- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Зеленые водоросли презентация
Содержание
- 1. Зеленые водоросли
- 2. ОТДЕЛ ЗЕЛЕНЫ Е ВОДОРОСЛИ характеризуются травянисто-зеленой окраской
- 3. ОТДЕЛ ЗЕЛЕНЫЕ ВОДОРОСЛИ Пиреноид представляет собой плотное
- 4. ОТДЕЛ ЗЕЛЕНЫЕ ВОДОРОСЛИ Глазок располагается внутри хлоропласта
- 8. ОТДЕЛ ЗЕЛЕНЫ Е
- 9. Систематика Класс собственно зеленые водоросли, или равножгутиковые
- 10. Класс собственно зеленые водоросли
- 11. Класс харовые водоросли
- 12. КЛАСС СОБСТВЕННО ЗЕЛЕНЫЕ Порядок вольвоксовые Организация: монадные
- 14. переходят пальмеллевидное состояние: теряют жгутики
- 15. размножение в благоприятных условиях - бесполое размножение
- 17. Разнообразие отсутствие клеточной стенки - род дуналиелла,
- 18. Порядок тетраспоровые —пальмеллоидный тип строения таллома .
- 20. Представители род апиоцистис - грушевидные
- 22. Порядок хлорококковые объединяет коккоидные формы: одноклеточные,
- 23. Род гидродикцион или водяна я сеточка
- 25. Порядок улотриксовые зеленые водоросли - таллом в
- 26. При бесполом размножении во всех зеленых
- 28. К порядку улотриксовые в качестве отдельного
- 33. Ульва и энтероморфа - морские водоросли,
- 34. Одни растения - бесполое поколение -
- 35. Порядок хетофоровые гетеротрихальное строение таллома, дифференцированы на
- 36. Порядок сифоновые сифоновый тип строения, -
- 38. Бриопсис Каулерпа кодиума - макроскопически
- 39. Кодиум
- 41. дазикладус Ацетабулярия
- 42. Порядок сифонокладовые многоклеточные талломы , из многоядерных клеток род кладофора
- 44. Бесполое размножение - зооспоры двух- или
- 46. КЛАСС КОНЪЮГАТЫ, ИЛИ СЦЕПЛЯНКИ одноклеточные и нитчатые
- 47. Класс конъюгаты Порядок мезотениевые Порядок зигнемовые Порядок десмидиевые
- 48. Порядок мезотениевые Одноклеточные формы. Клетки цилиндрические,
- 50. Порядок зигнемовые нитчатые пресноводные водоросли.
- 51. Половой процесс — конъюгация. Наиболее
- 53. Порядок десмидиевые одноклеточные, реже нитчатые водоросли.
- 55. КЛАСС ХАРОВЫЕ представлен всего одним порядком
- 58. КЛАСС ХАРОВЫЕ Клетки одеты целлюлозной оболочкой,
- 59. Размножение харовых Оогоний направлен вверх, антеридий —
- 60. Клетка базального узла антеридия, которая развивается
- 62. Внутренние стенки нитей утолщаются , опробковевают,

Слайд 2ОТДЕЛ ЗЕЛЕНЫ Е ВОДОРОСЛИ
характеризуются травянисто-зеленой окраской хлоропластов
преобладание хлорофиллов «а» и
«б» над
каротиноидами : а - и б-каротин, лютеином, неоксантином, виолаксантином , зеаксантином , антераксантином.
Хлоропласты окружены оболочкой из двух мембран
Ламеллы состоят из двух - шести или многих слившихся тилакоидов, образуют граны
каротиноидами : а - и б-каротин, лютеином, неоксантином, виолаксантином , зеаксантином , антераксантином.
Хлоропласты окружены оболочкой из двух мембран
Ламеллы состоят из двух - шести или многих слившихся тилакоидов, образуют граны

Слайд 3ОТДЕЛ ЗЕЛЕНЫЕ ВОДОРОСЛИ
Пиреноид представляет собой плотное образование белковой природы, окруженное снаружи
обкладкой в виде сплошного кольца или отдельных пластинок в числе от двух и более, обычно крахмальной природы
Пиреноид, погруженный в строму хлоропласта , пронизан тилакоидами .
Крахмал - внутри хлоропласта — вокруг пиреноида и в строме.
Пиреноид, погруженный в строму хлоропласта , пронизан тилакоидами .
Крахмал - внутри хлоропласта — вокруг пиреноида и в строме.

Слайд 4ОТДЕЛ ЗЕЛЕНЫЕ ВОДОРОСЛИ
Глазок располагается внутри хлоропласта и не связан со жгутиковым
аппаратом .
Жгутик и в числе двух, четырех, реже многих одинаковой длины (изоконтные) и одинакового строения (изоморфные)
Клетка поверх ЦПМ покрыта целлюлозной оболочкой.
Размножение вегетативное, бесполое и половое.
Имеются все возможные циклы развития (гаплонты, диплонты, изо- и гетероморфная смены генераций) .
Жгутик и в числе двух, четырех, реже многих одинаковой длины (изоконтные) и одинакового строения (изоморфные)
Клетка поверх ЦПМ покрыта целлюлозной оболочкой.
Размножение вегетативное, бесполое и половое.
Имеются все возможные циклы развития (гаплонты, диплонты, изо- и гетероморфная смены генераций) .


