- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Моделирование структуры биомакромолекул презентация
Содержание
- 1. Моделирование структуры биомакромолекул
- 2. Моделирование структуры биомакромолекул Для чего?
- 3. Структура белков определяет их функцию Моделирование структуры биомакромолекул
- 4. Уровни белковой организации
- 5. Первичная структура: последовательность Пептидная сязь
- 6. Первичная структура: последовательность
- 7. Первичная структура: последовательность
- 8. Вторичная структура: α-спирали, β-листы, петли
- 9. Вторичная структура: α-спирали
- 10. Вторичная структура: β-листы
- 11. Структурные мотивы
- 12. Третичная структура: домены
- 13. Мозаичная структура белков
- 14. Третичная структура: пространственная укладка белка (фолд)
- 15. Четвертичная структура: мультимерные белки и белковые комплексы
- 16. Сворачивание белка (фолдинг) Сворачивание белка - процесс
- 17. Нековалентные (“слабые”) взаимодействия Водородные
- 18. Гидрофобность аминокислот Гидрофобные эффекты играют важную роль
- 19. Определение пространственной структуры белка Экспериментальный подход
- 20. Преимущества метода рентгеноструктурного анализа. принципиально достижимо
- 21. Ограничения метода рентгеноструктурного анализа биомолекул Molecular
- 22. Схема рентгеноструктурного исследования
- 23. Наработка и очистка белка Выращивание
- 24. Снятие рентгенограмм кристаллов Molecular
- 25. Определение координат тяжелых атомов биомолекулы Molecular Conceptor v. 2.11, Synergix ltd., USA
- 26. Protein Structure Initiative (NIGMS, NIH, USA, 2001-2010,
- 27. Многомерная ЯМР спектроскопия Преимущества: молекулы в
- 28. Bruker Biospin AVANCE 1000 The World’s First
- 29. Ограничение метода многомерного ЯМР Molecular Conceptor
- 30. Физические принципы метода ядерного магнитного резонанса
- 31. Сбор данных Анализ, Соотнесение (assignment) Molecular Conceptor v. 2.11, Synergix ltd., USA
- 32. Sequential NOEs ("NOESY walks") in
- 33. Определение координат атомов молекулы Molecular Conceptor
- 34. Электронная микроскопия Определяется форма крупных
- 35. Банк белковых структур. Protein Data Bank (PDB) http://www.rcsb.org/pdb
- 36. https://www.dnastar.com/blog/structural-biology/why-structure-prediction-matters/
- 37. Ab initio - моделирование укладки “из первых
- 38. Предсказание структуры ab initio Функция потенциальной энергии
- 39. Предсказание структуры с использованием решетки
- 40. Предсказание структуры с использованием решетки
- 41. Используются структурно консервативные фрагменты длиной 4-10 аминокислот
- 42. Предсказание структуры на основе гомологии Выравнивание рассматриваемой
- 43. Примеры укладок (фолдов)
- 44. Предсказание структуры на основе гомологии
- 45. Тридинг (Threading) - предсказание структуры на основе
- 46. Основные компоненты тридинга библиотека уникальных укладок (фолдов)
- 47. CASP - конкурс методов предсказания структуры
- 48. Гомологическое моделирование третичной структуры белка на основе
- 49. Присвоение координат атомов В первую очередь
- 50. Поиск конформации соединяющих петель После того,
- 51. Построение пространственной структуры D-amino-acid oxidase из Trigonopsis
- 53. Типичная процедура регуляризации модельной структуры белка Энергетическая




Слайд 5Первичная структура: последовательность
Пептидная сязь
МОНОМЕР
ПОЛИМЕР
Аминокислота
Полипептид
Первичной структурой белка является его аминокислотная последовательность
>small ubiquitin-related
MSEEKPKEGVKTENDHINLKVAGQDGSVVQFKIKRHTPLSKLMKAYCERQG
LSMRQIRFRFDGQPINETDTPAQLEMEDEDTIDVFQQQTGGVPESSLAGHSF

Слайд 6
Первичная структура: последовательность
Двадцать аминокислот, встречающихся в белках, имеют различные свойства.


