- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Высшие жирные кислоты. Липиды презентация
Содержание
- 1. Высшие жирные кислоты. Липиды
- 2. Липиды - (от греч. lípos - жир)
- 3. Классификация липидов Липиды подразделяют на простые и
- 4. Нейтральные липиды - это сложные эфиры жирных
- 5. В структуру нейтральных липидов входит глицерол, остатки
- 7. Высшие жирные кислоты в составе липидов Жирные
- 8. Высшие жирные кислоты в составе природных липидов
- 9. Номенклатура жирных кислот основана на определении длины
- 10. Примеры названий нейтральных липидов (триацилглицеролов).
- 11. Модель структуры жирных кислот
- 12. Структура триацилглицеролов (триглицеридов)
- 13. Эйкозаноиды -обширная группа физиологически и фармакологически активных
- 14. Схема биосинтеза эйкозаноидов
- 15. Простагландины (ПГ, Pg). По существу ПГ представляют
- 16. ПГ проявляют свое действие в чрезвычайно низких
- 17. Простациклины образуются в стенках кровеносных сосудов и
- 18. Лейкотриены. Это производные 20-углеродных полиненасыщенных (эй-козановых) кислот.
- 19. Происхождение высших ЖК и их биологическое действие
- 20. Структура сложных липидов Глицерофосфолипиды
- 21. Глицерофосфолипиды – производные глицерол-3-фосфата. Природные глицерофосфолипиды имеют
- 22. Среди глицерофосфолипидов наиболее распространены L-фосфатиды:
- 23. Как правило, в природных фосфатидах в положении
- 24. Структура кардиолипина –фосфолипида, выделяемого из сердечной мышцы
- 25. Несколько менее распространены липиды с простой эфирной связью – плазмалогены:
- 26. Сфинголипиды представляют собой структурные аналоги глицерофосфолипидов, где
- 27. Важную группу сфинголипидов составляют сфингомиелины, впервые обнаруженные
- 28. Гликолипиды включают углеводные остатки, чаще всего D-галактозу,
- 29. Ганглиозиды – богатые углеводами сложные липиды, впервые
- 30. Характерной особенностью сложных липидов является их бифильность,
- 32. На поверхности раздела двух водных фаз полярные
- 33. Фрагмент структуры биологических мембран
- 34. Плазматическая мембрана
- 35. Липидные компоненты плазматической мембраны. Во внешнем
- 36. Мембраны двух соседних нервных клеток (электронный микроскоп,
- 37. Неомыляемые (сопутствующие) вещества 1. Жирорастворимые витамины (А,
- 38. Стероиды
- 39. Биологическая роль липидов Структурная функция. Молекулы фосфолипидов
- 40. Транспортная функция. Липиды являются переносчиками жирорастворимых витаминов
- 41. Функциональная классификация липидов: резервные; структурные; транспортные.
- 42. Липопротеины представляют собой сферические частицы (мицеллы),
- 43. Различают 4 класса плазменных липопротеинов:
- 44. Липопротеины низкой плотности
- 45. Фосфолипиды наружной оболочки липопротеинов связаны со специ-фическими
- 46. Аполипопротеины выполняют три основные функции:
- 47. Химические свойства омыляемых липидов Реакция гидролиза –
- 49. Реакция присоединения – Липиды содержащие остатки непредельных
- 50. Значения йодного числа для ряда природных масел, жиров, индивидуальных жирных кислот
- 51. В промышленности широко применяется каталитическое гидрогенирование ненасыщенных
- 52. Реакции окисления Процессы окисления липидов весьма
- 53. Указанная реакция является начальной стадией процесса так
- 55. Липиды – важный энергетический субстрат. Высвобождение энергии,
- 57. При атаке таким радикалом по метиленовой группе
- 58. Неомыляемые липиды Особую группу терпенов составляют каротиноиды
- 59. К терпенам относятся и другие жирорастворимые витамины,

Слайд 2Липиды - (от греч. lípos - жир) -это неоднородная группа химических
соединений,
 -это неоднородная группа химическихсоединений, общим свойством которых является")
Слайд 3Классификация липидов
Липиды подразделяют на простые и сложные, которые в свою очередь
К группе омыляемых липидов относят нейтральные липиды и полярные липиды (фосфолипиды).
Неомыляемые липиды включают группу жирорастворимых витаминов (А,Е, D, К, Q10), терпены, стерины и жирорастворимые гормоны. При этом терпены обнаруживают в липидах растений, а стерины и гормоны – в липидах животных организмов.