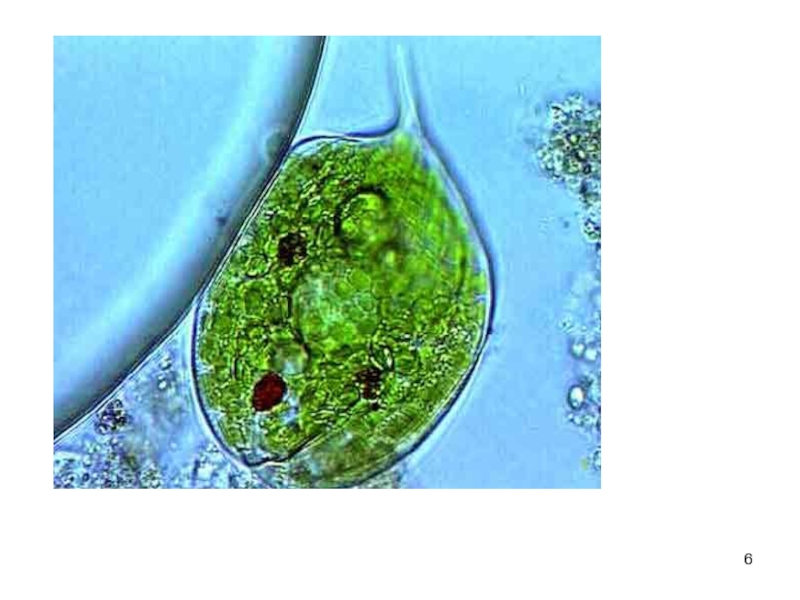

Слайд 8
ОТДЕЛ ЗЕЛЕНЫ Е ВОДОРОСЛИ
Класс собственно зеленые водоросли, или равножгутиковые
Класс конъюгаты
Класс харовые
водоросли

Слайд 9Систематика
Класс собственно зеленые водоросли, или равножгутиковые - характеризуется бесполым размножением обычно
зооспорами с двумя - четырьмя, реже многими изоконтными и изоморфными жгутиками, обращенными вперед, иногда апланоспорами .
Класс конъюгаты характеризуется отсутствием в жизненном цикле жгутиковых стадий, так как бесполое размножение отсутствует, а половой процесс — конъюгация.
Класс харовые водоросли включае т крупные водоросли со сложной морфологической дифференцировкой нитчатого таллома. Бесполого размножения нет. Половой процесс оогамный. Половые органы — оогонии и антеридии — многоклеточные, сложного строения.
Класс конъюгаты характеризуется отсутствием в жизненном цикле жгутиковых стадий, так как бесполое размножение отсутствует, а половой процесс — конъюгация.
Класс харовые водоросли включае т крупные водоросли со сложной морфологической дифференцировкой нитчатого таллома. Бесполого размножения нет. Половой процесс оогамный. Половые органы — оогонии и антеридии — многоклеточные, сложного строения.

Слайд 10
Класс собственно зеленые водоросли
Порядок вольвоксовые
Порядок тетраспоровые
Порядок хлорококковые
Порядок улотриксовые
Порядок хетофоровые
Порядок сифоновые


Слайд 12КЛАСС СОБСТВЕННО ЗЕЛЕНЫЕ
Порядок вольвоксовые
Организация:
монадные (снабженные жгутиками одноклеточные)
колониальные и ценобиальные
Типичный
представитель — одноклеточная водоросль из рода хламидомонада - в лужах , канавах, мелких пресных водоемах.
Сферическая или эллипсоидальная клетка
Оболочка
передний конец - два жгутика
Протопласт содержит
одно ядро
чашевидный хроматофор
пиреноид и глазок
сократительные вакуоли
Сферическая или эллипсоидальная клетка
Оболочка
передний конец - два жгутика
Протопласт содержит
одно ядро
чашевидный хроматофор
пиреноид и глазок
сократительные вакуоли
 колониальные и ценобиальные Типичный представитель — одноклеточная водоросль")
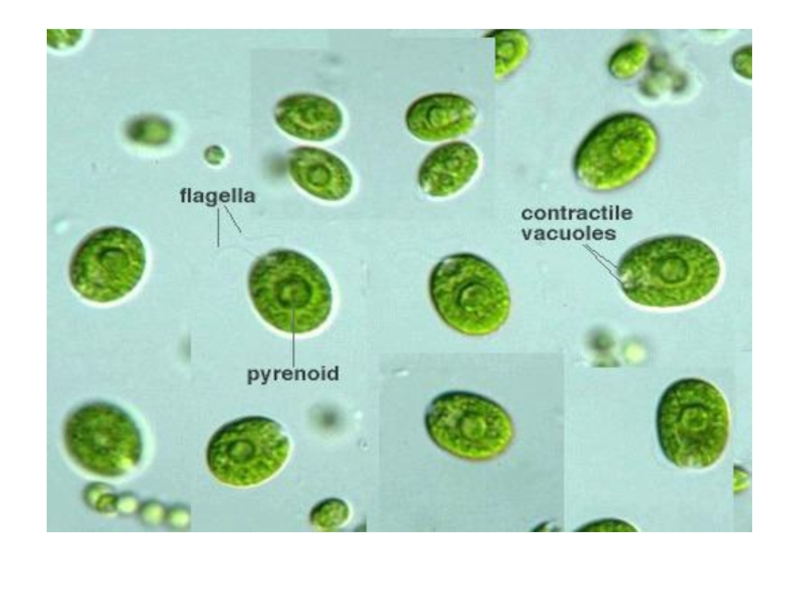
Слайд 14
переходят пальмеллевидное состояние:
теряют жгутики
стенка ослизняется
размножаются - система вложенных слизистых обверток
- неподвижные клетки

Слайд 15размножение
в благоприятных условиях - бесполое размножение - зооспоры
Половой процесс - изогамный.
У некоторых вольвоксовых гетерогамия и оогамия.

