- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
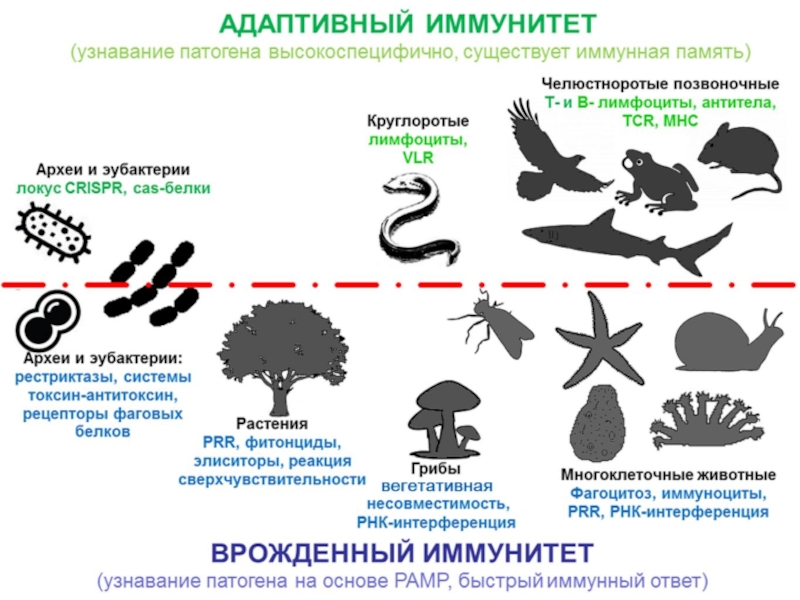
Защитные системы эукариот презентация
Содержание
- 1. Защитные системы эукариот
- 3. РНК-интерференция у эукариот Эндрю Файер и Крейг
- 4. Механизмы подавления экспрессии генов с помощью двуцепочечной
- 5. Гидом для pAgo является однонитевая ДНК, мишенью
- 6. Домены в составе pAgo pAgo есть у 30% архей и 10% эубактерий,
- 7. РНК-интерференция возникла за счёт объединения эубактериальных и архейных компонентов Dicer
- 8. Жизненный цикл амебозойного протиста Dictyostelium
- 9. Не все бактерии одинаково полезны (и питательны)
- 10. Механизмы передачи сигнала у слизевика
- 11. S-клетки (sentinel cells) отвечают за фагоцитоз бактерий в плодовом теле
- 12. Наличие «барьера Вейсмана» создает эволюционные предпосылки для его нарушения
- 13. Амебы – обманщики в химерных колониях не хотят образовывать ножку, и избегают ее зачатка
- 14. Плодовые тела диктиостелиума сильно структурированы и видоспецифичны, межвидовая агрегация отсутствует
- 15. Задача гифы – обеспечить рост апикальной клетки
- 16. Анастомозы, происходящие между соседними гифами внутри колонии,
- 17. При слиянии гиф двух или
- 18. Гетерокариоз – основа парасексуального процесса, т.е. рекомбинации без полового процесса.
- 19. Иногда мицелии принципиально не срастаются – происходит вегетативная несовместимость
- 20. Вегетативная несовместимость грибов
- 21. Барраж –гибель клеток в зоне контакта двух несовместимых мицелиев
- 22. Гетероаллельность по любому из Het генов вызывает
- 23. Вегетативная несовместимость основана на системе нескольких взаимодействующих генов
- 24. Аллели одного и того же гена несовместимости дают комплексный характер сочетаемости
- 25. Гибель клеток в зоне барража связана с избыточной аутофагией
- 26. Преимущества от вегетативной несовместимости грибов Защита от
- 27. О Р Ф Уровни иммунитета растения
- 28. ИММУНИТЕТ НА УРОВНЕ ЦЕЛОГО ОРГАНИЗМА (первая
- 29. Фитоиммунитет: распознавание элиситоров
- 30. Фитоиммунитет – распознавание PAMP (PTI) и эффекторных белков (ETI)
- 31. PAMP = неспецифические элиситоры
- 33. Важно детектировать не только грибы и бактерии, но и фитофагов
- 34. КАК РАСТЕНИЯ СПРАВЛЯЮТСЯ С ВИРУЛЕНТНЫМИ ПАРАЗИТАМИ (ТРЕТЬЯ
- 35. Белки с лейцин-богатыми повторами –основа и ETI, и PTI
- 36. R-белки – рецепторы с лейцин-богатыми повторами
- 37. «Игра в пятнашки» между патогеном и растением
- 38. Фитоантиципины – выделяются конститутивно (то есть постоянно)
- 39. Эффекторные белки фитоиммунитета Дефензины – низкомолекулярные
- 40. Реакция сверхчувствительности растений
- 41. Реакция сверхчувствительности и клеточная гибель
- 42. Механизмы реакции сверхчувствительности
- 43. РНК-интерференция против вирусов и вироидов
- 44. микроРНК как переключатель типов иммунного ответа у растений
- 45. Следующая лекция 5 марта – об иммунной системе дрозофилы