Слайд 4Нейтральные липиды - это сложные эфиры жирных кислот с различными спиртами.
Полярные липиды (фосфолипиды) представляют собой сложные эфиры жирных кислот со спиртами, содержащие, дополнительно, и другие группы.
Омыляемые сложные липиды делят на несколько групп: – фосфолипиды, сфинго-липиды и гликолипиды, сульфолипиды, аминолипиды, липопротеины.
Наиболее распространены фосфоипиды, которые характеризуются значительным содержанием ненасыщенных жирных кислот.

Слайд 5В структуру нейтральных липидов входит глицерол, остатки высших жирных кислот, в

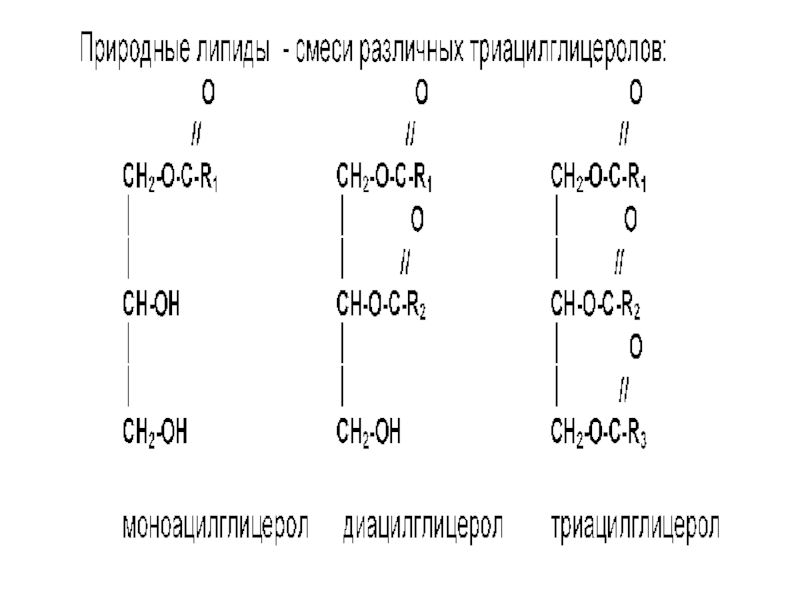
Слайд 7Высшие жирные кислоты в составе липидов
Жирные кислоты являются алифатическими длинноцепочечными карбоновыми

Слайд 8Высшие жирные кислоты в составе природных липидов
*-ω-число атомов углерода от двойной
**-номера углеродных атомов, после которых расположены π-связи

Слайд 9Номенклатура жирных кислот основана на определении длины цепи и положения двойных
Жирная кислота обозначается индексом, в котором длина цепи обозначается цифрой, через двоеточие обозначается количество двойных связей. Знаком Δ с соответствующей цифрой показывается удаление двойной связи от карбоксильной группы (т.е. от первого атома углерода).
Среди ненасыщенных жирных кислот выделяют семейства омега-9 (ω 9 или n), омега-6 (ω 6) и омега-3 (ω 3) в соответствии с положением двойных связей относительно концевой метильной группы углеводородной цепи (т.е последнего углеродного атома, наиболее удаленного от карбоксильной группы) (дистального конца). Например, насыщенная пальмитиновая кислота обозначается индексом (16:0), мононенасыщенная олеиновая кислота – (ω 9, 18:1, Δ9), индекс арахидоновой кислоты (ω 6 20:4, Δ5,8,11,14).

.")