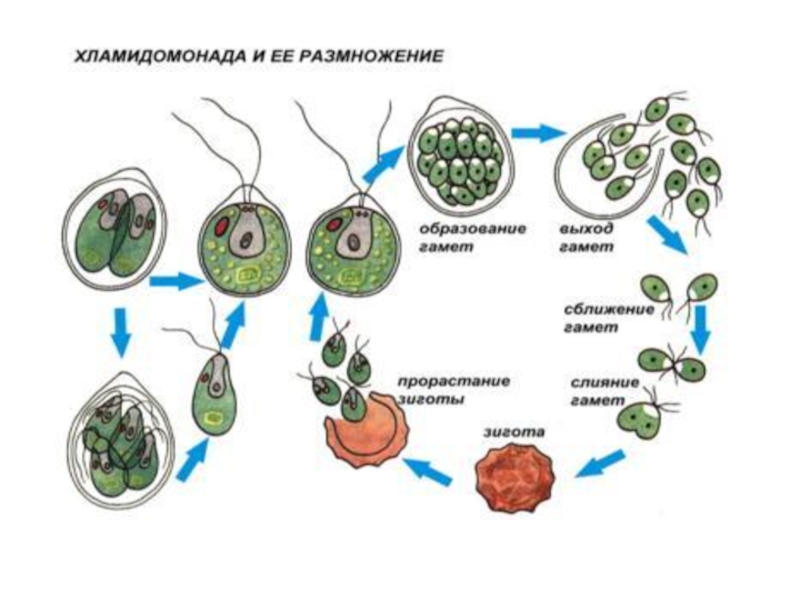
Слайд 17Разнообразие
отсутствие клеточной стенки - род дуналиелла, встречающийся в соленых озерах похож
на хламидомонаду
род астеромонас
У многих вольвоксовых клетки, имеющие строение хламидомонады соединяются в подвижные ценобии неодинаковой формы, состоящие из различного числа клеток.
Из ценобиальных представителей вольвоксовых чаще всего встречаются роды гониум, пандорина , эвдорина, вольвокс.
род астеромонас
У многих вольвоксовых клетки, имеющие строение хламидомонады соединяются в подвижные ценобии неодинаковой формы, состоящие из различного числа клеток.
Из ценобиальных представителей вольвоксовых чаще всего встречаются роды гониум, пандорина , эвдорина, вольвокс.


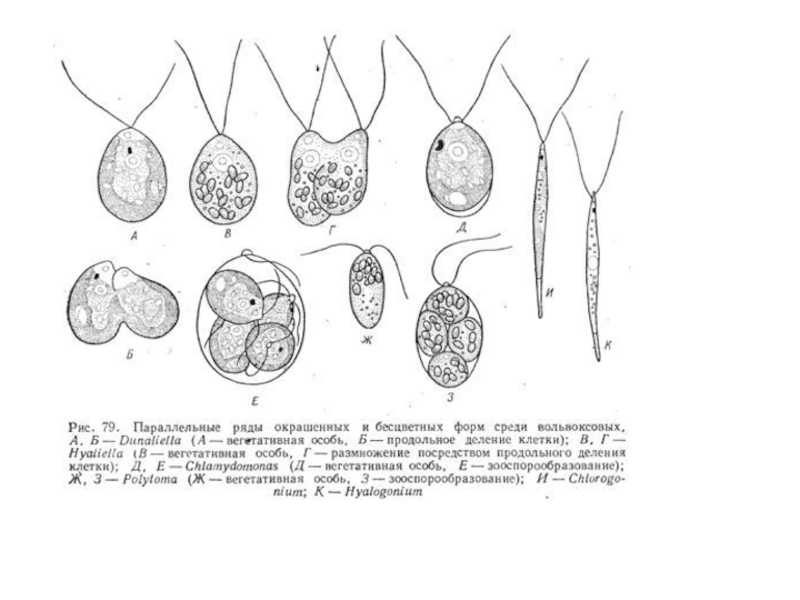
Слайд 20
Представители
род апиоцистис - грушевидные слизистые колонии, прикрепленные к водорослям
род
тетраспора - мешковидные, бесформенные, крупные, слизистые колонии.
В слизи - клетки, по строению напоминающие хламидомонаду.
В слизи - клетки, по строению напоминающие хламидомонаду.

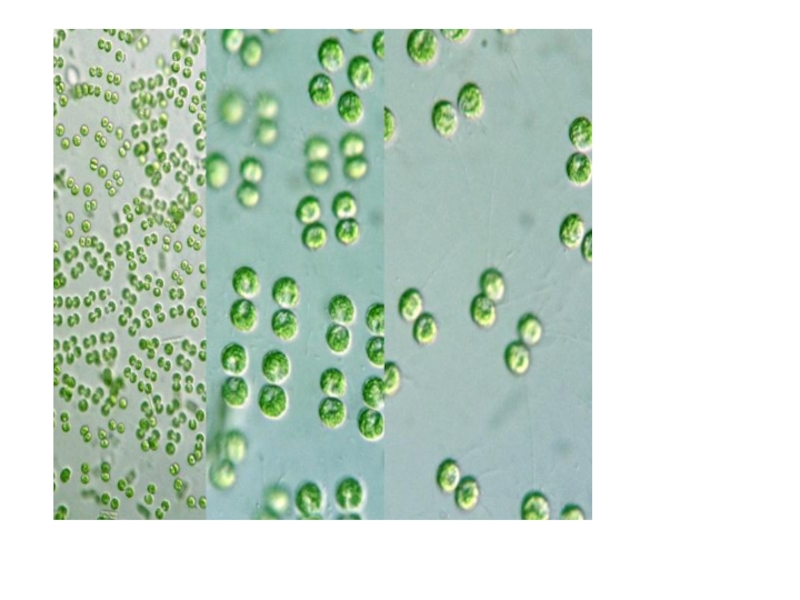
Слайд 22Порядок хлорококковые
объединяет коккоидные формы: одноклеточные, колониальные и ценобиальные.
Клетка - строение
вольвоксовых
Бесполое размножение — посредством зооспор и апланоспор (автоспор) .
У ценобиальных форм зооспоры и автоспоры внутри оболочки материнской клетки слагаются в дочерние ценобии
растут за счет увеличения размеров клетки, а число клеток остается постоянным.
Половой процесс изо-, гетеро- и оогамный.
Бесполое размножение — посредством зооспор и апланоспор (автоспор) .
У ценобиальных форм зооспоры и автоспоры внутри оболочки материнской клетки слагаются в дочерние ценобии
растут за счет увеличения размеров клетки, а число клеток остается постоянным.
Половой процесс изо-, гетеро- и оогамный.



