мишень (орган).
Генетическая предрасположенность сцеплена с генами МНС.
Пример: предрасположенность или резистентность к развитию инсулинзависимого диабета определяет различие по 1 остатку в позиции 57 молекулы HLA-DQ (резистентность обусловлена присутствием остатка аспарагиновой кислоты, предрасположенность – остатков валина, серина или аланина).
Среди негенетических факторов - пол (у женщин развиваются чаще) и возраст (с возрастом увеличивается вероятность развития заболевания).

Слайд 48Ассоциация аутоиммунных заболеваний с определенными антигенами МНС
В настоящее время диагностировано достаточно
много аутоиммунных заболеваний, у которых установлена ассоциация с аллелями МНС.

Слайд 49МЕХАНИЗМЫ ЗАПУСКА АУТОИММУННОЙ ПАТОЛОГИИ.
Нарушение функции регуляторных Т-лимфоцитов-Тreg (CD3+CD4+CD25+Foxp3+).
Срыв толерантности к ауто-АТ
(аутотолерантность) и возникновение аутоиммунного конфликта происходят из-за нарушения взаимодействия ИКК и механизмов регуляции их активности.
Поддержание самозащиты
(self-tolerans)
Сохранение способности адекватного ИО против патогенов
Иммунная регуляция
Тreg
Нарушение образования (развития) и функции Тreg
Запуск аутоиммунной патологии
В 2001 г. описан наследственный синдром - IPEX (Immunodysregulation, Polyendocrinopathy, Enteropathy X-linked - сцепленная с Х-хромосомой иммунная дисрегуляция, полиэндокринопатия, энтеропатия), обусловленный мутацией гена Foxp3, приводящей к глубокому дефициту регуляторных Т-клеток (CD3+CD4+CD25+Foxp3+). Заболевание выявляют в перинатальном и младенческом возрастах. Оно сопровождается развитием множественных тяжёлых аутоиммунных и аллергических патологий и вскоре заканчивается летальным исходом.
. Срыв толерантности к ауто-АТ (аутотолерантность) и возникновение аутоиммунного")
Слайд 50МЕХАНИЗМЫ ЗАПУСКА АУТОИММУННОЙ ПАТОЛОГИИ.
Микробные суперантигены вызывают поликлональную активацию лимфоцитов. Какие-то из клонов
лимфоцитов, специфичные к собственным АГ, могут войти в режим эффекторного иммунного ответа (ИО).
Деструкция тканей патогеном (цитопатогенное действие вирусов, бактерий и др.) приводит к попаданию тканевых АГ в активированные (тем же патогеном) ДК, которые транспортируют все АГ в периферические лимфоидные органы для инициирования ИО. В этих условиях ДК могут утратить способность к толерантности к собственным АГ и вместо этого запускать ИО, реализуемый, как правило, с участием Тh1-лимфоцитов.
Два TCR на одном лимфоците. Примерно 30% периферических Т-л несут, по крайней мере, 2 разных по специфичности TCR .
Есть вероятность, что один из TCR может иметь специфичность к патогену, а второй - к ауто-АГ. Активация иммуногенеза патогеном приведёт к созданию клона лимфоцитов, которые будут работать в качестве эффекторов против обоих АГ - чужеродного и своего.
Деструкция тканей патогеном (цитопатогенное действие вирусов, бактерий и др.) приводит к попаданию тканевых АГ в активированные (тем же патогеном) ДК, которые транспортируют все АГ в периферические лимфоидные органы для инициирования ИО. В этих условиях ДК могут утратить способность к толерантности к собственным АГ и вместо этого запускать ИО, реализуемый, как правило, с участием Тh1-лимфоцитов.
Два TCR на одном лимфоците. Примерно 30% периферических Т-л несут, по крайней мере, 2 разных по специфичности TCR .
Есть вероятность, что один из TCR может иметь специфичность к патогену, а второй - к ауто-АГ. Активация иммуногенеза патогеном приведёт к созданию клона лимфоцитов, которые будут работать в качестве эффекторов против обоих АГ - чужеродного и своего.

