- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Закономерности наследственности и изменчивости презентация
Содержание
- 1. Закономерности наследственности и изменчивости
- 2. Законы Менделя 1 закон. Закон единообразия или
- 3. Общие закономерности расщепления при скрещивании полигетерозигот Полигетерозиготы
- 4. Возвратное скрещивание Скрещивание гибридных форм с родительскими
- 6. Рецепроктное или обратное скрещивание Данный вид скрещивания
- 7. Сцепленное наследование Законы Моргана (хромосомная теории наследования):
- 8. Расстояние между локусами Морган предложил измерять расстояние
- 9. Полигенное наследование В большинстве случаев один признак
- 10. Основные типы взаимодействия генов : комплементарность, эпистаз
- 11. 2. Э п и с т а
- 12. 3. Полимерия Полимерия – это такой
- 13. Межаллельные взаимодействия Неполное доминирование – транскрипция рецессивного
- 14. Наследование признаков, сцепленных с полом, у
- 15. Несмотря на то, что женские особи
- 16. ГЕНЕТИКИ ПОПУЛЯЦИЙ С точки зрения генетики,
- 17. Характеристика идеальной популяции Идеальная популяция – это
- 18. Условия существования идеальной популяции 1. Рост численности
- 19. Реальные популяции в большей или меньшей
- 20. Определения Панмиксия (свободное скрещивание) означает, что
- 21. Партеногенез – это девиантная форма полового процесса,
- 22. Генетическая структура популяций Каждая популяция
- 23. Аллелофонд . Аллелофонд популяции – это совокупность
- 24. Генофонд. Термин генофонд употребляется в разных значениях.
- 25. Закон Харди–Вайнберга – основной закон популяционной генетики
- 26. Действие закона Харди-Вайнберга при полном доминировании Известно,
- 27. Практическое значение закона Харди–Вайнберга 1. В
- 28. 2. В селекции – позволяет выявить генетический
- 29. Генетические параметры популяции При описании популяций
- 30. Различные популяционно-генетические процессы по-разному влияют на
- 31. ИЗМЕНЧИВОСТЬ
- 32. Изменчивость -- это возникновение индивидуальных различий.
- 33. Классификация форм изменчивость Фенотипическая
- 34. Модификационная изменчивость Модификационная изменчивость не вызывает
- 35. Свойства модификаций: 1) ненаследуемость;
- 36. Генотипическая изменчивость Генотипическая изменчивость подразделяется на
- 37. Мутации классифицируют по уровню их возникновения.
- 38. Хромосомные мутации связаны с изменением структуры
- 39. Генные мутации (точечные) – истинные мутации
- 40. При выведении новых сортов растений и
- 41. Свойства мутаций. 1. Мутации возникают
- 42. Комбинативная изменчивость Комбинативная наследственная изменчивость возникает
- 43. Соотносительная (коррелятивная) изменчивость Возникает в результате свойства
- 44. Закон гомологических рядов В селекции важное значение
- 45. Помимо вышерассмотренных различают изменчивость:
- 46. Методы генетики Биохимический – позволяет по продукту
- 47. Типы наследования признаков - Аутосомно-доминантный тип наследования:
- 48. - Аутосомно-рецессивный тип наследования: а.
- 49. Наследование сцепленное с Х хромосомой, если
- 50. Наследование сцепленное с Х хромосомой, если
- 51. Наследование сцепленное с У хромосомой:
- 52. МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ В хранении, передаче
- 53. Спаривание оснований в ДНК Первым свидетельством
- 54. Структура ДНК Спаривание оснований, проиллюстрированное на
- 56. Молекулярные модели ДНК и тРНК
- 58. Молекулы РНК не могут образовывать двойную
- 59. Рибонуклеиновые кислоты РНК принимают участие
- 60. Транспортные РНК ( tRNAPhe )
- 62. Репликация Для передачи дочерним клеткам генетической
- 64. В настоящее время процесс репликации у
- 65. Транскрипция Для того чтобы хранящаяся в
- 66. Структура β-глобинового гена В качестве
- 68. Как уже упоминалось, РНК-полимераза II связывается
- 69. Созревание РНК Большинство меток организма содержит
- 70. Сплайсинг Сплайсинг РНК катализируется комплексами
- 72. Генетический код, активация аминокислот Большая часть
- 74. Активация аминокислот Для каждой из
- 75. Характеристика генетического кода
- 76. Классификация генов у эукариот 1. Акцепторные –
- 77. Классификация генов у эукариот 1) домашнего хозяйства
- 78. ОПЕРОН

Слайд 2Законы Менделя
1 закон. Закон единообразия или закон доминирования.
У гибридов 1-го поколения
2 закон. Закон расщипления
Рецессивный признак не исчезает, он сохраняется у гибридов первого поколения в неизменном виде и проявляется у гибридов второго поколения в соотношении 3:1 – признак детерминирован
3 закон. Закон независимого наследования
Признаки наследуются независимо друг от друга (каждый по отдельности признак у гибридов 2-го поколения дает расщепление 3:1), образует все возможные сочетания признаков в соотношении 9:3:3:1 – наследственность дискретна.
Наследственность детерминирована и дискретна

Слайд 3Общие закономерности расщепления при скрещивании полигетерозигот
Полигетерозиготы – это организмы, гетерозиготные по

Слайд 4Возвратное скрещивание
Скрещивание гибридных форм с родительскими формами (гомозиготами по доминантному или
Анализирующее скрещивание позволяет выявить гетерозиготность фенотипически доминантной форма, т.е. определить генотип фенотипически доминантной формы. Для этого анализируемую форму скрещивают с гомозиготой по рецессиву. Анализирующее скрещивание один из видов возвратного скрешивания.
АА x аа Аа – в Fв расщепления нет
Аа x аа 1 Аа : 1 аа – расщепление по генотипу
в Fв расщепление по фенотипу 1[А-] : 1 [аа]


Слайд 6Рецепроктное или обратное скрещивание
Данный вид скрещивания позволяет определить связанно ли наследование
OА-ХOаа OааХOА-

Слайд 7Сцепленное наследование
Законы Моргана (хромосомная теории наследования):
Гены наследуются совместно образуя группу сцепления
Локусы генов располагаются в хромосоме линейно
Хромосомы способны конъюгировать и обмениваться гомологичными участками (кроссинговер), образуя рекомбинанты. Сила сцепления между локусами зависит от расстояния между ними.
- абсолютное сцепление- гены расположены рядом
- относительное сцепление- возможны рекомбинации(до 46%)
- практически независимое наследование - количество рекомбинантов около 50%
Сила сцепления обратно пропорциональна расстоянию между локусами.
Вероятность кроссинговера прямо пропорционально расстоянию
между локусами и обратно пропорциональна силе сцепления
:Гены наследуются совместно образуя группу сцепления или хромосому Локусы генов")
Слайд 8Расстояние между локусами
Морган предложил измерять расстояние между локусами в процентах рекомбинантных
1% = 1 М
Пример: в потомстве из 180 мух 45 имели новые сочитания 2-х признаков, т.е. отличались сочитанием признаков от родительских форм – были рекомбинантами
расстояние между локусами 2 анализируемых генов находится так: 180 - 100%
45 - х % х = 25%