")
Слайд 13Эйкозаноиды -обширная группа физиологически и фармакологически активных соединений. К ним относятся
")

Слайд 15Простагландины (ПГ, Pg). По существу ПГ представляют собой 20-углеродные жирные кислоты,
. По существу ПГ представляют собой 20-углеродные жирные кислоты, содержащие 5-углеродное кольцо и")
Слайд 16ПГ проявляют свое действие в чрезвычайно низких концентрацияхПГ проявляют свое действие
В целом ПГ, не являясь гормонамиВ целом ПГ, не являясь гормонами, модулируют действие последних. Они преимущественно влияют на физиологические функции тех клетокВ целом ПГ, не являясь гормонами, модулируют действие последних. Они преимущественно влияют на физиологические функции тех клеток, в которых синтезируются. Характер воздействия ПГ зависит от типа клеткиВ целом ПГ, не являясь гормонами, модулируют действие последних. Они преимущественно влияют на физиологические функции тех клеток, в которых синтезируются. Характер воздействия ПГ зависит от типа клетки, и этим ПГ отличаются от гормонов с их однозначным эффектом.
ПГ могут использоваться как терапевтическое средство для предотвращения оплодотворения, стимулирования нормальных родов, прерывания беременностиПГ могут использоваться как терапевтическое средство для предотвращения оплодотворения, стимулирования нормальных родов, прерывания беременности, предупреждения развития или обезболивания язвы желудка, лечения воспалительных процессов и регуляции кровяного давления, а также для снятия приступов астмы и др.

Слайд 17Простациклины образуются в стенках кровеносных сосудов и являются сильными ингибиторамиобразуются в

Слайд 18Лейкотриены. Это производные 20-углеродных полиненасыщенных (эй-козановых) кислот. Название «лейкотриены» происходит от
Лейкотриены рассматриваются прежде всего как медиаторы воспалительных реакций; они вызывают сокращение мышечной тканиЛейкотриены рассматриваются прежде всего как медиаторы воспалительных реакций; они вызывают сокращение мышечной ткани бронхов в концентрацияхЛейкотриены рассматриваются прежде всего как медиаторы воспалительных реакций; они вызывают сокращение мышечной ткани бронхов в концентрациях, в 100–1000 раз меньших, чем гистамин; способствуют сокращению коронарных сосудов.
 кислот. Название «лейкотриены» происходит от двух слов: «лейкоциты» (впервые")
Слайд 19Происхождение высших ЖК и их биологическое действие
Растительные масла и липиды наземных
Наиболее ненасыщенные пента- и гексаеновые жирные кислоты 20:5 и 22:6, относящиеся к ряду ω-3 кислот, находятся в липидах гидробионтов, в первую очередь в морской рыбе.
Существуют конкурентные взаимоотношения между метаболитами, образуемыми из ω-6 и ω-3 кислот.
Простагландины из ω-3 кислот снижают давление, а из ω-6 – повышают;
Тромбоксаны из ω-3 кислот снижают свертываемость, а из ω-6 – повышают;
Лейкотриены из ω-3 кислот усиливают иммунитет, а из ω-6 – понижают его, способствуют развитию воспалительных реакций
Таким образом, в структуре питания должно быть оптимальное соотношение кислот ω-3 и ω-6 рядов (1:3).
Показано, что липиды, содержащие ω-3 кислоты, проявляют гипохолестеринемическое, антиаритмическое, иммуномоделирующее действие. Употребление липидов или рыбы не менее 2-3 раз в неделю вдвое снижает риск внезапной смерти от сердечно-сосудистых катастроф, увеличивает выживаемость при онкологических заболеваниях.


Слайд 21Глицерофосфолипиды – производные глицерол-3-фосфата. Природные глицерофосфолипиды имеют одинаковую конфигурацию и являются
.