Слайд 51СОМАТИЧЕСКИЕ КЛЕТКИ СТАНОВЯТСЯ АПК
В норме клетки организма (за исключением АПК) не
экспрессируют молекулы МНС II класса и не распознаются Тh.
Если клетки тех или иных органов начинают экспрессировать МНС II, они становятся мишенью для собственной иммунной системы.
примеры заболеваний с этим механизмом: инсулинзависимый сахарный диабет, тиреотоксикоз, аутоиммунный гепатит.
Причины необычной экспрессии молекул МНС II класса неизвестны. Усиление экспрессии этих молекул и их появление в необычных местах может вызвать ИФНγ.
ИФНγ -основной продукт Th1-клеток, возможно, этим объясняется способность полного адъюванта Фрейда индуцировать аутоиммунные процессы.
Во всех случаях такого рода заболеваний индуцируется аутоиммунный процесс клеточного типа.
Если клетки тех или иных органов начинают экспрессировать МНС II, они становятся мишенью для собственной иммунной системы.
примеры заболеваний с этим механизмом: инсулинзависимый сахарный диабет, тиреотоксикоз, аутоиммунный гепатит.
Причины необычной экспрессии молекул МНС II класса неизвестны. Усиление экспрессии этих молекул и их появление в необычных местах может вызвать ИФНγ.
ИФНγ -основной продукт Th1-клеток, возможно, этим объясняется способность полного адъюванта Фрейда индуцировать аутоиммунные процессы.
Во всех случаях такого рода заболеваний индуцируется аутоиммунный процесс клеточного типа.
 не экспрессируют молекулы МНС II")
Слайд 52Нарушение изоляции «иммунологически привилегированных» органов и тканей: ЦНС, внутренние среды глаза,
внутренние части семенников, фолликулы щитовидной железы и др.
В норме в эти органы не поступают иммунологически значимые сигналы.
При поступлении соответствующих АГ (основной белок миелина, тиреоглобулин, кристаллин и т.д.) может развиться аутоиммунный процесс.
Пример: аутоиммунное поражение парных органов при первоначальном повреждении (обычно при травме) одного из них:
«симпатическая офтальмия» (вовлечение в патологию здорового глаза при развитии воспалительного процесса в травмированном глазу),
поражения обоих яичек при аутоиммунном орхите, инициируемом травмой одного из них.
Для индукции аутоиммунного процесса требуется сочетание:
иммунизация «за барьерным» АГ,
воздействие на иммунную систему, вызывающее гиперактивацию Th1-клеток.
После преодоления барьеров гуморальное звено иммунной реакции срабатывает беспрепятственно: ауто-АТ, индуцированные в результате травмы одного глаза, «находят» второй глаз, не встречая преград.
В норме в эти органы не поступают иммунологически значимые сигналы.
При поступлении соответствующих АГ (основной белок миелина, тиреоглобулин, кристаллин и т.д.) может развиться аутоиммунный процесс.
Пример: аутоиммунное поражение парных органов при первоначальном повреждении (обычно при травме) одного из них:
«симпатическая офтальмия» (вовлечение в патологию здорового глаза при развитии воспалительного процесса в травмированном глазу),
поражения обоих яичек при аутоиммунном орхите, инициируемом травмой одного из них.
Для индукции аутоиммунного процесса требуется сочетание:
иммунизация «за барьерным» АГ,
воздействие на иммунную систему, вызывающее гиперактивацию Th1-клеток.
После преодоления барьеров гуморальное звено иммунной реакции срабатывает беспрепятственно: ауто-АТ, индуцированные в результате травмы одного глаза, «находят» второй глаз, не встречая преград.

Слайд 53АНТИГЕННАЯ МИМИКРИЯ.
У бактерий имеются АГ-детерминанты, перекрестно реагирующие с нормальными АГ.
В
норме ауто-реактивные клоны не активируются, т.к. на профессиональных АПК ауто-АГ присутствует в низких концентрациях, а на непрофессиональных АПК нет ко-стимулирующих молекул.
Появление перекрестно-реагирующего бактериального антигена (БАГ)– приведет аутореактивные клоны в активное состояние.
Другой механизм: в норме ауто-реактивные В-л не продуцируют АТ, так как лишены Тh помощи.
Но в качестве АПК, В-л захватывает перекрестно-реагирующий БАГ, презентирует фрагменты БАГ Тh .
В результате не аутореактивные Т-х начинают помогать аутореактивным В-л
Появление перекрестно-реагирующего бактериального антигена (БАГ)– приведет аутореактивные клоны в активное состояние.
Другой механизм: в норме ауто-реактивные В-л не продуцируют АТ, так как лишены Тh помощи.
Но в качестве АПК, В-л захватывает перекрестно-реагирующий БАГ, презентирует фрагменты БАГ Тh .
В результате не аутореактивные Т-х начинают помогать аутореактивным В-л