Слайд 9Полигенное наследование
В большинстве случаев один признак контролируется множеством разных (неаллельных) генов.
Олигогены – это главные гены, которые в наибольшей степени ответственны за формирование признака. Обеспечивают дискретную (прерывистую) изменчивость качественных признаков.
Супрессоры – гены, подавляющие действие других генов.
Бустеры – гены, усиливающие действие других генов.
Модификаторы – гены, оказывающие незначительное влияние на формирование признака, уточняющие действие других генов.
Полигены – множество генов, оказывающих сходное действие. Обеспечивают непрерывную изменчивость количественных
 генов. Такое наследование признаков, при")
Слайд 10Основные типы взаимодействия генов : комплементарность, эпистаз и полимерия.
1. Комплементарность.
Комплементарность – это такой тип полигенного наследования, при котором неаллельные гены взаимно дополняют друг друга. В простейшем случае рассматривается два гена, каждый из которых представлен двумя аллелями, причем, аллель А полностью доминирует над аллелем а, а аллель В полностью доминирует над аллелем b. Если два комплементарных гена не сцеплены между собой, то во втором поколении возможны следующие расщепления по фенотипу: 9:3:3:1, 9:6:1, 9:7

Слайд 112. Э п и с т а з
Эпистаз – это такой
Различают рецессивный и доминантный эпистаз. Обычно рассматривают случаи полного доминирования для каждого неаллельного гена, причем эпистатический и гипостатический гены не сцеплены между собой. Тогда при рецессивном эпистазе в F2 наблюдаются расщепления 9 : 3 : 4 или 9 : 7, а при доминантном эпистазе – 13:3 или 12:3:1.

Слайд 123. Полимерия
Полимерия – это такой тип полигенного наследования, при котором признак
Некумулятивная полимерия. Для качественных признаков характерна некумулятивная полимерия с полным доминированием и расщеплением 15 : 1 (в случае действия двух пар аллелей)
Кумулятивная полимерия. Для количественных признаков характерна кумулятивная полимерия с неполным доминированием и расщеплением 1 : 4 : 6 : 4 : 1 (в случае действия двух пар аллелей).

Слайд 13Межаллельные взаимодействия
Неполное доминирование – транскрипция рецессивного гена не подавляется действием доминантного.
Кодоминирование – совместное действие генов с одинаковой фенотипической экспрессией. В результате гибриды имеют новый признак.
Множественный аллелизм – существование в популяции 3 и более аллелей одного гена (серии генов), возникших в результате многократных мутаций в одном и том же локусе.
Аллели в серии проявляют или соотносительное домини-рование, или кодоминирование. У
Сверхдоминирование – в гетерозиготном состоянии признак проявляется с большей фенотипической силой чем у гомозигот. Такое явление (гетерозисная сила) проявляется при скрещивании (гетерозисе) особей инбридных линий. Гетерозисная сила объясняется возрастанием общей жизнестойкости и жизнеспособности гибридов в следствии перевода в гетерозиготное состояние большинства рецессивных «вредных» генов.

Слайд 14
Наследование признаков, сцепленных с полом, у млекопитающих
У млекопитающих самки являются гомогаметным
Пол у млекопитающих определяется наличием Y–хромосомы. В состав Y-хромосомы входит H–Y–антиген, который запускает экспрессию генов, определяющих формирование основных мужских половых признаков. В большинстве случаев у самцов млекопитающих Y–хромосома практически не несет генов. Однако некоторые аномальные признаки самцов (например, волосатые уши у мужчин, ихтиодермия или развитие у них перепонок между вторым и третьим пальцами ног) обусловлены изменениями в Y–хромосоме. Подобные аномалии стопроцентно наследуются по мужской линии от отца ко всем его сыновьям и далее. Такие признаки называются голандрическими.

Слайд 15
Несмотря на то, что женские особи млекопитающих имеют две Х-хромосомы, а

Слайд 16ГЕНЕТИКИ ПОПУЛЯЦИЙ
С точки зрения генетики,
популяция – это
Таким образом, с точки зрения генетики, популяция представляет собой множество особей, объединенных достаточно высокой степенью родства.
В рамках генетического подхода выделяется представление об идеальной популяции.

Слайд 17Характеристика идеальной популяции
Идеальная популяция – это абстрактное понятие, которое широко используется

Слайд 18Условия существования идеальной популяции
1. Рост численности популяции ничем не ограничен
3. Особи со всеми видами генотипа обладают одинаковой жизнеспособностью. Естественный отбор не оказывает заметного влияния на частоту большинства аллелей
4. Особи с разными генотипами имеют возможность скрещиваться с равной вероятностью
5. Скрещивание особей равновероятно
6. Скорость прямых мутаций равна скорости обратных (нет мутационных изменений генотипа и генофонда)
7. Отсутствуют межаллельные и межгенные взаимодействия

Слайд 19
Реальные популяции в большей или меньшей степени отличаются от идеальной. Одним
По способу воспроизведения различают следующие типы популяций:
амфимиктические – основным способом размножения является нормальное половое воспроизведение;
амфимиктические панмиктические – при формировании брачных пар наблюдается панмиксия (свободное скрещивание);
амфимиктические инбредные – при формирование брачных пар наблюдается близкородственное скрещивание (инбридинг, инцухт, инцест); крайним случаем близкородственного скрещивания является самооплодотворение;
апомиктические – наблюдаются различные отклонения от нормального полового процесса, например, апомиксис, партеногенез, гиногенез, андрогенез; наблюдается у агамных (бесполых) форм;
клональные – при отсутствии полового процесса и размножении только вегетативным путем или с помощью спор бесполого размножения (например, конидий); частным случаем клонирования является полиэмбриония – развитие нескольких зародышей из одной зиготы:
комбинированные – например, клонально-амфимиктические при метагенезе у кишечнополостных (чередовании бесполого и полового размножения) и гетерогонии (чередовании партеногенетического и амфимиктического поколений у червей, некоторых членистоногих и низших хордовых).

Слайд 20Определения
Панмиксия (свободное скрещивание) означает, что на формирование брачных пар не влияет
Инбридинг – близкородственное скрещивание у животных; инцухт – близкородственное скрещивание у растений; инцест (кровосмешение) – при которых не происходит объединения двух клеток. Обычно этот термин используют по отношению к растениям. При апомиксисе новый организм может развиваться из неоплодотворенной яйцеклетки (см. партеногенез), а также из какой-либо другой специализированной клетки зародышевого мешка (например, из клеток–антипод или синергид), реже – непосредственно из клеток нуцеллуса или покровов семязачатка. Близкородственное скрещивание у человека.
Апомиксис – это множество форм образования зародышей. Примеры растений–апомиктов: ястребинки, одуванчики, манжетки.
 означает, что на формирование брачных пар не влияет генотип или возраст особей,")
Слайд 21Партеногенез – это девиантная форма полового процесса, при которой новый организм
Гиногенез – это девиантная форма полового процесса, при которой мужские гаметы служат для стимуляции развития нового организма из яйцеклетки, но оплодотворения не происходит, и мужское ядро (пронуклеус) погибает. В этом случае у дочернего организма сохраняются только материнские хромосомы. Гиногенез встречается у гибридов рыб, земноводных, а также в бессамцовых популяциях.
Андрогенез – это девиантная форма полового процесса, при которой происходит оплодотворение, но затем женское ядро (пронуклеус) погибает, а мужское ядро замещает его в качестве ядра зиготы. В этом случае у дочернего организма сохраняются только отцовские хромосомы. Андрогенез обычно наблюдается в лабораторных условиях.
Агамные формы – организмы, у которых отсутствует нормальный половой процесс
Партеногенез – это девиантная форма полового процесса, при которой новый организм развивается из неоплодотворенной яйцеклетки без участия мужских гамет. Различают нередуцированный партеногенез с развитием зародыша из диплоидной клетки и редуцированный партеногенез с развитием зародыша из гаплоидной яйцеклетки. Как правило, партеногенез чередуется с нормальным половым размножением (при цикломорфозе у коловраток, дафний, тлей).
Гиногенез – это девиантная форма полового процесса, при которой мужские гаметы служат для стимуляции развития нового организма из яйцеклетки, но оплодотворения не происходит, и мужское ядро (пронуклеус) погибает. В этом случае у дочернего организма сохраняются только материнские хромосомы. Гиногенез встречается у гибридов рыб, земноводных, а также в бессамцовых популяциях.
Андрогенез – это девиантная форма полового процесса, при которой происходит оплодотворение, но затем женское ядро (пронуклеус) погибает, а мужское ядро замещает его в качестве ядра зиготы. В этом случае у дочернего организма сохраняются только отцовские хромосомы. Андрогенез обычно наблюдается в лабораторных условиях.
Агамные формы – организмы, у которых отсутствует нормальный половой процесс