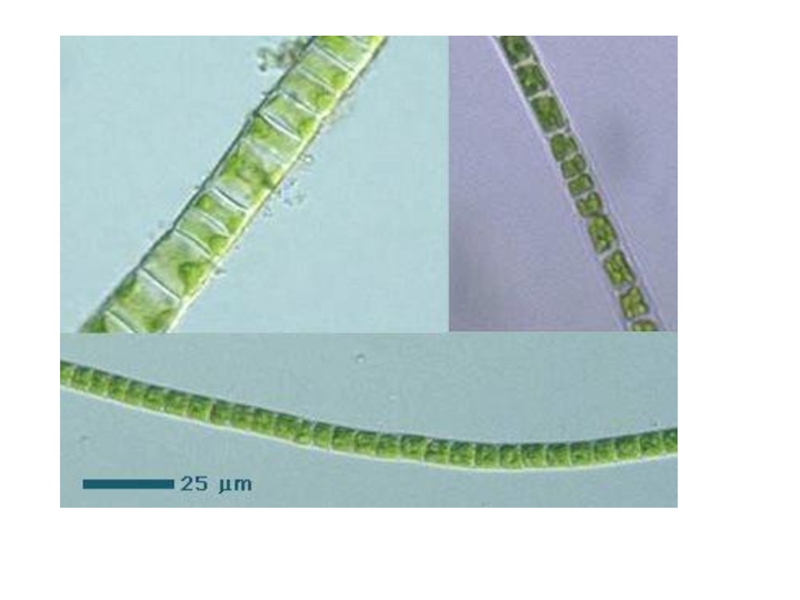
Слайд 25Порядок улотриксовые
зеленые водоросли - таллом в виде неразветвленной нити, сложенной из
одноядерных клеток.
Реже - пластинчатый или трубчатый, но в онтогенезе развивается из нитчатого.
Представитель род улотрикс
Под Москвой эта водоросль встречается сравнительно редко в быстро текущих ручьях. Севернее, например в Неве, в прибойной поло-се больших озер часто образует ватообразны е обрастани я на камнях.
Нить - из одного ряд а клеток, одетых толстой оболочкой, под которой находится постенная цито-плазма , содержаща я пластинчатый хроматофор, опоясывающий клетку изнутри в виде незамкнутого кольца.
Все клетки нити одинаковые, за исключением базальной, - короткий ризоид для прикрепления к субстрату.
Все клетки, кроме базальной, способны делиться - непрерывное нарастание таллома , образовывать репродуктивные клетки.
Реже - пластинчатый или трубчатый, но в онтогенезе развивается из нитчатого.
Представитель род улотрикс
Под Москвой эта водоросль встречается сравнительно редко в быстро текущих ручьях. Севернее, например в Неве, в прибойной поло-се больших озер часто образует ватообразны е обрастани я на камнях.
Нить - из одного ряд а клеток, одетых толстой оболочкой, под которой находится постенная цито-плазма , содержаща я пластинчатый хроматофор, опоясывающий клетку изнутри в виде незамкнутого кольца.
Все клетки нити одинаковые, за исключением базальной, - короткий ризоид для прикрепления к субстрату.
Все клетки, кроме базальной, способны делиться - непрерывное нарастание таллома , образовывать репродуктивные клетки.

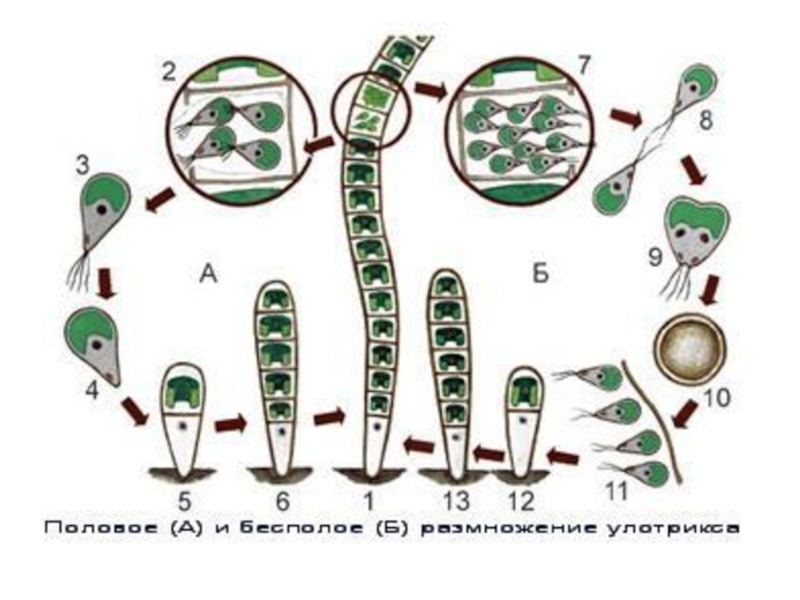
Слайд 26
При бесполом размножении во всех зеленых клетках развивается от двух до
16(32) четырехжгутиковых зооспор.
При половом размножении - двужгутиковые гаметы (4)—8—32(64)
Гаметы выходят из клетки, заключенные в слизистый пузырь, в котором они двигаются,
Половой процесс изогамный
При половом размножении - двужгутиковые гаметы (4)—8—32(64)
Гаметы выходят из клетки, заключенные в слизистый пузырь, в котором они двигаются,
Половой процесс изогамный
 четырехжгутиковых зооспор. При")
Слайд 28
К порядку улотриксовые в качестве отдельного семейства относят ульвовые. Представители этой
группы отличаются тем, что имеют не нитчатый, а пластинчатый таллом .
на ранних стадиях развития - талломы их состоят из прикрепленной к субстрату нити,
У близкого к ульве рода энтероморфа - вид кишки или трубки
на ранних стадиях развития - талломы их состоят из прикрепленной к субстрату нити,
У близкого к ульве рода энтероморфа - вид кишки или трубки