Слайд 55МОДИФИКАЦИЯ СТРУКТУРЫ СОБСТВЕННЫХ БЕЛКОВ ОРГАНИЗМА.
Присоединение гаптенов приводит к формированию эпитопов,
включающих, кроме гаптена, часть белковой молекулы. В случае перекрестного распознавания рецепторами Т- и В-клеток нормальных аутологичных эпитопов развивается аутоиммунная реакция.
α-метил-ДОФА индуцирует аутоиммунную гемолитическую анемию, при которой мишенью АТ становятся молекулы D (Rh) АГ.
Пенициллинамид и прокаинамид вызывают системную аутоагрессию вплоть до волчаночного синдрома.
Изониазид может вызвать образование антиядерных антител с клиническими проявлениями в виде полиартрита.
β-адреномиметики – астматический статус.
Однако строгие доказательства прямой связи индукции аутоиммунного процесса с модификацией ауто-АГ отсутствуют.
α-метил-ДОФА индуцирует аутоиммунную гемолитическую анемию, при которой мишенью АТ становятся молекулы D (Rh) АГ.
Пенициллинамид и прокаинамид вызывают системную аутоагрессию вплоть до волчаночного синдрома.
Изониазид может вызвать образование антиядерных антител с клиническими проявлениями в виде полиартрита.
β-адреномиметики – астматический статус.
Однако строгие доказательства прямой связи индукции аутоиммунного процесса с модификацией ауто-АГ отсутствуют.

Слайд 56Иммунологические механизмы аутоиммунных поражений
гуморального типа накопление ауто-АТ - IgG их участие
в :
АТ зависимой цитотоксичности –
ГНТ II типа (гемолитическая анемия и др. поражения клеток крови)
иммунокомплексной –
ГНТ III типа (системная красная волчанка)
стимулирующей (ауто-АТ к рецепторам TSH при тиреотоксикозе)
путем подключения комплемента (комплемент зависимый цитолиз), МФ (опсонизация), NK (АТ зависимый клеточно-опосредованный цитолиз),
запуска активирующих сигналов через рецептор—мишень ауто-АТ.
АТ зависимой цитотоксичности –
ГНТ II типа (гемолитическая анемия и др. поражения клеток крови)
иммунокомплексной –
ГНТ III типа (системная красная волчанка)
стимулирующей (ауто-АТ к рецепторам TSH при тиреотоксикозе)
путем подключения комплемента (комплемент зависимый цитолиз), МФ (опсонизация), NK (АТ зависимый клеточно-опосредованный цитолиз),
запуска активирующих сигналов через рецептор—мишень ауто-АТ.
клеточного типа- более тяжелые и менее чувствительные к лечебным воздействиям.
варианты клеточных механизмов
цитотоксический – цитолиз, опосредованный CD8+ (инсулинзависимый сахарный диабет)
ГЗТ – разрушение макрофагами (активированными Th1) и их продуктами с последующим формированием очага хронического иммунного воспаления (рассеянный склероз и ревматоидный артрит).
При цитотоксическом механизме поражения более локализованные, менее деструктивные, последствия связаны с уникальностью поражаемых клеток (сахарный диабет)
При развитии ГЗТ в патологию вовлекаются значительные массивы тканей, повреждения более выраженные.

Слайд 57Классификация аутоиммунных заболеваний
В настоящее время нет единой классификации аутоиммунных заболеваний

Слайд 58Системные аутоиммунные заболевания - когда поражаются различные органы либо ткани , причем
поражаются органы в разной степени и тогда лечение проходит у того врача симптомы болезни которой наиболее ярко выражены или у нескольких специалистов.
Системный склероз (склеродермия)
Системная красная волчанка
Антифосфолипидный синдром
Полимиозит и дермаполимиозит
Синдром Шегрена
Ревматоидный артрит
Болезнь Бехчета
Системные васкулиты - это группа разных отдельных болезней, объединенных на основании воспаления сосудов
Системный склероз (склеродермия)
Системная красная волчанка
Антифосфолипидный синдром
Полимиозит и дермаполимиозит
Синдром Шегрена
Ревматоидный артрит
Болезнь Бехчета
Системные васкулиты - это группа разных отдельных болезней, объединенных на основании воспаления сосудов

Слайд 59Диффузный токсический зоб характеризуется триадой — гипертиреоидизм, зоб экзофтальм (выпученные глаза).
Аутоиммунный тиреоидит
- Хашимото
Аутоиммунные заболевания органов эндокринной системы
Аутоиммунный тиреоидит - Хашимото
Сахарный диабет 1-го типа
Болезнь Грейвса
. Аутоиммунный тиреоидит - Хашимото Аутоиммунные заболевания")

Слайд 61Аутоиммунные заболевания крови
Аутоиммунная гемолитическая анемия
Аутоиммунная нейтропения
Аутоиммунная тромбоцитопения
Тромбоцитопеническая пурпура
Аутоиммунная тромбоцитопения

Слайд 62Аутоиммунные заболевания кожи
Витилиго
Пемфигоид
Изолированные
васкулиты кожи
Псориаз
Хроническая крапивница
Некоторые формы
алопеции
Дискоидная
красная волчанка