Слайд 22Генетическая структура популяций
Каждая популяция обладает собственной генетической структурой. Генетическая структура

Слайд 23Аллелофонд
. Аллелофонд популяции – это совокупность аллелей в популяции. Если рассматриваются
Популяции, в которых структура аллелофонда остается относительно постоянной в течение длительного времени, называются стационарными.
Если рассматриваются три аллеля одного гена: а1, а2,, а3, то структура аллелофонда описывается уравнением: p а1 + q а2 + r а3 = 1. В этом уравнении символами p, q, r обозначаются соответствующие частоты аллелей.

Слайд 24Генофонд.
Термин генофонд употребляется в разных значениях. Основоположник учения о генофонде и
При изучении природных популяций часто приходится сталкиваться с полным доминированием: фенотипы гомозигот АА и гетерозигот Аа неразличимы. Кроме того, в природе широко распространено полигенное определение признаков, причем типы взаимодействия неаллельных генов (комплементарность, эпистаз, полимерия) не всегда известны. Поэтому на практике часто изучают не генофонд, а фенофонд популяций, то есть соотношение фенотипов. В настоящее время развивается раздел генетики популяций, который называется фенетика популяций.

Слайд 25Закон Харди–Вайнберга – основной закон популяционной генетики
Структура генофонда в панмиктической стационарной
Если известны относительные частоты аллелей p и q и общая численность популяции Nобщ, то можно рассчитать ожидаемую, или расчетную абсолютную частоту (то есть численность особей) каждого генотипа. Для этого каждый член уравнения нужно умножить на Nобщ:
p2 AA · Nобщ + 2·p·q Aa · Nобщ + q2 aa · Nобщ = Nобщ
В данном уравнении:
p2 AA · Nобщ – ожидаемая абсолютная частота (численность) доминантных гомозигот АА
2·p·q Aa · Nобщ – ожидаемая абсолютная частота (численность) гетерозигот Аа
q2 aa · Nобщ – ожидаемая абсолютная частота (численность) рецессивных гомозигот аа

Слайд 26Действие закона Харди-Вайнберга при полном доминировании
Известно, что черная окраска шерсти у
Предположим, что в одной из городских популяций кошек на о. Сахалин из 100 просмотренных животных полную или частичную черную окраску имели 36 животных.
Прямой расчет структуры аллелофонда популяции в этом случае невозможен из-за полного доминирования: гомозиготы АА и гетерозиготы Аа фенотипически неразличимы. Согласно уравнению Харди-Вайнберга частота черных кошек составляет q2 аа. Тогда можно рассчитать частоты аллелей:
q2aa = 36/100 = 0,36; qa = 0,36 –1/2 =0,6; pA = 1 – 0,6 = 0,4
Таким образом, структура аллелофонда данной популяции описывается соотношением: р А + q a = 0,4 + 0,6 = 1. Частота рецессивного аллеля оказалась выше, чем частота доминантного.
Рассчитаем частоты генотипов:
р2 АА = 0,42 = 0,16; 2 pq Аа = 2 х 0,4 х 0,6 = 0,48; q2aa = 0,62 = 0,36
Рассчитаем число особей с разными генотипами:
n AA= 0,16 х100=16 особей;
n Aa= 0,48х100=48 особей;
п aa = 0,36х 100=36 особей.

Слайд 27Практическое значение закона Харди–Вайнберга
1. В здравоохранении – позволяет оценить популяционный
Пример 1. Известно, что альбинизм – это аутосомно-рецессивное заболевание. Установлено, что в большинстве европейских популяций частота рождения детей-альбиносов составляет 1 на 20 тысяч новорожденных. Следовательно,
q2aa = 1/20000 = 0,00005; qa = 0,00005–1/2 = 0,007; pA = 1 – 0,007 = 0,993 ≈ 1
Поскольку для редких заболеваний рА ≈ 1, то частоту гетерозиготных носителей можно рассчитать по формуле 2·q. В данной популяции частота гетерозиготных носителей аллеля альбинизма составляет 2 q Аа = 2 х 0,007 = 0,014, или примерно каждый семидесятый член популяции.
Пример 2. Пусть в одной из популяций у 1% населения выявлен рецессивный аллель, который не встречается в гомозиготном состоянии (можно предположить, что в гомозиготном состоянии этот аллель летален). Тогда 2 q Аа = 0,01, следовательно, qa = 0,01:2 = 0,005. Зная частоту рецессивного аллеля, можно установить частоту гибели зародышей–гомозигот: q2aa = 0,0052 = 0,000025 (25 на миллион, или 1 на 40 тысяч).

Слайд 282. В селекции – позволяет выявить генетический потенциал исходного материала (природных
3. В экологии – позволяет выявить влияние самых разнообразных факторов на популяции. Дело в том, что, оставаясь фенотипически однородной, популяция может существенно изменять свою генетическую структуру под воздействием ионизирующего излучения, электромагнитных полей и других неблагоприятных факторов. По отклонениям фактических частот генотипов от расчетных величин можно установить эффект действия экологических факторов. (При этом нужно строго соблюдать принцип единственного различия. Пусть изучается влияние содержания тяжелых металлов в почве на генетическую структуру популяций определенного вида растений. Тогда должны сравниваться две популяции, обитающие в крайне сходных условиях. Единственное различие в условиях обитания должно заключаться в различном содержании определенного металла в почве).
2. В селекции – позволяет выявить генетический потенциал исходного материала (природных популяций, а также сортов и пород народной селекции), поскольку разные сорта и породы характеризуются собственными аллелофондами, которые могут быть рассчитаны с помощью закона Харди-Вайнберга. Если в исходном материале выявлена высокая частота требуемого аллеля, то можно ожидать быстрого получения желаемого результата при отборе. Если же частота требуемого аллеля низка, то нужно или искать другой исходный материал, или вводить требуемый аллель из других популяций (сортов и пород).
3. В экологии – позволяет выявить влияние самых разнообразных факторов на популяции. Дело в том, что, оставаясь фенотипически однородной, популяция может существенно изменять свою генетическую структуру под воздействием ионизирующего излучения, электромагнитных полей и других неблагоприятных факторов. По отклонениям фактических частот генотипов от расчетных величин можно установить эффект действия экологических факторов. (При этом нужно строго соблюдать принцип единственного различия. Пусть изучается влияние содержания тяжелых металлов в почве на генетическую структуру популяций определенного вида растений. Тогда должны сравниваться две популяции, обитающие в крайне сходных условиях. Единственное различие в условиях обитания должно заключаться в различном содержании определенного металла в почве).