Слайд 33
Ульва и энтероморфа - морские водоросли,
приуроченные к прибрежной (литоральной) зоне
как в северных, так и в южных морях. Многие виды обоих родов выносят заметное опреснение воды и часто поднимаются в устья рек, а некоторые виды энтероморфы обычны и в пресноводных водоемах:
Вегетативное размножение - участками таллома
Бесполое размножение — зооспорами, как правило четырехжгутиковыми, которые образуются путем последовательного деления протопласта любой клетки на четыре - восемь частей.
Половое размножение - двужгутиковыми гаметами. Изогамия или гетерогамия.
Вегетативное размножение - участками таллома
Бесполое размножение — зооспорами, как правило четырехжгутиковыми, которые образуются путем последовательного деления протопласта любой клетки на четыре - восемь частей.
Половое размножение - двужгутиковыми гаметами. Изогамия или гетерогамия.
 зоне как в северных, так")
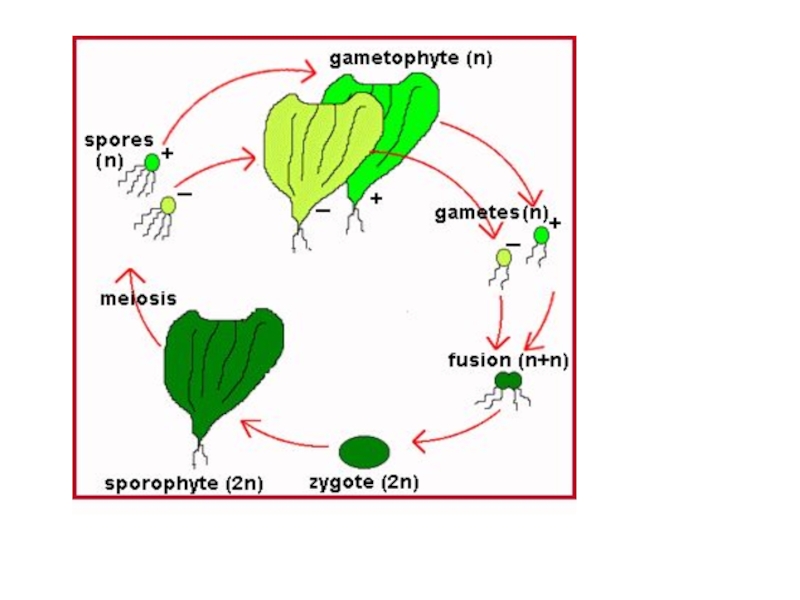
Слайд 34
Одни растения - бесполое поколение - спорофиты (2n) – мейоз –
зооспора (n)
Гаметофиты (n) – половое поколение – гаметы (n) - зигота.
Внешне (морфологически) оба поколения друг от друга неотличимы - изоморфная смена поколений.
И ульва , и энтероморфа — гетероталличные формы - гамето-фиты различны в половом отношении: одни продуцируют гаметы со зна-ком «+» , другие — со знако м «—».
Копуляция происходит, как и у улотрикса , лишь при встрече гамет разного знака .
У гетерогамных видов таллом ы разного знака можно различить и невооруженным глазом по окраске плодущих участков: желтовато й у мужски х и темно-зеленой у женских.
Гаметофиты (n) – половое поколение – гаметы (n) - зигота.
Внешне (морфологически) оба поколения друг от друга неотличимы - изоморфная смена поколений.
И ульва , и энтероморфа — гетероталличные формы - гамето-фиты различны в половом отношении: одни продуцируют гаметы со зна-ком «+» , другие — со знако м «—».
Копуляция происходит, как и у улотрикса , лишь при встрече гамет разного знака .
У гетерогамных видов таллом ы разного знака можно различить и невооруженным глазом по окраске плодущих участков: желтовато й у мужски х и темно-зеленой у женских.
 – мейоз – зооспора (n)Гаметофиты (n) –")
Слайд 35Порядок хетофоровые
гетеротрихальное строение таллома, дифференцированы на
распростертую по субстрату систему нитей
вертикально
стоящие, более или менее разветвленные нити.
Род стигеоклониум. Виды рода распространены в стоячей или медленно текущей воде на подводных предметах,
Род стигеоклониум. Виды рода распространены в стоячей или медленно текущей воде на подводных предметах,

Слайд 36Порядок сифоновые
сифоновый тип строения, - отсутствие клеточных перегородок,
возникают при основании
органов размножения
Таллом крупных размеров - одна клетка ,
Пигменты обычны, содержа т два особых ксантофилла — сифонеин и сифоноксантин,
Под хроматофорам - многочисленные ядра .
Размножение вегетативное и половое. Бесполого нет.
Половой процесс гетерогамный, реже изогамный.
Большинство - 2n,
обитатели тропических морей
Таллом крупных размеров - одна клетка ,
Пигменты обычны, содержа т два особых ксантофилла — сифонеин и сифоноксантин,
Под хроматофорам - многочисленные ядра .
Размножение вегетативное и половое. Бесполого нет.
Половой процесс гетерогамный, реже изогамный.
Большинство - 2n,
обитатели тропических морей


Слайд 38
Бриопсис
Каулерпа
кодиума - макроскопически крупные талломы возникают в результате плотного переплетения
простых нитей типичного сифонового строения. Так образуются крупные компактны е псевдопаренхиматозные талломы .
Таллом , прикрепленный к субстрат у базальным диском, имеет вид цилиндрических разветвленных темно-зеленых шнуров, достигающих длины 50 см при диаметр е 8 мм.
Таллом , прикрепленный к субстрат у базальным диском, имеет вид цилиндрических разветвленных темно-зеленых шнуров, достигающих длины 50 см при диаметр е 8 мм.