Слайд 63Аутоиммунные заболевания нервной системы
Множественный или рассеянный склероз
Синдром Гиена-Баре-самая частая острая полиневропатия
Миастения
Гравис
Миастения Гравис
Множественный или рассеянный склероз

Слайд 64Классификация аутоиммунных заболеваний (по механизму иммунопатологического процесса) - в основе патогенеза
тип реакции гиперчувствительности по Джеллу и Кумбсу
 - в основе патогенеза тип реакции гиперчувствительности по")
Слайд 65Классификация аутоиммунных заболеваний (по механизму иммунопатологического процесса) - продолжение
 - продолжение")
Слайд 66Классификация аутоиммунных заболеваний (по механизму иммунопатологического процесса) - продолжение
 - продолжение")

Слайд 68Противоопухолевый
иммунитет
Часть 3
Онкопатология занимает 2 место после сердечно-сосудистых заболеваний среди причин
смерти в цивилизованных странах.
Опухоли (новообразования) возникают, когда по каким-либо причинам наблюдается неконтролируемая избыточная пролиферация клеток.
Обычно во всех органах и тканях поддерживается баланс между процессами гибели клеток и их новообразованием. Это обеспечивается механизмами контроля и регуляции гомеостаза тканей.
Опухоли (новообразования) возникают, когда по каким-либо причинам наблюдается неконтролируемая избыточная пролиферация клеток.
Обычно во всех органах и тканях поддерживается баланс между процессами гибели клеток и их новообразованием. Это обеспечивается механизмами контроля и регуляции гомеостаза тканей.


Слайд 70ОПУХОЛИ
(незрелые, гетерологичные) состоят из умеренно- и малодифференцированных клеток.
могут утратить сходство
с тканью, из которой они исходят.
Характерен:
быстрый, чаще инфильтрирующий рост,
метастазирование и рецидивирование,
наличие общего влияния на организм.
характерен как клеточный, так и тканевой атипизм.
Характерен:
быстрый, чаще инфильтрирующий рост,
метастазирование и рецидивирование,
наличие общего влияния на организм.
характерен как клеточный, так и тканевой атипизм.
(зрелые, гомологичные) состоят из клеток, дифференцированных в такой мере, что можно определить, из какой ткани они растут.
Характерен:
медленный экспансивный рост,
отсутствие метастазов,
отсутствие общего влияния на организм.
могут малигнизироваться.
Доброкачественные
Злокачественные
 состоят из умеренно- и малодифференцированных клеток. могут утратить сходство с тканью, из которой")
Слайд 71Опухоли способны
возникать спонтанно, без видимых на то причин
могут быть индуцированы.
Индуцировать канцерогенез
способны
различные химические вещества (метилхолантрен, бензпирен, афлатоксин В1),
физические факторы (ультрафиолетовый свет, ионизирующая радиация)
вирусы ( герпес-, адено- и ретровирусы).
У человека 3 типа вирусов тесно связаны с опухолями:
вирус Эпштейна-Барр (лимфома Беркита),
цитомегаловирус (саркома Капоши)
вирус папилломы.
различные химические вещества (метилхолантрен, бензпирен, афлатоксин В1),
физические факторы (ультрафиолетовый свет, ионизирующая радиация)
вирусы ( герпес-, адено- и ретровирусы).
У человека 3 типа вирусов тесно связаны с опухолями:
вирус Эпштейна-Барр (лимфома Беркита),
цитомегаловирус (саркома Капоши)
вирус папилломы.

Слайд 72Противоопухолевые иммунные факторы
Клеточные:
1) NK-клетки
2) Т-лимфоциты-киллеры;
3) активированные МФ;НГ
II. Гуморальные:
1) специфические АТ к АГ опухоли приемущественно IgM и IgG;
АТ не разрушают опухоли, адсорбируясь усиливают цитолиз
2) ИЛ-1 и ИЛ-2;
3) фактор некроза опухолей (ФНО);
4) интерфероны.
 NK-клетки 2) Т-лимфоциты-киллеры; 3) активированные МФ;НГ II.")
Слайд 73ИММУННЫЙ НАДЗОР ОПУХОЛЕВОГО РОСТА
Концепция была предложена Ф.М. Бернетом в 70-е годы.
У человека за сутки возникает > 1000 000 мутантных клеток, значительная часть которых подвергается опухолевой трансформации.
Причина низкой частоты реализации этой опасности - наличие иммунного надзора.
ИС осуществляет контроль элиминируя любые клетки, несущие чужеродную генетическую информацию.
немногие клетки, маскирующие проявления своей чужеродности обходят этот контроль и дают начало опухолям.
приведенные факты объясняются вирусной этиологией опухолей.
У здоровых людей многие вирусы циркулируют всю жизнь, но репликация их подавлена ИО.
Доказана этиологическая роль:
вируса гепатита В - рак печени,
вируса папилломы человека - рак шейки матки,
вируса Эпштейна-Барр -лимфомы и рак носоглотки,
вируса HTLV-1( T-лимфотропный вирус человека типа 1) -Т-клеточный лейкоз.
На большинство опухолей ИО или вообще не развивается или он поздний и неэффективный.
Причина -отсутствие иммуногенности злокачественных опухолей, иначе отсутствие опухолеспецифических АГ