Слайд 29Генетические параметры популяции
При описании популяций или их сравнении между собой
Полиморфизм. Популяция называется полиморфной по данному локусу, если в ней встречается два или большее число аллелей. Если локус представлен единственным аллелем, говорят о мономорфизме. Исследуя много локусов, можно определить среди них долю полиморфных, т.е. оценить степень полиморфизма, которая является показателем генетического разнообразия популяции.
Гетерозиготность. Важной генетической характеристикой популяции является гетерозиготность – частота гетерозиготных особей в популяции. Она отражает также генетическое разнообразие.
Коэффициент инбридинга. С помощью этого коэффициента оценивают распространенность близкородственных скрещиваний в популяции.
Ассоциация генов. Частоты аллелей разных генов могут зависеть друг от друга, что характеризуется коэффициентами ассоциации.
Генетические расстояния. Разные популяции отличаются друг от друга по частоте аллелей. Для количественной оценки этих различий предложены показатели, называемые генетическими расстояниями.

Слайд 30
Различные популяционно-генетические процессы по-разному влияют на эти параметры: инбридинг приводит к
Прилагая эти подходы и методы к исследованию популяций человека, животных, растений и микроорганизмов, можно решить многие проблемы эволюции, экологии, медицины, селекции и др. Рассмотрим несколько примеров, демонстрирующих связь популяционной генетики с другими науками.


Слайд 32
Изменчивость -- это возникновение индивидуальных различий. На основе изменчивости организмов появляется

Слайд 33
Классификация форм изменчивость
Фенотипическая (ненаследуемая - паратипическая)
Модификационная
Фенокопии – фенотипическое копирование мутаций при
Случайная – не обусловлена воздействием типичных факторов, обычно носит травматический характер
Генотипическая (наследуемая)
Комбинативная
Мутационная: генные (точечные), хромосомные, геномные.
Соотносительная (коррелятивная)
МодификационнаяФенокопии – фенотипическое копирование мутаций при критическом изменении действующего фактора")
Слайд 34Модификационная изменчивость
Модификационная изменчивость не вызывает изменений генотипа, она связана с реакцией
Модификационная изменчивость обычно колеблется в определенных пределах. Степень варьирования признака у организма, то есть пределы модификационной изменчивости, называется нормой реакции.
Широкая норма реакции свойственна таким признакам, как удои молока, размеры листьев, окраска у некоторых бабочек;
узкая норма реакции -- жирности молока, яйценоскости у кур, интенсивности окраски венчиков у цветков и другое.
Фенотип формируется в результате взаимодействий генотипа и факторов среды. Фенотипические признаки не передаются от родителей потомкам, наследуется лишь норма реакции, то есть характер реагирования на изменение окружающих условий. У гетерозиготных организмов при изменении условий среды можно вызвать различные проявления данного признака

Слайд 35
Свойства модификаций:
1) ненаследуемость;
2) групповой характер изменений;
3) соотнесение изменений
4) обусловленность пределов изменчивости генотипом.
 ненаследуемость; 2) групповой характер изменений; 3) соотнесение изменений действию определенного фактора среды;")
Слайд 36Генотипическая изменчивость
Генотипическая изменчивость подразделяется на мутационную и комбинативную.
Мутациями называются скачкообразные
Классификация мутаций.
Мутации можно объединять, в группы -классифицировать по характеру проявления, по месту или, по уровню их возникновения.
Мутации по характеру проявления бывают доминантными и рецессивными. Мутации нередко понижают жизнеспособность или плодовитость. Мутации, резко снижающие жизнеспособность, частично или полностью останавливающие развитие, называют полулетальными а несовместимые с жизнью - летальными.
Мутации подразделяют по месту их возникновения. Мутация, возникшая в половых клетках, не влияет на признаки данного организма, а проявляется только в следующем поколении. Такие мутации называют генеративными. Если изменяются гены в соматических клетках, такие мутации проявляются у данного организма и не передаются потомству при половом размножении. Но при бесполом размножении, если организм развивается из клетки или группы клеток, имеющих изменившийся -- мутировавший -- ген, мутации могут передаваться потомству. Такие мутации называют соматическими.

Слайд 37Мутации классифицируют по уровню их возникновения.
Геномные - изменение кариотипа (изменение
Полиплоидия -- увеличение числа хромосом, кратное гаплоидному набору. В соответствии с этим у растений различают триплоиды (Зп), тетраплоиды (4п) и т.д. В растениеводстве известно более 500 полиплоидов (сахарная свекла, виноград, гречиха, мята, редис, лук и др.). Все они выделяются большой вегетативной массой и имеют большую хозяйственную ценность.Полиплоиды получают в результате воздействия на растения температуры, ионизирующей радиации, химических веществ (колхицин), которые разрушают веретено деления клетки. У таких растений гаметы диплоидны, а при слиянии с гаплоидными половыми клетками партнера в зиготе возникает триплоидный набор хромосом (2п + п = Зп). Такие триплоиды не образуют семян, они бесплодны, но высокоурожайны. Четные полиплоиды образуют семена.
Гетероплоидия - изменение числа хромосом, не кратное гаплоидному набору. При этом набор хромосом в клетке может быть увеличен на одну, две, три хромосомы (2п + 1; 2п + 2; 2п + 3) или уменьшен на одну хромосому (2л-1). Например, у человека с снндромом Дауна оказывается одна лишняя хромосома по 21-й паре и кариотип такого человека составляет 47 хромосом. У людей с синдромом Шерешевского --Тернера (2п-1) отсутствует одна Х-хромосома и в кариотипе остается 45 хромосом. Эти и другие подобные отклонения числовых отношений в кариотипе человека сопровождаются расстройством здоровья, нарушением психики и телосложения, снижением жизнеспособности и др.
Мутации классифицируют по уровню их возникновения.
Геномные - изменение кариотипа (изменение числа хромосом).
Полиплоидия -- увеличение числа хромосом, кратное гаплоидному набору. В соответствии с этим у растений различают триплоиды (Зп), тетраплоиды (4п) и т.д. В растениеводстве известно более 500 полиплоидов (сахарная свекла, виноград, гречиха, мята, редис, лук и др.). Все они выделяются большой вегетативной массой и имеют большую хозяйственную ценность.Полиплоиды получают в результате воздействия на растения температуры, ионизирующей радиации, химических веществ (колхицин), которые разрушают веретено деления клетки. У таких растений гаметы диплоидны, а при слиянии с гаплоидными половыми клетками партнера в зиготе возникает триплоидный набор хромосом (2п + п = Зп). Такие триплоиды не образуют семян, они бесплодны, но высокоурожайны. Четные полиплоиды образуют семена.
Гетероплоидия - изменение числа хромосом, не кратное гаплоидному набору. При этом набор хромосом в клетке может быть увеличен на одну, две, три хромосомы (2п + 1; 2п + 2; 2п + 3) или уменьшен на одну хромосому (2л-1). Например, у человека с снндромом Дауна оказывается одна лишняя хромосома по 21-й паре и кариотип такого человека составляет 47 хромосом. У людей с синдромом Шерешевского --Тернера (2п-1) отсутствует одна Х-хромосома и в кариотипе остается 45 хромосом. Эти и другие подобные отклонения числовых отношений в кариотипе человека сопровождаются расстройством здоровья, нарушением психики и телосложения, снижением жизнеспособности и др.
. Полиплоидия --")
Слайд 38
Хромосомные мутации связаны с изменением структуры хромосом.
Существуют следующие виды перестроек
делеции - отрыв различных участков хромосомы,
дупликации - удвоение отдельных фрагментов,
инверсии - поворот участка хромосомы на 180°
транслокации - присоединение отдельного участка хромосомы к другой хромосоме.
транспозиция - перенос участка хромосомы в другой участок той же хромосомы.
Подобное изменение влечет за собой нарушение генетического гомеостаза, функции генов в хромосоме и наследственных свойств организма, а иногда и его гибель.