Слайд 23Как правило, в природных фосфатидах в положении С1 находится остаток насыщенной



Слайд 26Сфинголипиды представляют собой структурные аналоги глицерофосфолипидов, где вместо глицерина используется сфингозин

Слайд 27Важную группу сфинголипидов составляют сфингомиелины, впервые обнаруженные в нервной ткани. В

Слайд 28Гликолипиды включают углеводные остатки, чаще всего D-галактозу, и не содержат остатков

Слайд 29Ганглиозиды – богатые углеводами сложные липиды, впервые выделенные из серого вещества

Слайд 30Характерной особенностью сложных липидов является их бифильность, обусловленная присутствием в составе

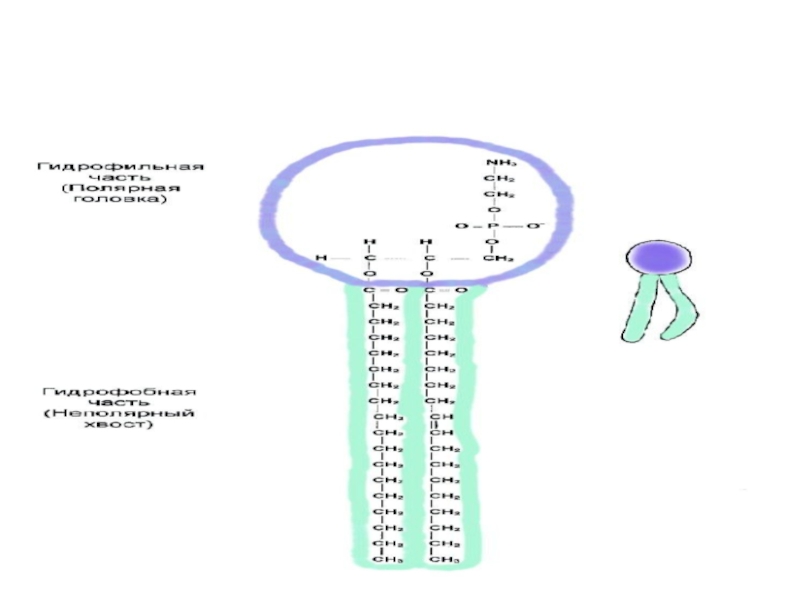
Слайд 32На поверхности раздела двух водных фаз полярные фосфолипиды самопроизвольно формируют бислои.



Слайд 35Липидные компоненты плазматической мембраны. Во внешнем слое липидного бислоя преобладают фосфатидилхолин,

Слайд 36Мембраны двух соседних нервных клеток (электронный микроскоп, увелич. в 400 000
. Каждая мембрана имеет")
Слайд 37Неомыляемые (сопутствующие) вещества
1. Жирорастворимые витамины (А, Е, D, К, убихинон Q10
 вещества1. Жирорастворимые витамины (А, Е, D, К, убихинон Q10 ) локализваны в биологической")

Слайд 39Биологическая роль липидов
Структурная функция. Молекулы фосфолипидов обладают поверхностно-активными свойствами. В водных
Трансформационная функция. Линоленовая, арахидоновая и эйкозапентаеновая кислоты в организме человека трансформируются в эйкозаноиды -высокоактивные биогенные соединения, являющимися модуляторами функционирования практически всех систем организма. Ввиду исключительной биологической ценности ПНЖК они являются эссенциальными (незаменимыми) (витамин F).
Энергетическая и резервная функция. Калорийность липидов примерно в в 2 раза выше калорийности белков и углеводов, поэтому они являются существенными источниками энергии и скапливаются в “жировом депо” человека в качестве запасного субстрата для синтеза АТФ.

Слайд 40Транспортная функция. Липиды являются переносчиками жирорастворимых витаминов группы А, D, Е,

Слайд 41Функциональная классификация липидов:
резервные;
структурные;
транспортные.
Транспортные и плазменные липопротеины.
Все липиды за исключением свободных жирных
Липопротеины в отличии от остальных липидов растворимы в воде, что определяется их строением.