Слайд 8Вторичная структура: α-спирали, β-листы, петли
α-спирали и β-листы формируются путем образования водородных


Слайд 10Вторичная структура: β-листы
Антипараллельный β-лист
Параллельный β-лист
β-лист
β-тяж
Смешанный β-лист




")
Слайд 15Четвертичная структура: мультимерные белки и белковые комплексы
Гемоглобин
- тетрамер
Рибосома: РНК-белковый комплекс
- синтез
Реплисома
- копирование ДНК

Слайд 16Сворачивание белка (фолдинг)
Сворачивание белка - процесс укладки полипептидной цепи в компактную
Аминокислотная последовательность белка одназначно определяет его пространственную структуру [Anfinsen et al., 1960s].
Пространственная структура белка определяет его функцию.
Сворачивание белка - процесс укладки полипептидной цепи в компактную пространственную структуру.Аминокислотная последовательность белка")
Слайд 17Нековалентные (“слабые”) взаимодействия
Водородные связи
Ионные связи
Гидрофобные взаимодействия
Ван-дер-Ваальсовы взаимодействия
 взаимодействияВодородные связиИонные связиГидрофобные взаимодействияВан-дер-Ваальсовы взаимодействия")
Слайд 18Гидрофобность аминокислот
Гидрофобные эффекты играют важную роль в сворачивании белка
Экспериментально измеренные уровни

Слайд 19Определение пространственной структуры белка
Экспериментальный подход
Вычислительный подход
- Предсказание пространственной структуры белка на
- Рентгеноструктурный анализ
- Метод многомерного ЯМР
- Метод криоэлектронной микроскопии
Molecular Conceptor v. 2.11, Synergix ltd., USA

Слайд 20Преимущества метода рентгеноструктурного анализа.
принципиально достижимо высокое разрешение. Разрешение выше 1Å позволяет
возможность разрешать структуры объектов большого размера (вирусная капсида, рибосома, фотосинтетический реакционный центр, т.д.), состоящих из нескольких десятков тысяч атомов.
Molecular Conceptor v. 2.11, Synergix ltd., USA

Слайд 21Ограничения метода рентгеноструктурного анализа биомолекул
Molecular Conceptor v. 2.11, Synergix ltd., USA
20


Слайд 23Наработка и очистка белка
Выращивание кристалл(а/ов)
Molecular Conceptor v. 2.11, Synergix ltd., USA
Molecular Conceptor v. 2.11, Synergix ltd., USA")
Слайд 24Снятие рентгенограмм кристаллов
Molecular Conceptor v. 2.11, Synergix ltd., USA
Регулярный Кристалл Размером

Слайд 25Определение координат тяжелых атомов биомолекулы
Molecular Conceptor v. 2.11, Synergix ltd., USA

Слайд 26Protein Structure Initiative (NIGMS, NIH, USA, 2001-2010, 2011-2015)
Выбор объекта
$750M
Экспрессия белка
С высоким
Высокоэффективная
очистка
Кристаллизация
Отбор кристаллов, Сбор данных
4 крупных и 6 малых центров разрешили за 7 лет более 3000 белковых структур (40% новых структур)
Полуавтоматическое
Разрешение структуры
Валидация стр-ры, Помещение в PDB
Полуавтоматическая
публикация
Новая структура каждые 2 дня!
GroEL за 2 месяца.
Service R.B., Science 319, 1610 (2008)
Выбор объекта$750MЭкспрессия белкаС высоким выходомВысокоэффективнаяочисткаКристаллизацияОтбор кристаллов, Сбор данных4")
Слайд 27Многомерная ЯМР спектроскопия
Преимущества:
молекулы в растворе (тяжѐлая вода), не нужно выращивать кристалл
положения
информация о динамике
атомов м.б. определена
В бел ках 103 – 104 протонов Метильные и метиленовые группы 0,8-3,5 ppm, ароматические, индольные и иммидазольные кольца 6,5-8 ppm
В ДНК/РНК Н-2/Н-8 пуринов 8,4-
9 ppm, Н-5 пиримидинов 6,3-6,6 ppm, Н-6 8,0-8,5 ppm, метильная группа тимидина 2,3-2,4 ppm.
Molecular Conceptor v. 2.11, Synergix ltd., USA
, не нужно выращивать кристаллположения атомов водорода м.б. определеныинформация")
Слайд 28Bruker Biospin AVANCE 1000
The World’s First 1 Gigahertz NMR Spectrometer
World’s First
23.5 Tesla Standard-Bore, Persistent Superconducting Magnet (12-tonne, 4.5-metre-tall machine)
Dr. Lyndon Emsley, European Centre for High Field NMR (CRMN) in Lyon, France.
Nature 463,605(2010).
€11.7-million (US$16.3-million)
http://www.bruker-biospin.com/av1000-dir.html

Слайд 29Ограничение метода многомерного ЯМР
Molecular Conceptor v. 2.11, Synergix ltd., USA
Структура белков
Дороговизна получения образцов с изменѐнным изотопным составом
(13С, 19F, 31P)
Невысокая точность разрешения структуры

Слайд 30Физические принципы метода ядерного магнитного резонанса
Характерные спектры химических групп и соединений
Molecular

Molecular Conceptor v. 2.11, Synergix ltd., USA")
Слайд 32
Sequential NOEs ("NOESY walks") in aromatic- H1'/H5 region of TWJ-TC acquired
Leontis, N. et al., Biophysical Journal, 68, 251 (1995).