Слайд 39Генные мутации (точечные) –
истинные мутации
Затрагивают структуру самого гена и влекут
Существуют следующие виды генных мутаций:
делеции - потеря участков генов,
дупликации - удвоение отдельных фрагментов гена,
инверсии - поворот участка гена на 180°
транслокации – встраивания в ген участка не гомологичного гена.
транспозиция - перенос участка гена в другой участок того же гена.
Потеря или добавление участка гена вызывает изменение рамки считывание и нарушает последовательность нуклеотидов, а следовательно и изменение качества триплетов. Данный вид генных мутаций наиболее разрушителен.
 – истинные мутацииЗатрагивают структуру самого гена и влекут за собой изменение свойств")
Слайд 40
При выведении новых сортов растений и штаммов микроорганизмов используют индуцированные мутации,
В природе мутации носят спонтанный характер
Мутации могут быть:
ядерные
внеядерные (цитоплазматические) – возникают в клеточных структурах имеющих собственное ДНК (митохондрии, пластиды, плазмиды и т.д.), передаются по материнской линии, т.к. при оплодотворении цитоплазматические структуры сперматозоида в формировании зиготы не участвуют.
Способность к мутированию - одно из свойств гена. Каждая отдельная мутация вызывается какой-то причиной, но в большинстве случаев эти причины неизвестны. Мутации связаны с изменениями во внешней среде. Это убедительно доказывается тем, что путем воздействия внешними факторами удается резко повысить их число.

Слайд 41Свойства мутаций.
1. Мутации возникают внезапно, скачкообразно.
2. Мутации наследственны, то
3. Мутации ненаправленные - мутировать может любой локус, вызывая изменения как незначительных, так и жизненно важных признаков.
4. Одни и те же мутации могут возникать повторно.
5. По своему проявлению мутации могут быть полезными и вредными, доминантными и рецессивными.

Слайд 42Комбинативная изменчивость
Комбинативная наследственная изменчивость возникает в результате
2) как следствие независимого расхождения хромосом при мейозе;
3) случайного сочетания хромосом при скрещивании.
Изменчивость может быть обусловлена не только мутациями, но и сочетаниями отдельных генов и хромосом, новая комбинация которых при размножении приводит к изменению определенных признаков и свойств организма. Такой тип изменчивости называют комбинативной наследственной изменчивостью. Новые комбинации генов возникают:
1) при кроссинговере, во время профазы первого мейотического деления;
2) во время независимого расхождения гомологичных хромосом в анафазе первого мейотического деления;
3) во время независимого расхождения дочерних хромосом в анафазе второго мейотического деления
4) при слиянии разных половых клеток.
Сочетание в зиготе рекомбинированных генов может привести к объединению признаков разных пород и сортов.
Комбинативная изменчивость обеспечивает появление особей с наиболее благоприятным сочетанием признаков.
 обмена гомологичными участками гомологичных хромосом")
Слайд 43Соотносительная (коррелятивная)
изменчивость
Возникает в результате свойства генов влиять на формирование не одного,
Обеспечивает постоянство взаимосвязанных признаков, целостность организма как системы
Пример:
Длинноногие животные имеют длинную шею. У столовых сортов свеклы согласованно изменяется окраска корнеплода, черешков и жилок листа
 изменчивостьВозникает в результате свойства генов влиять на формирование не одного, а двух и")
Слайд 44Закон гомологических рядов
В селекции важное значение имеет закон гомологических рядов наследственной
Он гласит:
Внутри разных видов и родов, генетически близких (т. е. имеющих единое происхождение), наблюдаются сходные ряды наследственной изменчивости.
Такой характер изменчивости выявлен у многих злаков (рис, пшеница, овес, просо и др.), у которых сходно варьируют окраска и консистенция зерна, холодостойкость и иные качества.
Зная характер наследственных изменений у одних сортов, можно предвидеть сходные изменения у родственных видов и, воздействуя на них мутагенами, вызывать у них подобные полезные изменения, что значительно облегчает получение хозяйственно ценных форм.
Известны многие примеры гомологической изменчивости и у человека; например, альбинизм (дефект синтеза клетками красящего вещества) обнаружен у европейцев, негров и индейцев; среди млекопитающих -- у грызунов, хищных, приматов; малорослые темнокожие люди - пигмеи встречаются в тропических лесах экваториальной Африки, на Филиппинских островах и в джунглях полуострова Малакки; некоторые наследственные дефекты и уродства, присущие человеку, отмечены и у животных. Таких животных используют в качестве модели для изучения аналогичных дефектов у человека. Например, катаракта глаза бывает у мыши, крысы, собаки, лошади; гемофилия - у мыши и кошки, диабет - у крысы; врожденная глухота - у морской свинки, мыши, собаки; заячья губа - у мыши, собаки, свиньи и т. д. Эти наследственные дефекты -- убедительное подтверждение закона гомологических рядов наследственной изменчивости Н. И. Вавилова.

Слайд 45Помимо вышерассмотренных различают изменчивость:
индивидуальную и групповую;
качественную и количественную;
независимую изменчивость разных признаков и коррелятивную (соотносительную);
направленную (определенную, по Ч.Дарвину) и ненаправленную (неопределенную, по Ч.Дарвину);
адаптивную (приспособительную) и неадаптивную.
При решении общих проблем биологии и особенно эволюции наиболее существенно подразделение изменчивости, с одной стороны, на наследственную и ненаследственную, а с другой - на индивидуальную и групповую. Все категории изменчивости могут встречаться в наследственной и ненаследственной, групповой и индивидуальной изменчивости.
 и непрерывную; качественную")
Слайд 46Методы генетики
Биохимический – позволяет по продукту (белку) определить нарушения (мутации) в
Кариотипирование – позволяет обнаружить геномные и хромосомные мутации
Генеологический – позволяет выявить характер наследования признака (аутосомно-доминантный, аутосомно- рецессивный, сцепленный с Х-хромосомой доминантный или рецессивный, сцепленный с У-хромосомой)
Близнецовый –позволяет определить роль наследственности и среды в формирование фенотипа
Популяционно-статистический – определить соотношение генотипов в популяции, концентрацию различных аллелей
Гибридологический (предложен Г.Менделем) – позволяет проанализировать генотипы особей, выявить формы взаимодействия и взаимовлияния генов и аллелей.
Картирование – позволяет установить структуру хромосом и место локализации генов
 определить нарушения (мутации) в структуре генаКариотипирование – позволяет")
Слайд 47Типы наследования признаков
- Аутосомно-доминантный тип наследования:
а. При достаточном числе потомков

Слайд 48
- Аутосомно-рецессивный тип наследования:
а. Признак может передаваться через поколение даже

Слайд 49
Наследование сцепленное с Х хромосомой, если ген, контролирующий проявления признака -
а. Мужчины наследуют чаще, чем женщины б. Наследуют такой признак девочки только от отца в. В браках, где оба супруга здоровы, могут родиться дети, имеющие его, при этом он наследуется 50% сыновей и 100% здоровых дочерей г. Прослеживается чередование больных мужчин в поколениях: где их больше, где - меньше

Слайд 50
Наследование сцепленное с Х хромосомой, если ген, контролирующий проявления признака, -
а. Мужчины наследуют реже, чем женщины б. Если признак только у супруги, то наследуют его все дети (мать гомозиготная), или половина детей (мать гетерозиготная) в. Если только у супруга, то наследуют все лица женского пола

Слайд 51
Наследование сцепленное с У хромосомой:
а. Страдают только сыновья,
б.