Слайд 42
Липопротеины представляют собой сферические частицы (мицеллы), состоя-щие в наружном полярном слое
Классификация липопротеинов зависит от их плотности, а плотность – от содержания в них липидов. Чем больше липидов - тем меньше плотность.
, состоя-щие в наружном полярном слое из смеси белков, холестерина")
Слайд 43
Различают 4 класса плазменных липопротеинов:
хиломикроны;
липопротеины очень низкой плотности (ЛПОНП);
липопротеины низкой
липопротеины высокой плотности (ЛПВП).
Функции транспортных липопротеинов плазмы.
Все липопротеины выполняют транспортную роль. Они переносят экзогенные, всосавшиеся из кишечника триглицериды и холестерин в печень. От печени, где синтезируются эндогенные фосфолипиды и холестерин, липопротеины транспортируются ко всем внутренним органам, где используются. Поэтому липопротеины плазмы называют транспортными формами липидов.
Каждый класс липопротеинов транспортирует ту фракцию липидов, которая в нем преобладает.
;липопротеины низкой плотности (ЛПНП);липопротеины высокой плотности")

Слайд 45Фосфолипиды наружной оболочки липопротеинов связаны со специ-фическими белками - аполипо-протеинами или
Аполипопротеины связываются с фосфолипидами за счет гидрофобных взаимодействий между жирнокислотными цепями фосфолипидов и неполярными областями белковой структуры.
Ионные взаимодействия между полярными группами головок фосфолипидов и парами противоположно заряженных аминокислот в альфа-спиральных участках апопротеинов играют вторичную стабилизирующую роль.

Слайд 46Аполипопротеины выполняют три основные функции:
1. помогают солюбилизировать (обеспечи-вают растворимость) в
2. регулируют реакции липидов2. регулируют реакции липидов и липопротеинов2. регулируют реакции липидов и липопротеинов с ферментами (ЛХАТ , липо-протеинлипазой и печеночной липазой).
3. связываются с рецепторами3. связываются с рецепторами на поверхности клеток, запуская перенос липидов внутрь клетки и определяя таким образом места захвата холестерина .
 в воде эфиров холесте-рина и")
Слайд 47Химические свойства омыляемых липидов
Реакция гидролиза – гидролиз - первая стадия процесса

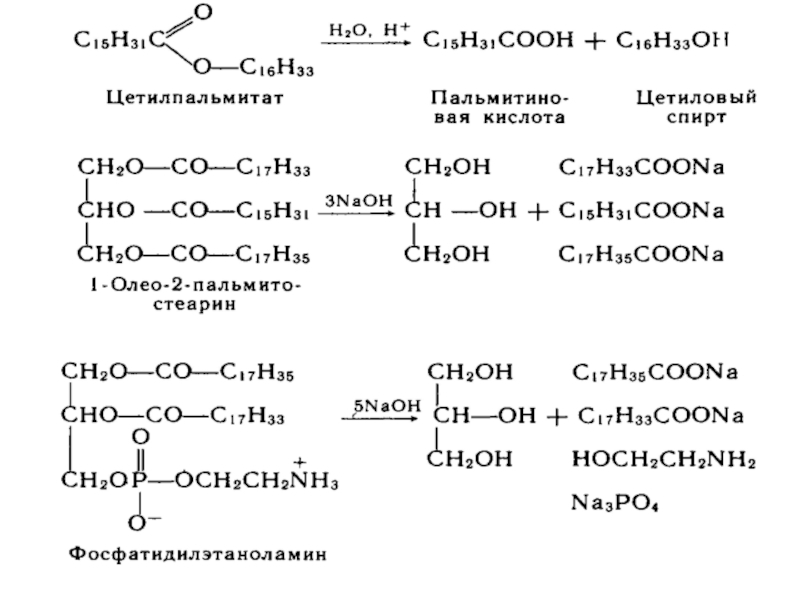
Слайд 49Реакция присоединения – Липиды содержащие остатки непредельных ЖК присоединяют по двойным


Слайд 51В промышленности широко применяется каталитическое гидрогенирование ненасыщенных растительных масел в результате