Слайд 44
Бесполое размножение - зооспоры двух- или четырехжгутиковые, выходят через боковое отверстие
в стенке.
Половой процесс изогамный, гаметы двужгутиковые, мельче зооспор.
У ряда морских видов кладофоры установлена изоморфная смена генераций. У этих видов гаметы и зооспоры всегда, развиваютс я на разных растениях: первые на гаплоидных — гаметофитах, вторые на диплоидных — спорофитах. Перед образованием зооспор ядро редукционно делится. Зооспоры развиваются в гаплоидные гаметофиты
Половой процесс изогамный, гаметы двужгутиковые, мельче зооспор.
У ряда морских видов кладофоры установлена изоморфная смена генераций. У этих видов гаметы и зооспоры всегда, развиваютс я на разных растениях: первые на гаплоидных — гаметофитах, вторые на диплоидных — спорофитах. Перед образованием зооспор ядро редукционно делится. Зооспоры развиваются в гаплоидные гаметофиты


Слайд 46КЛАСС КОНЪЮГАТЫ, ИЛИ СЦЕПЛЯНКИ
одноклеточные и нитчатые зеленые водоросли
отсутствие в
жизненном цикле жгутиковых стадий,
бесполого размножения нет
половой процесс — конъюгация — слияние амебоидных протопластов обычных вегетативных клеток.
распространено вегетативное размножение
бесполого размножения нет
половой процесс — конъюгация — слияние амебоидных протопластов обычных вегетативных клеток.
распространено вегетативное размножение


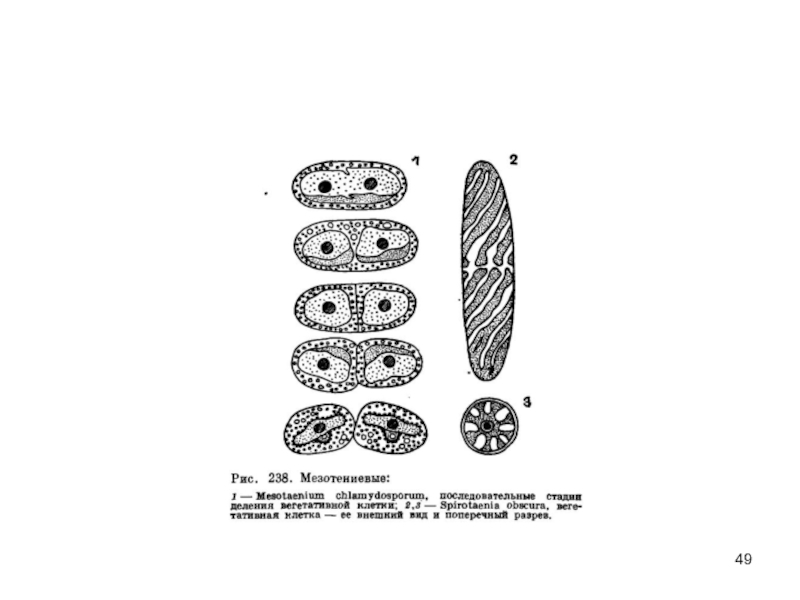
Слайд 48Порядок мезотениевые
Одноклеточные формы. Клетки цилиндрические, с закругленными концами
Слизистая обвертка
Одноядерные клетки
содержат хроматофоры , различающиеся у разных родов по форме.
распространены в торфяных болотах, некоторые виды
встречаются на сырой земле, образуя слизистые скопления.
распространены в торфяных болотах, некоторые виды
встречаются на сырой земле, образуя слизистые скопления.

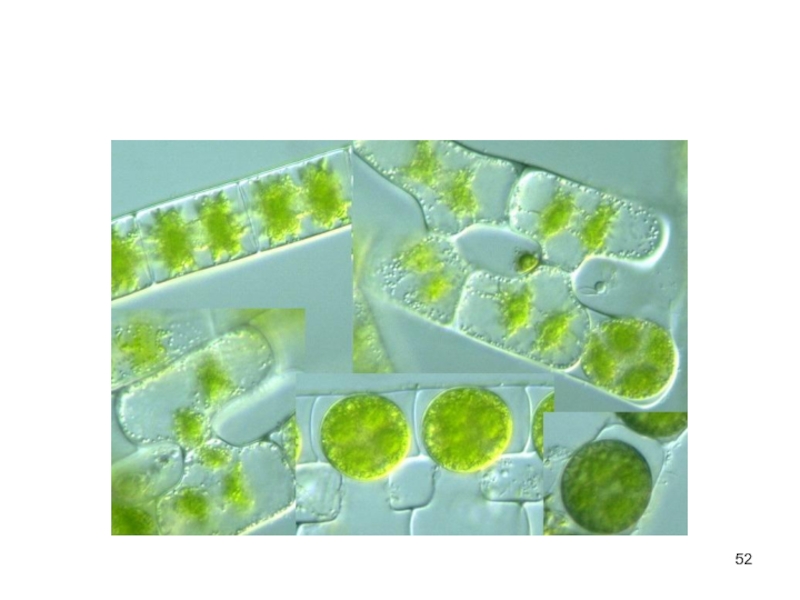
Слайд 50Порядок зигнемовые
нитчатые пресноводные водоросли.
неветвящиеся, состоят из одного ряда цилиндрических
клеток, одетых цельной оболочкой без пор и слизистым чехлом.