Слайд 74Антигены, ассоциированные с опухолями
Опухолеспецифические АГ (трансплантационные АГ)
присутствуют только на опухолевых клетках
Имеют
вирусное происхождение
Опухолеассоциированные АГ – гетероорганные АГ
нельзя отнести к чужеродным
типичные АГ для нормальной ткани
АГ с упрощенным строением
Исследование АГ необходимо для расшифровки механизма ИО и создания новых способов диагностики и лечения.
Доказательств существования АГ, специфичных только для опухолей мало.
Далеко не все опухоли содержат маркерные АГ
Все эти АГ распознаются с МНС I класса, чаще всего HLA-A2 и индуцируют дифференцировку цитотоксических Т-клеток.
присутствуют только на опухолевых клеткахИмеют вирусное происхождениеОпухолеассоциированные АГ")
Слайд 75АНТИГЕНЫ, АССОЦИИРОВАННЫЕ С ОПУХОЛЯМИ
раково-эмбриональный антиген (РЭА)- ассоциирующий с раком толстой кишки,
хорионический
гонадотропин (ХГЧ) (гормон, в норме продуцируемый в плаценте)- выделяемый хориокарциномой, семиномой и рядом опухолей, происходящих из эмбриональных тканей.
Большинство этих АГ – результат экспрессии генов, которые в норме «работают» в эмбриональном периоде.
мембранные АГ опухолевых клеток – опухолеспецифические трансплантационные АГ (Tumour-specific transplantation antigen, TSTA).
К опухолеассоциированным АГ также относят муцины. Описано 7 представителей этой группы (MUC1, MUC2 и т.д.). Их гены образуют кластер в хромосоме 11.
В нормальных эпителиальных клетках они участвуют в распознавании молекул адгезии. Муцины особенно характерны для карцином кишечника. Полагают, что муцины опухолевых клеток имеют значение в процессе метастазирования.
Большинство этих АГ – результат экспрессии генов, которые в норме «работают» в эмбриональном периоде.
мембранные АГ опухолевых клеток – опухолеспецифические трансплантационные АГ (Tumour-specific transplantation antigen, TSTA).
К опухолеассоциированным АГ также относят муцины. Описано 7 представителей этой группы (MUC1, MUC2 и т.д.). Их гены образуют кластер в хромосоме 11.
В нормальных эпителиальных клетках они участвуют в распознавании молекул адгезии. Муцины особенно характерны для карцином кишечника. Полагают, что муцины опухолевых клеток имеют значение в процессе метастазирования.
- ассоциирующий с раком толстой кишки,хорионический гонадотропин (ХГЧ) (гормон,")
Слайд 76
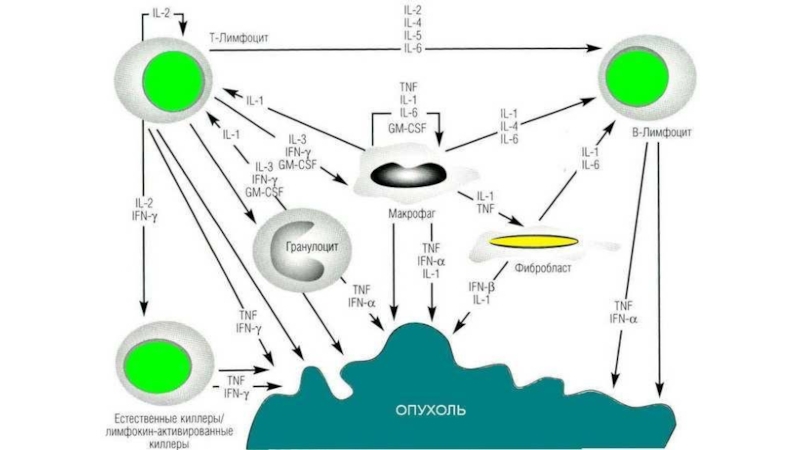
Противоопухолевый иммунитет основан на Тh1-зависимом клеточном иммунном ответе, активирующем цитотоксические Т-л,
макрофаги, дендритные клетки, NK-, T-NK-клетки. Роль гуморального (антительного) невелика.