Слайд 33Определение координат атомов молекулы
Molecular Conceptor v. 2.11, Synergix ltd., USA
Для структур

Слайд 34Электронная микроскопия
Определяется форма крупных межмолекулярных комплексов методом диффракции электронных пучков.
Типичное разрешение
Образцы в замороженном состоянии, что предотвращает радиационные повреждения и удерживает их в нативном состоянии.
Molecular Conceptor v. 2.11, Synergix ltd., USA

Слайд 35Банк белковых структур. Protein Data Bank
(PDB)
http://www.rcsb.org/pdb
Research Collaboratory for Structural Bioinformatics
Каждая структура имеет
файл, в котором приведены координаты тяжѐлых атомов.
Структура PDB файла. (Brookhaven Protein Data Bank)
http://www.rcsb.org/pdbResearch Collaboratory for Structural BioinformaticsКаждая структура имеет свой идентификатор (4 символа)")

Слайд 37Ab initio - моделирование укладки “из первых принципов” - без использования
Предсказание на основе гомологии (homology modeling) -
моделирование на основе известных структур схожих белков.
Тридинг (Threading) - моделирование на основе слабой гомологии.
Предсказание пространственной структуры белка

Слайд 38Предсказание структуры ab initio
Функция потенциальной энергии
Модель водного раствора
Оценка попарного взаимодействия между
Поиск в пространстве всевозможных конформаций
Модель на основе “решетки”
Молекулярная динамика
Использование библиотек известных 3D фрагментов
Предсказание вторичной структуры

Слайд 39Предсказание структуры с использованием решетки
HP-модель (Hydrophobic-Polar) - рассматривает гидрофобные взаимодействия как
Не существует эффективных алгоритмов
Плохо отражает реальность
 - рассматривает гидрофобные взаимодействия как наиболее важные.Не существует эффективных")

Слайд 41Используются структурно консервативные фрагменты длиной 4-10 аминокислот
Поиск в пространстве конформаций осуществляется
Полученные структуры кластеризуются и в качестве результата выдаются наилучшие структуры для каждого кластера
ROSETTA

Слайд 42Предсказание структуры на основе гомологии
Выравнивание рассматриваемой последовательности с последовательностями белков с
Наложение моделируемой последовательности на известную структуру согласно выравниванию
Локальное улучшение полученной пространственной структуры
Число уникальных укладок (фолдов), наблюдающихся в белках, ограничено
(несколько тысяч)
90% помещаемых в PDB структур имеют уже известные укладки (фолды)

")
Слайд 44Предсказание структуры на основе гомологии
Raw model
Loop modeling
Side chain placement
Refinement

Слайд 45Тридинг (Threading) - предсказание структуры на основе слабой гомологии
MTYKLILN …. NGVDGEWTYTE
Главное
 - предсказание структуры на основе слабой гомологииMTYKLILN …. NGVDGEWTYTEГлавное отличие от моделирования по")
Слайд 46Основные компоненты тридинга
библиотека уникальных укладок (фолдов)
функция, определяющая вес выравнивания последовательности со
алгоритм нахождения наилучшего выравнивания
функция, определяющая вес выравнивания последовательности со структуройалгоритм нахождения наилучшего выравнивания")
Слайд 47
CASP - конкурс методов предсказания структуры белков
Critical Assessment of protein Structure
FoldIt

Слайд 48Гомологическое моделирование третичной структуры
белка на основе первичной структуры
Стратегия построения пространственной структуры
Определения круга гомологичных белков;
Нахождение структурно-консервативных элементов в структуре гомологов (SCRs);
Выравнивание последовательности модельного белка с последовательностями гомологов, с учѐтом наличия SCR;
Присвоение координат атомов остатков, входящих в SCR, соответствующим атомам модельного белка согласно выравниванию;
Предсказание конформации петель, соединяющих SCR, а также N- и С-концов пептидной цепи белка;
Поиск оптимальной конформации боковых остатков аминокислот модельного
белка, отличающихся от остатков опорного белка;
Использование методов регуляризации структуры (энергетическая минимизация и молекулярная динамика) для уточнения молекулярной структуры с целью устранения стерических напряжений созданных при построении моделей.