Слайд 51
Половой процесс — конъюгация.
Наиболее обычна лестничная конъюгация, которая происходит между
клетками двух нитей
Нити располагаются параллельно друг другу и вначале склеиваются слизью.
Затем противолежащие клетки - выросты
Оба протопласта вступают в контакт
образуют зиготу
Нити располагаются параллельно друг другу и вначале склеиваются слизью.
Затем противолежащие клетки - выросты
Оба протопласта вступают в контакт
образуют зиготу

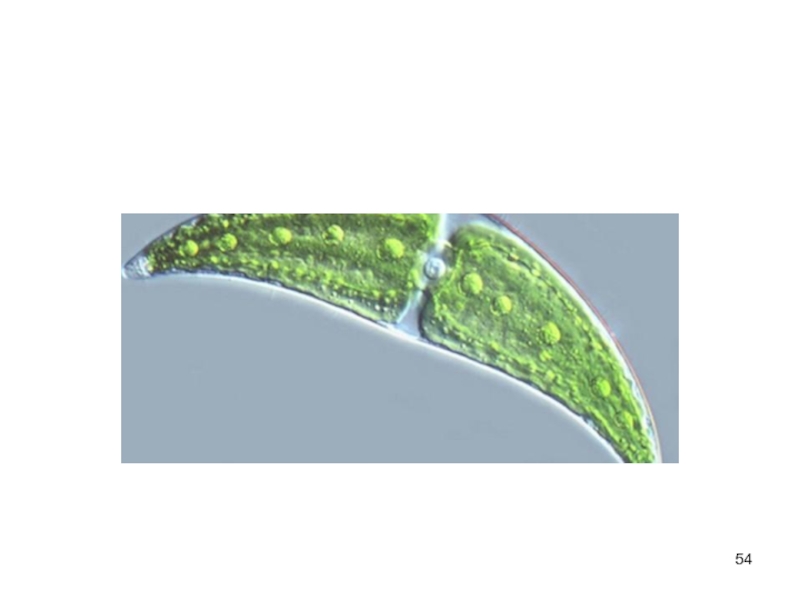
Слайд 53Порядок десмидиевые
одноклеточные, реже нитчатые водоросли.
Их клетки всегда состоят из
двух симметричных половинок — полуклеток.
род клостериум
Род космариум,
род эуаструм
род клостериум
Род космариум,
род эуаструм

Слайд 55КЛАСС ХАРОВЫЕ
представлен всего одним порядком
с тремя семействами, из которых два
объединяют ископаемые формы и одно - современные
Наиболее обычны роды хара и нителла.
От других зеленых водорослей они отличаются сложно устроенными многоклеточными половыми органами и расчленением нитчатого таллома .
Нитчатые талломы харовых, всегда вертикально стоящие, достигают высоты около 20-30 см (иногда и метра)
Мутовчатое ветвление - сходство с хвощами
Как неограниченно нарастающие оси («стебли»), так и боковые ветви ограниченного роста («листья») дифференцированы на узлы и междоузлия . Узлы представляю т собой клеточные пластинки. Они возникают из одной клетки в результате деления ее продольной перегородкой и затем отчленения продольными перегородками ряда (6—8) периферических клеток от двух центральных. От периферических клеток узла «стебля» отделяютс я верхушечные клетки боковых ветвей ограниченного роста — «листьев», благодаря этому расположенных всегда мутовками. Узлы чередуются с междо-узлиями, представленными у нителлы одной вытянутой (у самых крупных видов до 25 см) клеткой;
у хары клетка междоузлия снаружи одета специальной корой.
Наиболее обычны роды хара и нителла.
От других зеленых водорослей они отличаются сложно устроенными многоклеточными половыми органами и расчленением нитчатого таллома .
Нитчатые талломы харовых, всегда вертикально стоящие, достигают высоты около 20-30 см (иногда и метра)
Мутовчатое ветвление - сходство с хвощами
Как неограниченно нарастающие оси («стебли»), так и боковые ветви ограниченного роста («листья») дифференцированы на узлы и междоузлия . Узлы представляю т собой клеточные пластинки. Они возникают из одной клетки в результате деления ее продольной перегородкой и затем отчленения продольными перегородками ряда (6—8) периферических клеток от двух центральных. От периферических клеток узла «стебля» отделяютс я верхушечные клетки боковых ветвей ограниченного роста — «листьев», благодаря этому расположенных всегда мутовками. Узлы чередуются с междо-узлиями, представленными у нителлы одной вытянутой (у самых крупных видов до 25 см) клеткой;
у хары клетка междоузлия снаружи одета специальной корой.



Слайд 58КЛАСС ХАРОВЫЕ
Клетки одеты целлюлозной оболочкой, в наружны х слоях которой
отлагаетс я карбона т кальция .
В постенной цитоплазме находятся мелкие дисковидные хроматофоры, без пиреноидов.
Бесполого размножения у харовых нет.
Вегетативное размножение с помощью клубеньков, возникающих из нижних стеблевых узлов.
Половой процесс оогамный.
Половые органы имеют сложное строение.
Как правило, оогонии и антеридии развиваются в непосредственной близости друг от друга, однако есть и двудомные виды.
Оба вида половых органов формируются на вторичных боковых ветвях
В постенной цитоплазме находятся мелкие дисковидные хроматофоры, без пиреноидов.
Бесполого размножения у харовых нет.
Вегетативное размножение с помощью клубеньков, возникающих из нижних стеблевых узлов.
Половой процесс оогамный.
Половые органы имеют сложное строение.
Как правило, оогонии и антеридии развиваются в непосредственной близости друг от друга, однако есть и двудомные виды.
Оба вида половых органов формируются на вторичных боковых ветвях