Слайд 78МЕХАНИЗМЫ ИЗБЕГАНИЯ ОПУХОЛЯМИ ИММУННОГО НАДЗОРА
1. Слабая экспрессия молекул МНС I класса.
опухоли
слабо экспрессируют МНС I, необходимые для распознавания АГ CD8+-Т-л.
Но полная утрата мембранных МНС-I делает опухолевые клетки чувствительными к NK-клеток.
Оптимальным для опухолевых клеток является промежуточное «решение»: утрата молекул МНС I класса некоторых типов (например, HLA-A) при сохранности других типов этих молекул. (при раке толстой кишки).
2. Нарушение адгезии и корецепции.
опухолевые клетки лишены молекул адгезии( LFA-1, LFA-3, ICAM-1),и могут экспрессировать антиадгезивные молекулы (муцин).
Нарушение адгезии снижает эффективность фагоцитоза и килинга.
Эффективный механизм защиты опухолей – отсутствие экспрессии молекул CD80 и CD86, распознаваемых ко-рецептором CD28.
Без сигнала, поступающего с ко-рецептора, развивается анергия Т-клетки.
Возможно, именно этот механизм является главным способом самозащиты опухолевых клеток.
разрушаются клетки опухоли, лишенные молекулы CD80 (адаптивный иммунитет за счет цитотоксических CD8+-клеток, не нуждающихся в костимуляции).
Но полная утрата мембранных МНС-I делает опухолевые клетки чувствительными к NK-клеток.
Оптимальным для опухолевых клеток является промежуточное «решение»: утрата молекул МНС I класса некоторых типов (например, HLA-A) при сохранности других типов этих молекул. (при раке толстой кишки).
2. Нарушение адгезии и корецепции.
опухолевые клетки лишены молекул адгезии( LFA-1, LFA-3, ICAM-1),и могут экспрессировать антиадгезивные молекулы (муцин).
Нарушение адгезии снижает эффективность фагоцитоза и килинга.
Эффективный механизм защиты опухолей – отсутствие экспрессии молекул CD80 и CD86, распознаваемых ко-рецептором CD28.
Без сигнала, поступающего с ко-рецептора, развивается анергия Т-клетки.
Возможно, именно этот механизм является главным способом самозащиты опухолевых клеток.
разрушаются клетки опухоли, лишенные молекулы CD80 (адаптивный иммунитет за счет цитотоксических CD8+-клеток, не нуждающихся в костимуляции).

Слайд 79МЕХАНИЗМЫ ИЗБЕГАНИЯ ОПУХОЛЯМИ ИММУННОГО НАДЗОРА
3.Гены опухолевых АГ мутируют.
в ходе естественной селекции
отбираются варианты опухолевых клеток, несущие мутантные АГ с которыми АТ и ЦТЛ( CD8+-лимфоциты) не реагируют- резко снижают эффективность иммунной защиты
4.Формирование иммунологической толерантности к опухолевому антигену.
может формироваться под действием низкодозных растворимых форм опухолевого АГ. Индуцирует развитие толерантности Т-л, отсутствие на опухолевых клетках ко-стимулирующих молекул.
5. Опухолевые клетки подавляют иммунный ответ.
выделяют цитокины, которые подавляют иммунный ответ (TFRα и β, ИЛ-10, простагландин Е2).
Способствуют выживанию опухолевых клеток ( фактор, активирующий ростовую активность меланоцитов, а также факторы роста фибробластов и тромбоцитов).
4.Формирование иммунологической толерантности к опухолевому антигену.
может формироваться под действием низкодозных растворимых форм опухолевого АГ. Индуцирует развитие толерантности Т-л, отсутствие на опухолевых клетках ко-стимулирующих молекул.
5. Опухолевые клетки подавляют иммунный ответ.
выделяют цитокины, которые подавляют иммунный ответ (TFRα и β, ИЛ-10, простагландин Е2).
Способствуют выживанию опухолевых клеток ( фактор, активирующий ростовую активность меланоцитов, а также факторы роста фибробластов и тромбоцитов).