Слайд 52МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ
В хранении, передаче и преобразовании генетической информации центральное место

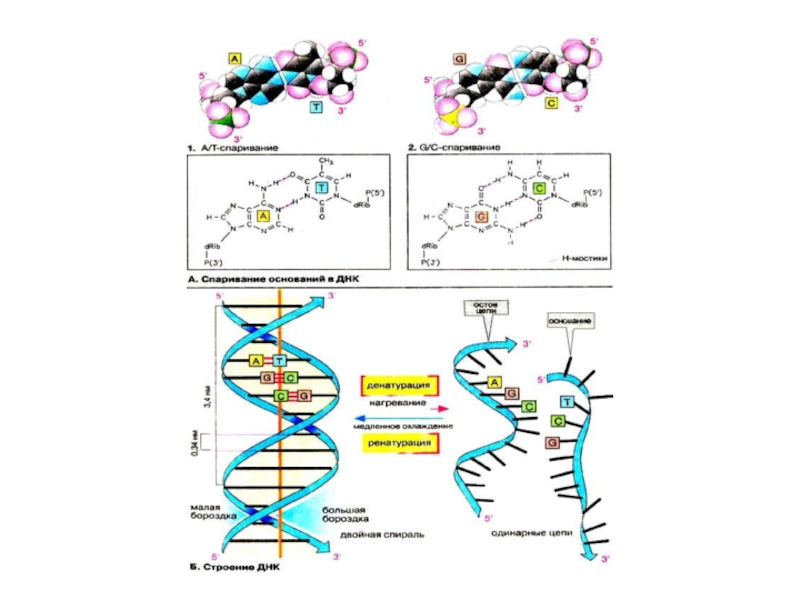
Слайд 53Спаривание оснований в ДНК
Первым свидетельством существования таких структур пocлyжил тот факт,
Комплементарность А и T, соответственно G и С, становится понятной, если рассмотреть возможные водородные мостики между основаниями. В качестве доноров (см. с. 14) выступают аминогруппы (аденина, цитозина, гуанина) и NН-группы гетероциклов (тимина и гуанина). Возможными акцепторами являются карбонильные группы (тимина, цитозина, гуанина) и атомы азота гетероциклов. Пара A-T может образовывать два, а пара G-C даже три линейных и поэтому особенно устойчивых мостика. Урацил, содержащийся в РНК вместо тимина, ведет себя при спаривании основании подобно тимину.

Слайд 54Структура ДНК
Спаривание оснований, проиллюстрированное на схеме А, охватывает в молекуле ДНК
Преобладающая в клетке конформация ДНК (так называемая В-ДНК) представлена схематически на схеме 1, в виде вандерваальсовой модели. На схеме 1 дезоксирибозофосфатный остов изображен в виде ленты. Основания (здесь указаны в виде полос) расположены внутри двойной спирали. Следовательно, эта область ДНК неполярна.
Напротив, внешняя сторона молекулы полярна и заряжена отрицательно за счет углеводных остатков и фосфатных групп остова. Цепи ДНК на протяжении всего тяжа образуют два желоба, которые носят названия «малая бороздка» и «большая бороздка».
Так как обе цепи связаны только нековалентными взаимодействиями, двойная спираль при нагревании или инкубации в щелочном растворе легко распадается на отдельные цепи (денатурирует). При медленном охлаждении ранее неупорядоченные отдельные цепи благодаря спариванию оснований вновь образуют двойную спираль (молекула ренатурирует). Процессы де- и ренатурации играют важную роль в генной инженерии .
В функциональном отношении две цепи ДНК не эквивалентны. Кодирующей цепью (матричной, смысловой) является та из них, которая считывается в процессе транскрипции . Именно эта цепь служит матрицей для PHK. Некодирующая цепь (антисмысловая) по последовательности подобна РНК (при условии замены T на U). Общепринято давать структуру гена в виде последовательности некодирующей цепи ДНК в направлении 5'→3'. Если прочитать кодоны в этом направлении, то с помощью генетического кода можно воспроизвести аминокислотную последовательность белка в принятом порядке, от N- к С-концу.

Слайд 56Молекулярные модели ДНК и тРНК
На схеме представлены вандерваальсовые модели двух небольших
При исследовании синтетических молекул ДНК было показано, что ДНК может принимать различные конформации. Наиболее часто встречается приведенная здесь В-форма ДНК. Молекула состоит из двух антипараллельных полидезоксинуклеотидных цепей, которые закручены в правую двойную спираль. Остов этого тяжа образован остатками дезоксирибозы и фосфатными группировками, соединенными фосфодиэфирными связями. Остов одной цепи выделен темно-синим цветом, другой — синим, а основания обеих цепей — голубым. Ароматические кольца отстоят друг от друга на 0,34 нм и почти перпендикулярны оси спирали. Каждое основание повернуто по отношению к предыдущему на 35о. На каждый виток двойной спирали (360о) приходится примерно 10 пар оснований, ход спирали — 3,4 нм. Между остовами двух отдельных цепей имеются две широкие бороздки. Большая бороздка видна на модели снизу и сверху, малая бороздка — в центре. ДНК-ассоциированные белки взаимодействуют с наиболее доступными основаниями в области большой бороздки. Пространственные соотношения между остовом и основаниями более наглядно видны на модели одинарной цепи (справа). На всех моделях аденин окрашен в желтый цвет, а комплементарный ему тимин — в оранжевый. На виде сверху (в центре; одна цепь выше другой) основания просматриваются наиболее полно.

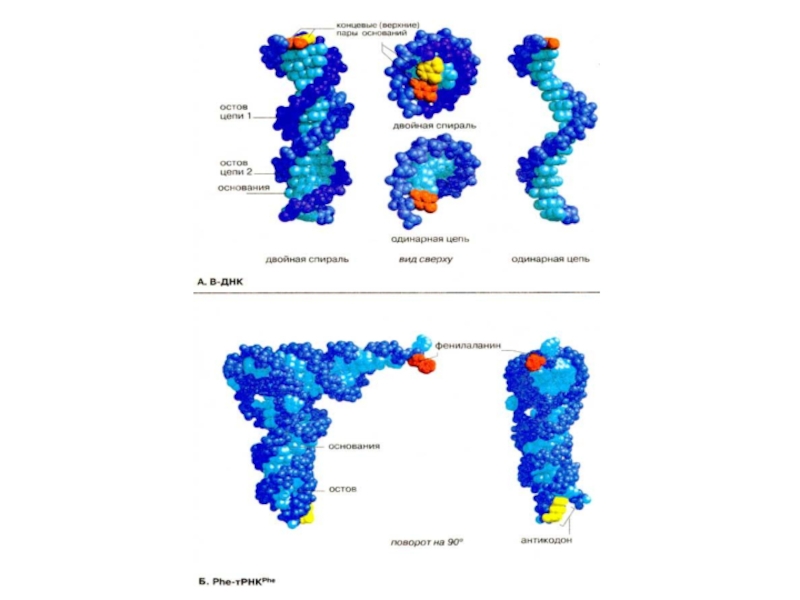
Слайд 58
Молекулы РНК не могут образовывать двойную спираль и поэтому не столь
На модели видно, что тРНК свернута в компактную клинообразную структуру. Как и в ДНК, большинство основании находятся внутри молекулы, в то время как полярный остов — на поверхности. Исключение составляют три основания антикодона (окрашены в желтый цвет), который должен взаимодействовать с мРНК. Большинство оснований принимает участие в межмолекулярном спаривании, некоторые из образованных пар не соответствуют общим принципам спаривания оснований, обычно соблюдаемым в ДНК (A + U, G + С). Рибонуклеиновые кислоты [PHK(RNA)] представляют собой полимеры из нуклеозидфосфатных звеньев, соединенных фосфодиэфирной связью. В качестве азотистых оснований в РНК присутствуют урацил, цитозин, аденин и тимин. В РНК можно также встретить множество необычных и модифицированных азотистых оснований.