Слайд 59Размножение харовых
Оогоний направлен вверх, антеридий — вниз
Плоские изогнутые клетки —
щитки, выпуклой стороной обращенные кнаружи. Щитки составляют стенку шаровидного антеридия.
Средние клетки значительно удлиняются в радиальном направлении, каждая образует клетку-рукоятку , которая таким образом отходит от центра щитка и несет на внутреннем конце шаровидную клетку — головку, развившуюся из самой внутренней клетки первоначального диагонального ряда.
Каждая клетка-головка дает начало шести вторичным головкам, на которых развиваются длинные нити, обычно дважды раздвоенные у основания.
Эти сперматогенные нити заполняю т полость антеридия плотным клубком . Каждая такая нить состоит из 100—200 дисковидных клеток, в которых образуется по одному спирально изогнутому двужгутиковому 1 сперматозоиду, сперматозоиды освобождаются при расхождении щитков антеридиальной стенки и ослизнении стенок клеток сперматогенных нитей.
Средние клетки значительно удлиняются в радиальном направлении, каждая образует клетку-рукоятку , которая таким образом отходит от центра щитка и несет на внутреннем конце шаровидную клетку — головку, развившуюся из самой внутренней клетки первоначального диагонального ряда.
Каждая клетка-головка дает начало шести вторичным головкам, на которых развиваются длинные нити, обычно дважды раздвоенные у основания.
Эти сперматогенные нити заполняю т полость антеридия плотным клубком . Каждая такая нить состоит из 100—200 дисковидных клеток, в которых образуется по одному спирально изогнутому двужгутиковому 1 сперматозоиду, сперматозоиды освобождаются при расхождении щитков антеридиальной стенки и ослизнении стенок клеток сперматогенных нитей.

Слайд 60
Клетка базального узла антеридия, которая развивается в оогоний, претерпевает два поперечных
деления с образованием ряда из трех клеток. Самая верхняя из них дает оогоний, при этом она сильно увеличивается, самая нижняя — одноклеточную ножку, а средняя образует узел, который делится на центральную и пять периферических клеток.
Последние вырастают в пять коровых нитей, которые уже на ранних стадиях окружают оогоний и вскоре отчленяют на своих концах одну (у хары) или две (у нителлы) клетки коронки . При дальнейшем развитии нити, не делясь, удлиняются и располагаются спирально.
Клетки коронки мелкие, вертикально стоящие. В оогонии формируется одна яйцеклетка. Под коронкой, клетки которой плотно соединены, нити слегка разъединяются , та к что образуетс я щель, через которую проникает сперматозоид. Верхушка оогониальной стенки ослизняется, и сперматозоид оплодотворяет яйцеклетку.
Вокруг оплодотворенной яйцеклетки выделяется целлюлозная оболочка
Последние вырастают в пять коровых нитей, которые уже на ранних стадиях окружают оогоний и вскоре отчленяют на своих концах одну (у хары) или две (у нителлы) клетки коронки . При дальнейшем развитии нити, не делясь, удлиняются и располагаются спирально.
Клетки коронки мелкие, вертикально стоящие. В оогонии формируется одна яйцеклетка. Под коронкой, клетки которой плотно соединены, нити слегка разъединяются , та к что образуетс я щель, через которую проникает сперматозоид. Верхушка оогониальной стенки ослизняется, и сперматозоид оплодотворяет яйцеклетку.
Вокруг оплодотворенной яйцеклетки выделяется целлюлозная оболочка


Слайд 62
Внутренние стенки нитей утолщаются , опробковевают, в них может откладыватьс я
кремнезем, а в полостях коровых нитей — обычно еще и известь. Таки м образом , вокруг ооспоры образуется очень твердая обвертка и ооспоры переходят в состояние покоя. При прорастании ооспоры копуляционное ядро редукционно делится, формируются четыре гаплоидных ядра и появляетс я перегородка, делящая ооспору на две неравны е части: верхняя клетка получает одно из четырех гаплоидных ядер, а три остальных ядра оказываютс я в нижней клетке, богатой запасным и веществами . Эти три ядра затем дегенерируют. Стенка ооспоры разрывается на верхушке и верхняя клетка продольно делится.
Получившиеся две клетки растут в противоположных направлениях , образуя первый ризоид и вертикальную нить — предросток, или протонему, на которой развивается затем нормальный побег.
По сложности строения вегетативных и половых органов, по нали-чию стадии протонемы, или предростка, при развитии зиготы харовы е
значительно отличаются от других зеленых ' водорослей, и ря д авторов
рассматриваю т их не только ка к отдельный класс, но даж е ка к само-стоятельный отдел. Однак о состав пигментов, наличи е крахмала ,
гаплоидный характе р (зиготическая редукция) , относительно простое
строение нителлы скорее свидетельствуют о том, что харовы е водоросли
представляю т собой лиш ь очень специализированну ю ветвь зеленых
водорослей.
Харовы е распространены" преимущественно в пресных водах, хотя
некоторые виды встречаются в солоноватых. Обычно они предпочитают
водоемы, где нет сильных движений воды и где имеется песчаный или
илистый грунт, в котором укореняются ризоиды.
Получившиеся две клетки растут в противоположных направлениях , образуя первый ризоид и вертикальную нить — предросток, или протонему, на которой развивается затем нормальный побег.
По сложности строения вегетативных и половых органов, по нали-чию стадии протонемы, или предростка, при развитии зиготы харовы е
значительно отличаются от других зеленых ' водорослей, и ря д авторов
рассматриваю т их не только ка к отдельный класс, но даж е ка к само-стоятельный отдел. Однак о состав пигментов, наличи е крахмала ,
гаплоидный характе р (зиготическая редукция) , относительно простое
строение нителлы скорее свидетельствуют о том, что харовы е водоросли
представляю т собой лиш ь очень специализированну ю ветвь зеленых
водорослей.
Харовы е распространены" преимущественно в пресных водах, хотя
некоторые виды встречаются в солоноватых. Обычно они предпочитают
водоемы, где нет сильных движений воды и где имеется песчаный или
илистый грунт, в котором укореняются ризоиды.