Слайд 80МЕХАНИЗМЫ ИЗБЕГАНИЯ ОПУХОЛЯМИ ИММУННОГО НАДЗОРА
6.Противоопухолевые антитела защищают опухоль.
АТ, индуцируемые к опухолевым
АГ, защищают опухоли от действия Т-л.
( феномен был описан в 50-е годы и назван феноменом усиления (enhancing) опухолевого роста)
Данный факт свидетельствует о ведущей роли клеточных механизмов противоопухолевой защиты.
Переключение на гуморальный тип ИО свидетельствует о низкой эффективности клеточного типа.
Защитный эффект обусловлен тем, что АТ неспособны вызвать комплементзависимый цитолиз, так как на опухолевых клетках (как и соматических) экспрессируются CR1 и DAF.
АТ ( изотипов IgG1 и IgG3) не опсонизируют клетки-мишени, и поэтому, не способствуют их фагоцитозу или киллингу NK-клетками.
Отсутствие опсонизации можно объяснить двумя причинами:
АТ связываются с мембранным АГ и образующийся комплекс не погружается внутрь клетки, ресинтез мембранного АГ не происходит
опухолевые клетки могут синтезировать секреторный вариант АГ , либо растворимые формы мембранных АГ смываются с клеток.
Растворимый антиген связывает противоопухолевые антитела, не позволяя им взаимодействовать с опухолевой клеткой.
7 Растворимый антиген блокирует цитотоксические клетки.
( феномен был описан в 50-е годы и назван феноменом усиления (enhancing) опухолевого роста)
Данный факт свидетельствует о ведущей роли клеточных механизмов противоопухолевой защиты.
Переключение на гуморальный тип ИО свидетельствует о низкой эффективности клеточного типа.
Защитный эффект обусловлен тем, что АТ неспособны вызвать комплементзависимый цитолиз, так как на опухолевых клетках (как и соматических) экспрессируются CR1 и DAF.
АТ ( изотипов IgG1 и IgG3) не опсонизируют клетки-мишени, и поэтому, не способствуют их фагоцитозу или киллингу NK-клетками.
Отсутствие опсонизации можно объяснить двумя причинами:
АТ связываются с мембранным АГ и образующийся комплекс не погружается внутрь клетки, ресинтез мембранного АГ не происходит
опухолевые клетки могут синтезировать секреторный вариант АГ , либо растворимые формы мембранных АГ смываются с клеток.
Растворимый антиген связывает противоопухолевые антитела, не позволяя им взаимодействовать с опухолевой клеткой.
7 Растворимый антиген блокирует цитотоксические клетки.

Слайд 81Пробластомные факторы
подавляют иммунитет
1. Супресcивные вещества, продуцируемые ЛФ и МФ
2. Блокирующие
АТ
3. Циркулирующие иммунные комплексы
4. Простагландины ПГЕ2
5. ИЛ-10
6. Трансформирующий фактор роста (TGFβ), подавляющий:
а) продукцию цитокинов (ИЛ-12);
б) созревание Т-киллеров;
в) экспрессию рецепторов к цитокинам.
на определенном этапе развития опухоли иммунная система хозяина начинает выделять пробластомные (способствующие росту опухоли) факторы, которые:
усиливают рост опухоли:
1. Фактор роста опухоли, продуцируемый МФ
2. ИЛ-2 и ИЛ- 6.
3.ИФН y.
4. Фактор роста сосудистого эндотелия.
5. Иммунодефицитное состояние:
а) нарушение созревания Т-к;
б) нарушение функции АПК



Слайд 85Иммунодиагностика рака-МЕТОД ИФА
основана на выявлении в сыворотке и жидкостях больного
опухолеспецифических АГ или маркеров опухолевого роста (опухолеассоциированных белков).
тест-системы:
– СЕА; используется для выявления РЭА, положительный тест при карциноме толстой кишки и поджелудочной железы.
– AFP; для выявления альфа-фетопротеина, положительный тест при первичном раке печени.
– СА-125 маркер рака яичника.
– СА-15-3 маркер рака молочной железы.
– СА-19-9 маркер рака ЖКТ.
– СА-242 маркер рака поджелудочной железы, толстой и прямой кишки.
– PSA EIA специфический антиген простаты.
– PSA Free; специфический АГ простаты, определение свободного PSA.
– NSE; маркер мелкоклеточного рака легких.
– UBC; маркер рака мочевого пузыря.
– CASA маркер рака яичника.
– MSA; маркер рака молочной железы.
– Chromogranin A маркеры опухолей эндокринного происхождения.
– CD30 (Ki-1 Antigen); опухолилимфопролиферативного происхождения
тест-системы:
– СЕА; используется для выявления РЭА, положительный тест при карциноме толстой кишки и поджелудочной железы.
– AFP; для выявления альфа-фетопротеина, положительный тест при первичном раке печени.
– СА-125 маркер рака яичника.
– СА-15-3 маркер рака молочной железы.
– СА-19-9 маркер рака ЖКТ.
– СА-242 маркер рака поджелудочной железы, толстой и прямой кишки.
– PSA EIA специфический антиген простаты.
– PSA Free; специфический АГ простаты, определение свободного PSA.
– NSE; маркер мелкоклеточного рака легких.
– UBC; маркер рака мочевого пузыря.
– CASA маркер рака яичника.
– MSA; маркер рака молочной железы.
– Chromogranin A маркеры опухолей эндокринного происхождения.
– CD30 (Ki-1 Antigen); опухолилимфопролиферативного происхождения
.