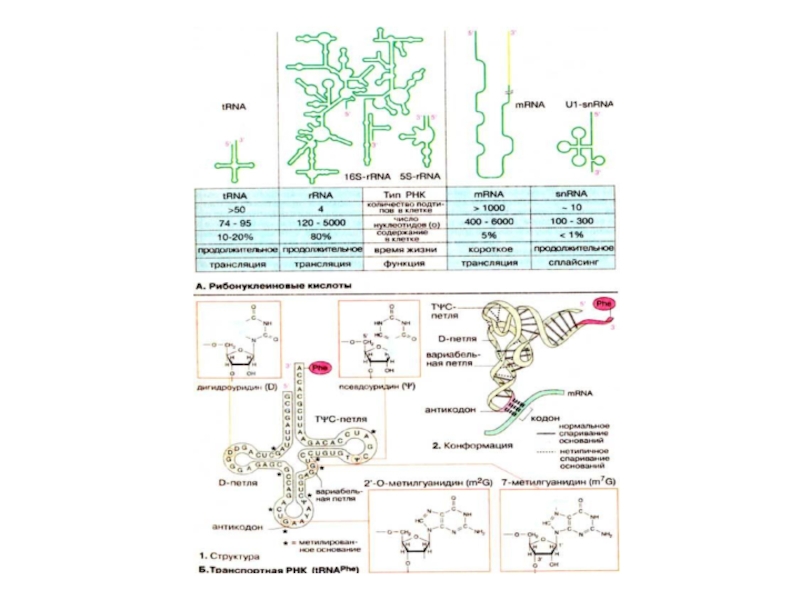
Слайд 59Рибонуклеиновые кислоты
РНК принимают участие во всех стадиях процесса генной экспрессии и
В отличие от ДНК, РНК не образуют двойных спиралей, но содержат короткие участки со спаренными основаниями. Это приводит к образованию субструктур, которые при двумерном изображении напоминают «шпильки» и петли, образующие фигуру типа «кленового листа». В таких структурах двухцепочечные участки соединены петлями. Множество фрагментов, в которых чередуются структуры типа шпилька—петля, содержится в высокомолекулярных РНК, таких, например, как рибосомная 16S-рРНК (16S-rRNA) (в центре). Кроме того, эти фрагменты образуют трехмерные структуры; следовательно, РНК подобно белкам имеют четвертичную структуру. До настоящего времени установлена четвертичная структура небольших PHK, прежде всего тРНК (tRNA). Из иллюстраций, приведенных на схеме Б очевидно, что трехмерная укладка структуры типа «кленовый лист» окончательно не установлена.
PHK клетки существенно различаются по размерам, строению и продолжительности существования. Преобладающую часть представляют рибосомные РНК [рРНК (rRNA)], которые в различных формах составляют структурный и функциональные части рибосом . Рибосомные РНК синтезируются в ядре в процессе транскрипции на ДНК, там же подвергаются процессингу и ассоциируют с рибосомными белками, образуя рибосому . Приведенная на схеме А бактериальная 16S-рРНК, включающая 1542 нуклеотида, является компонентом малой рибосомной субчастицы, в то время как небольшая 5S-рРНК (из 120 нуклеотидов) входит в состав большой субчастицы.
Матричная РНК [мРНК (mRNA)] переносит генетическую информацию из клеточного ядра в цитоплазму. Ее транскрипты также сильно модифицируются в ядре (созревание мPHK) Так как мРНК считывается на рибосоме кодон за кодоном она не должна складываться в стабильную третичную структуру. Спариванию оснований препятствуют белки, ассоциированные с мРНК. Из-за различного объема информации, которую могут нести мРНК, РНК этого типа сильно варьируют по размерам. Для мРНК характерно короткое время жизни, так как они быстро распадаются после трансляции. В сплайсинге предшественников мРНК принимают участие малые ядерные РНК [мяРНК (snRNA, от англ. small nuclear RNA)]. Они ассоциированы c рядом белков, образуя «сплайсомы». . Из-за различного объема информации, которую могут нести мРНК, РНК этого типа сильно варьируют по размерам. Для мРНК характерно короткое время жизни, так как они быстро распадаются после трансляции. В сплайсинге предшественников мРНК принимают участие малые ядерные РНК [мяРНК (snRNA, от англ. small nuclear RNA)]. Они ассоциированы c рядом белков, образуя «сплайсомы». . Из-за различного объема информации, которую могут нести мРНК, РНК этого типа сильно варьируют по размерам. Для мРНК характерно короткое время жизни, так как они быстро распадаются после трансляции. В сплайсинге предшественников мРНК принимают участие малые ядерные РНК [мяРНК (snRNA, от англ. small nuclear RNA)]. Они ассоциированы c рядом белков, образуя «сплайсомы».

Слайд 60Транспортные РНК ( tRNAPhe )
Транспортные РНК [тРНК (tRNA)] участвуют в процессе
Последовательность оснований и третичная структура фенилаланинспецифичной тРНК (tRNAPhe) из дрожжей являются типичными для всех тРНК. В молекуле этой тРНК содержится довольно много минорных и модифицированных оснований (1, выделены темно-зеленым цветом). К ним относятся псевдоуридин (ψ), дигидроуридин (D), тимидин (T), встречающийся обычно в ДНК, а также множество метилированных нуклеотидов, таких, например, как 7-метилгуанидин (m7G) и входящий в состав антикодона 2'-О-метилгуанидин (m2G). Конформацию молекулы стабилизируют многочисленные пары оснований, часть из которых не соответствуют общим принципам спаривания
 Транспортные РНК [тРНК (tRNA)] участвуют в процессе трансляции в качестве")
Слайд 62Репликация
Для передачи дочерним клеткам генетической информации в процессе репликации ДНК (DNA)
 должна быть создана")

Слайд 64
В настоящее время процесс репликации у прокариот достаточно изучен, в то
Поскольку матричная цепь всегда читается в направлении 3'→5', только одна из цепей может считываться непрерывно (розовая/фиолетовая; 2). Другая цепь (голубого цвета) считывается в направлении, противоположном движению репликативной вилки. В результате на матрице вначале синтезируются короткие фрагменты новой цепи ДНК (зеленый/оранжевый), так называемые фрагменты Оказаки (OF), названные так по имени их первооткрывателя. Каждый фрагмент начинается с короткой РНК-затравки (праймера, зеленого цвета), необходимой для функционирования ДНК-полимеразы. Праймер синтезируется специальной РНК-полимеразой («праймаза», на схеме не показана), ДНК-полимераза III достраивает этот праймер до фрагмента ДНК длиной 1000-2000 дезоксинуклеотидных звеньев (оранжевого цвета). Синтез этого фрагмента далее прерывается, и новый синтез начинается со следующего РНК-праймера. Индивидуальные фрагменты Оказаки первоначально не связаны друг с другом и все еще имеют РНК на 5'-концах (3). На некотором расстоянии от репликативной вилки ДНК-полимераза I начинает замещать РНК-праймер последовательностью ДНК. В завершение остающиеся одноцепочечные разрывы репарируются ДНК-лигазой. В образованной таким образом двойной спирали ДНК только одна из цепей синтезирована заново. Поэтому говорят, что репликация ДНК происходит по полуконсервативному механизму