Слайд 49Присвоение координат атомов
В первую очередь присваиваются координаты атомам полипептидной цепи.
Затем присваиваются
вероятно вызывая сильные напряжения в структуре модельного белка.

Слайд 50Поиск конформации соединяющих петель
После того, как присвоены координаты атомам, составляющим петли,
Геометрия пептидной цепи модельной структуры должна быть регулярной (транс- конформация пептидных групп, близкие к равновесным значения валентных углов и дли связей);
Атомы не должны перекрываться, т.е. расстояния между несвязанными атомами не должны быть существенно меньше, чем сумма их ван-дер-ваальсовских радиусов;
Боковые цепи аминокислот должны находиться в равновесной конфигурации;
▪Если в молекуле имеются дисульфидные мостики (Cys-Cys связи), то расстояния между соответствующими атомами серы должны быть приведены в соответствие с геометрией;
В структуру должны быть помещены необходимые простетические группы.

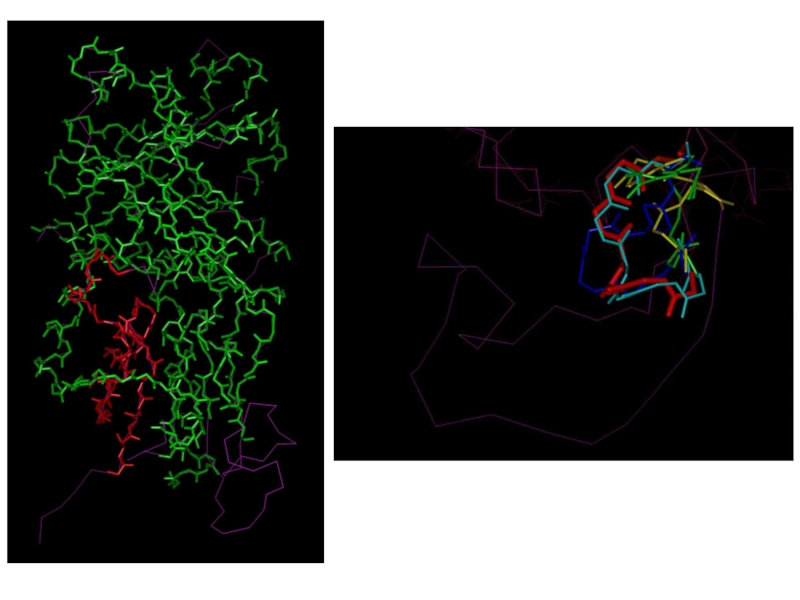
Слайд 51Построение пространственной структуры D-amino-acid
oxidase из Trigonopsis variabilis (Yeast)
В качестве опорного белка
В качестве опорного белка была использована пространственная структура")
Слайд 53Типичная процедура регуляризации модельной структуры белка
Энергетическая минимизация участков сочленения SCR и
Энергетическая минимизация пептидной цепи и боковых остатков петель;
Энергетическая минимизация боковых цепей аминокислот, принадлежащих SCR, подвергшихся замене при присваивании координат;
Энергетическая минимизация всех боковых остатков белка;
Энергетическая минимизация (500-1000 шагов) всей структуры модельного белка;
Молекулярная динамика модельного белка в вакууме на протяжении 20-50 пикосекунд;
Финальная энергетическая минимизация структуры белка (200-500 шагов).
Результатом этой процедуры будет белковая структура с прав ильно й стереох имией (длины валентных связей и значения валентных углов не будут существенно отличаться от равновесных значений), с отрицательной энергией несвязанных взаимодействий (свидетельство того, что не наблюдается перекрытие ван-дер-ваальсовских радиусов атомов), с отрицательной энергией электростатических взаимодействий (произошло сближение противоположно заряженных атомов) и с ненулевой энергией водородных связей (в молекуле установились водородные связи).
Дальнейшая регуляризация структуры приведѐт к еѐ улучшению с точки зрения стереохимии, но
при этом возрастут искажения структуры активного центра (центра связывания) вашей структуры.
Модельная ст руктур а по ст р о ена и отрелаксирована. Она обладает участками структурно- консервативных областей, унаследованных от белков гомологов, правильной стереохимией (результат регуляризации). Дальнейшие манипуляции с этой структурой (подгонка геометрии активного центра, точечные мутации) зависят от цели исследований. Полученную структуру надо рассматривать как средство иллюстрации результатов вашей работы (объяснения экспериментальных фактов, гипотезы).