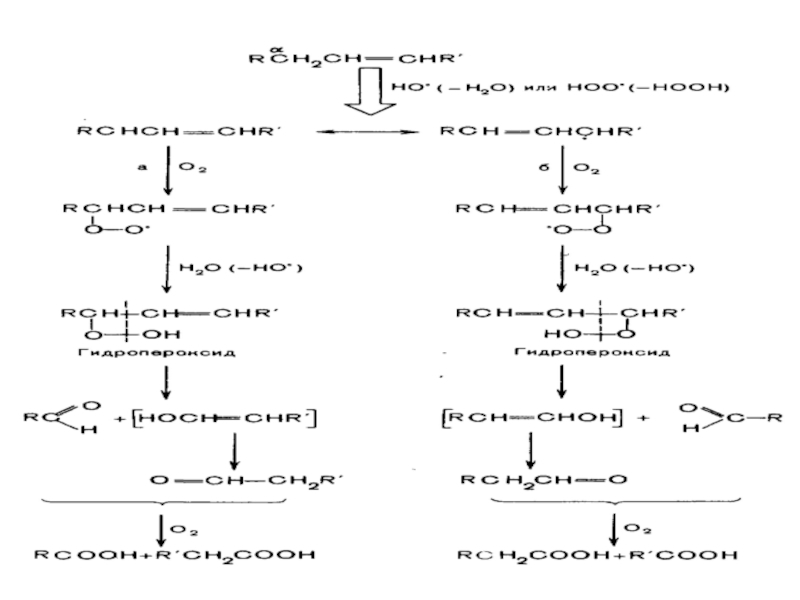
Слайд 52Реакции окисления
Процессы окисления липидов весьма разнообразны. В частности, окисление кислородом

Слайд 53Указанная реакция является начальной стадией процесса так называемого свободнорадикального пероксидного окисления
Fe2+ + H2O2→ Fe3+ + OH- + OH•
. В настоящее")
Слайд 55Липиды – важный энергетический субстрат. Высвобождение энергии, заключенной в химических связях

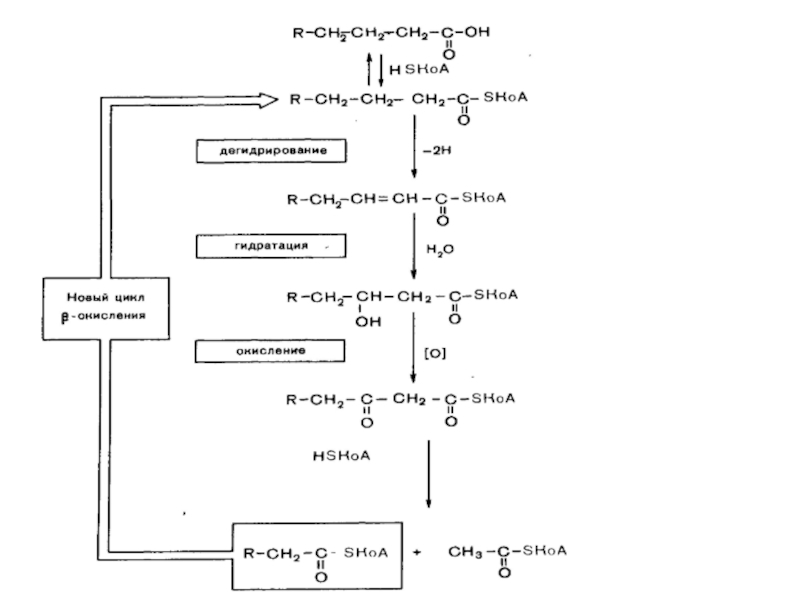
Слайд 57При атаке таким радикалом по метиленовой группе липида, соседней с двойной
Образовавшийся радикал может давать начало промежуточным пероксидам, которые будучи весьма нестабильными, легко распадаются с образованием продуктов альдегидной и кетонной природы, которые далее окисляются с образованием одно- и двухосновных карбоновых кислот – конечных продуктов реакции.

Слайд 58Неомыляемые липиды
Особую группу терпенов составляют каротиноиды – растительные пигменты. Некоторые из

Слайд 59К терпенам относятся и другие жирорастворимые витамины, такие как витамины группы
, витамины группы")