Слайд 3РНК-интерференция у эукариот
Эндрю Файер и Крейг Мелло, открыли явление в 1998

Слайд 4Механизмы подавления экспрессии генов с помощью двуцепочечной РНК (РНК интерференция)
цитоплазма
ядро
Dicer
расщепление
деградация мРНК
мРНК
подавление
трансляции мРНК
Dicer
siРНК
двунитевая РНК
мРНК
РНК шпилька
репрессия хроматина и
подавление транскрипции
гена в ядре
short interfering RNA (siRNA) – короткие РНК 20-25 нт.
РНКаза III
в комплекс с короткими РНК
входят белки Argonaute (AGO)
комплекс RISC
комплекс RIТS
 цитоплазмаядроDicerрасщепление идеградация мРНКмРНКподавление трансляции мРНКDicersiРНКдвунитевая")
Слайд 5Гидом для pAgo является однонитевая ДНК, мишенью – иногда РНК, иногда
Pyrococcus furiosus
Aquifex aeolicus
А вот у Rhodobacter sphaeroides pAgo есть, но вообще не работает




")

 отвечают за фагоцитоз бактерий в плодовом теле")


Слайд 14Плодовые тела диктиостелиума сильно структурированы и видоспецифичны, межвидовая агрегация отсутствует


Слайд 16Анастомозы, происходящие между соседними гифами внутри колонии, армируют ее делают более
Мицелиальные грибы – поскольку в мицелии делятся только апикальные клетки, но возможны анастамозы между гифами – тоже имеют барьер Вейсмана

Слайд 17 При слиянии гиф двух или более контактирующих колоний происходит
Гетерокариоз не тождественен диплоидности





Слайд 22Гетероаллельность по любому из Het генов вызывает острую реакцию гибели слившихся




Слайд 26Преимущества от вегетативной несовместимости грибов
Защита от эгоистичных паразитов, преодолевающих барьер Вейсмана
Защита
Защита территории и ресурсов от конкурентов с другим генотипом
Повышение генетического разнообразия популяций
Генетическая изоляция, приводящая к разделению экологических ниш и видообразованию


Слайд 28
ИММУНИТЕТ НА УРОВНЕ ЦЕЛОГО ОРГАНИЗМА
(первая линия обороны)
Цель – не допустить патогенных
тканям и клеткам
Факторы Животные Растения
Физические Плотное соединение Кутикулярный покров,
барьеры эпителиальных клеток полисахаридные стенки
Химические Жирные кислоты (кожа), Летучие антимикробные
барьеры лизоцим (слюна, слезы), вещества (фитонциды),
пепсин (кишечник), фенолы, терпены, депо-
дефензины (кишечник) нированные в листовых
волосках и мертвых кле-
тках покровных тканей
Микробиоло- Конкурентная микрофло- Микроорганизмы филло-
гические ба- ра эпителия, антибиоти- и ризопланы, их антиби-
рьеры ческие вещества отики, хитинолитические
ферменты и др.
Цель – не допустить патогенных организмов к чувствительным органам,")

 и эффекторных белков (ETI)")



Слайд 34КАК РАСТЕНИЯ СПРАВЛЯЮТСЯ С ВИРУЛЕНТНЫМИ ПАРАЗИТАМИ (ТРЕТЬЯ ЛИНИЯ ОБОРОНЫ)
У растений возникла
Например, фумонизин является фитотоксином для ряда растений и элиситором защитных реакций для Arabidopsis; бактериальный токсин коронатин в разных растениях может выступать как токсин, фитогормон и элиситор. Специфические элиситоры – эффекторы, которые паразит выделяет в зараженное растение для подавления его защитного потенциала и улучшения условий питания (супрессоры). Поскольку структура многих таких соединений может меняться без потери жизнеспособности, возможны мутации, изменяющие домены, которые узнаются специфическими клеточными рецепторами растения. Однако, часто это сопровождается снижением патогенности (“сильные гены устойчивости” Вандерпланка, “цена вирулентности” Леонарда).
 У растений возникла система ген-на ген, в")



Слайд 38Фитоантиципины – выделяются конститутивно (то есть постоянно)
Фитоалексины –выделяются в ответ на
Низкомолекулярные соединения летят на помощь
Фитоалексины –выделяются в ответ на появление фитофага или патогенаНизкомолекулярные")
Слайд 39Эффекторные белки фитоиммунитета
Дефензины – низкомолекулярные (45-54 аминокислот) белки, богатые цистеином. Продуцируются
Растительные дефензины делятся на: 1) морфогенные (вызывают морфологические изменения у восприимчивых видов грибов)
2) неморфогенные (ингибируют рост без морфозов), ингибируют альфа-амилазу.
Связываются с рецепторами на мембранах грибов, усиливают потерю К+ и поступление Са2+.
Тионины – богатые серой белки, накапливающиеся в семенах и проростках растений. Взаимодействуют с мембранными фосфолипидами и вызывают образование пор в мембранах.
RIP-белки – подавляют синтез белка на чужеродных рибосомах на стадии элонгации
 белки, богатые цистеином. Продуцируются млекопитающими, грибами, насекомыми и")