Слайд 86Иммунотерапия больных с онкопатологией
ведется по двум направлениям
1) коррекция иммунных расстройств,
развивающихся в процессе проведения химио- и радиотерапии;
2) активация противоопухолевых иммунных механизмов.
Для лечения гемо- и иммунодепрессий, развивающихся в результате токсического действия опухолей на гемопоэз и иммуногенез, а также как следствия применения высоких доз радио- и химиотерапии, в соответствии с показаниями, могут быть использованы:
трансфузия совместимого костного мозга
трансфузия лейкомассы и тромбоцитарной взвеси (в том числе и аутологичной);
иммунотропные препараты, влияющие на Т-В-л и МФ.
Иммунотропные препараты оказывают лечебное и иммунокоррегирующее действие только в случае наличия клеточной базы компенсации дефектов гемо- и лимфопоэза.
2) активация противоопухолевых иммунных механизмов.
Для лечения гемо- и иммунодепрессий, развивающихся в результате токсического действия опухолей на гемопоэз и иммуногенез, а также как следствия применения высоких доз радио- и химиотерапии, в соответствии с показаниями, могут быть использованы:
трансфузия совместимого костного мозга
трансфузия лейкомассы и тромбоцитарной взвеси (в том числе и аутологичной);
иммунотропные препараты, влияющие на Т-В-л и МФ.
Иммунотропные препараты оказывают лечебное и иммунокоррегирующее действие только в случае наличия клеточной базы компенсации дефектов гемо- и лимфопоэза.
 коррекция иммунных расстройств, развивающихся в процессе проведения")

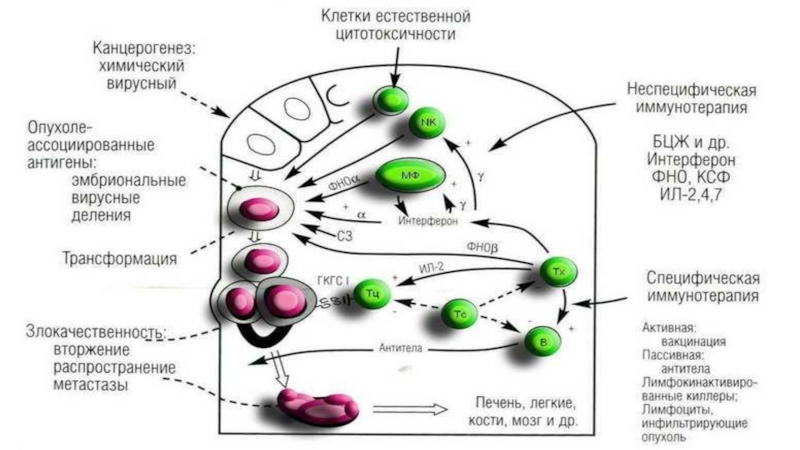
Слайд 89
ДЕЙСТВИЕ ОНКОВАКЦИНЫ НА ОСНОВЕ DC
заключается в стимуляции ЦТЛ (СD8+лф) пациента против определённой опухоли:
активация дендритных клеток (ДК) in vitro опухолевыми АГ,
активированные ДК, презентирующие опухолевый АГ на своей поверхности взаимодействуют с наивными Т-лимфоцитами,
последние дифференцируются в "профессиональные цитотоксические лимфоциты-киллеры (ЦТЛ)", мигрируют и лизируют определённую опухоль в организме.
заключается в стимуляции ЦТЛ (СD8+лф) пациента против определённой опухоли:
активация дендритных клеток (ДК) in vitro опухолевыми АГ,
активированные ДК, презентирующие опухолевый АГ на своей поверхности взаимодействуют с наивными Т-лимфоцитами,
последние дифференцируются в "профессиональные цитотоксические лимфоциты-киллеры (ЦТЛ)", мигрируют и лизируют определённую опухоль в организме.

Слайд 90Схема действия «мгновенного иммунитета». Выработавшиеся в ответ на вакцину универсальные антитела
сначала ковалентно связываются с отдельно вводимым в кровь адаптером, подбираемым в зависимости от мишени. Потом эта система связывается с мишенью на поверхности опухолевой клетки, в данном случае – двумя типами интегринов. И уже после этого опухолевую клетку, облепленную антителами «замечают» естественные киллеры и система комплемента (CDC). //Carlos F. Barbas /PNAS