Слайд 65Транскрипция
Для того чтобы хранящаяся в ДНК информация могла быть использована, ее
Транскрипция осуществляется ДНК-зависимыми РНК-полимеразами. Они действуют подобно ДНК-полимеразам, за исключением того, что включают во вновь синтезируемую цепь РНК (RNA) рибонуклеотиды вместо дезоксирибонуклеотидов и не нуждаются в праймерах. Эукариотические клетки обычно содержат по крайней мере три различных типа РНК-полимераз, РНК-полимераза I катализирует синтез РНК с коэффициентом седиментации 45S, которая служит предшественником трех различных рибосомных РНК. РНК-полимеразы II синтезируют гяРНК (hnRNA), которые служат предшественниками мРНК (mRNA) и мяРНК (snRNA). Наконец, РНК-полимераза III транскрибирует гены, которые кодируют тРНК (tRNA), 5S- рРНК (rRNA) и некоторые мяРНК. Эти РНК служат предшественниками функциональных РНК, которые образуются в процессе созревания РНК .
")
Слайд 66Структура β-глобинового гена
В качестве примера организации небольшого эукариотического гена представлена схема
У многих генов разделение на экзоны и интроны еще более выражено. Так, общая длина гена дигидрофолатредуктазы составляет выше 30 тыс.п.о. Информация об аминокислотной последовательности распределена по 6 экзонам, которые в сумме составляют только ~6 тыс.п.о.

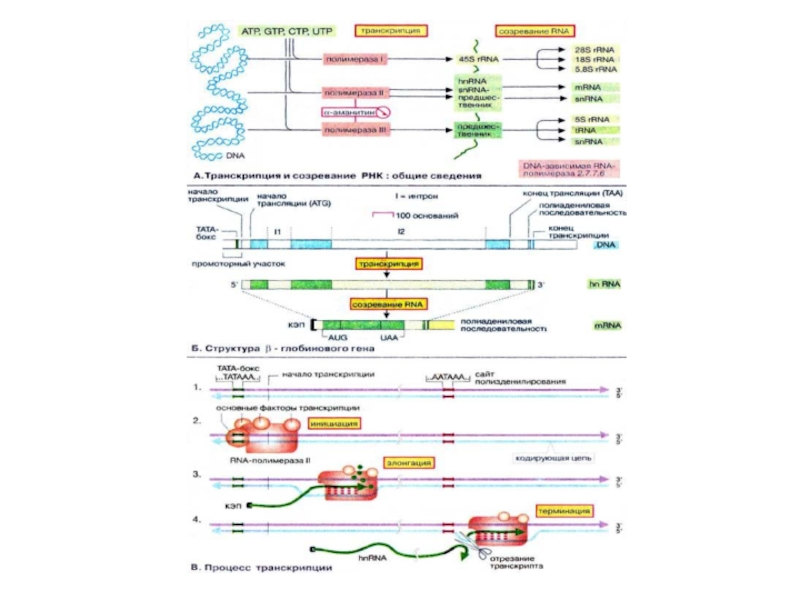
Слайд 68
Как уже упоминалось, РНК-полимераза II связывается с 3'-концом промоторного участка. Последовательность,
После инициации синтеза (2), РНК-полимераза движется в направлении 3'→5' матричной цепи. В процессе инициации фермент разделяет короткий участок двойной спирали ДНК на две отдельные цепочки. Нуклеозидтрифосфаты связываются комплементарно на кодирующей цепочке ДНК водородными связями и шаг за шагом присоединяются к растущей молекуле РНК (3). Вскоре после начала элонгации 5'-конец транскрипта защищается «кэпом» (от англ. cap). Как только транскрипция доходит до сайта полиаденилирования (обычно это последовательность ...ААТААА...), транскрипт отщепляется (4). После этого полимераза прекращает транскрипцию и диссоциирует от ДНК.

Слайд 69Созревание РНК
Большинство меток организма содержит полный набор генов, но обычно из
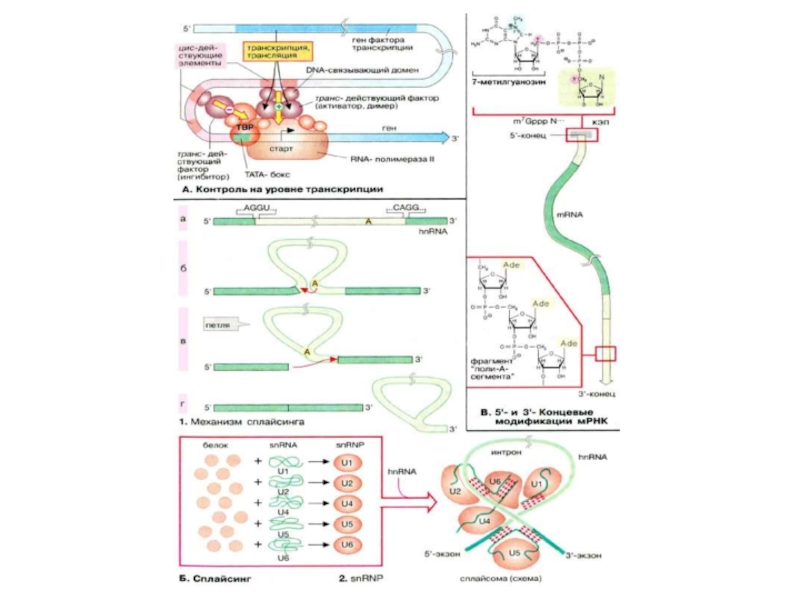
Контроль на уровне транскрипции
Порядок транскрибирования генов определяется регуляторной системой, которая носит название системы регуляции транскрипции. Контроль транскрипции осуществляется структурами двух типов. Большинство генов содержат в своем промоторном участке несколько коротких сегментов ДНК (DNA) (регуляторные элементы, цис-действующие элементы), с которыми могут связываться факторы транскрипции. Регуляторные элементы, стимулирующие транскрипцию связанных с ними генов, называются энхансерами (усилителями, от англ. enhancer). Белки, подавляющие транскрипцию. — сайленсерами (успокоителями, от англ. silencer). Факторы транскрипции — это белки, т. е. продукты других, независимых генов. Поэтому их называют опосредованно действующими факторами. Для процесса транскрипции генов требуются не только РНК-полимераза, но и другие белки, называемые основными факторами транскрипции. Установлено, что у эукариот таким фактором является ТАТА-связывающий белок (ТСБ, англ. ТАТА-Вох Binding Protein, TBP), который взаимодействует с основным регуляторным элементом. ТАТА-боксом, присутствующим в большинстве генов. С этим комплексом затем связываются другие основные факторы транскрипции и РНК-полимеразы. Дополнительные факторы могут влиять на инициацию транскрипции, связываясь с другими регуляторными элементами. Отсюда они взаимодействуют с основным транскрипционным комплексом, либо активируя, либо ингибируя его. Такие факторы активируют, например, комплексы стероидных гормонов с рецепторами. По завершении транскрипции из гяРНК вырезаются интроны, содержащие некодирующие последовательности- процесс процессинга

Слайд 70Сплайсинг
Сплайсинг РНК катализируется комплексами белков с РНК, известными как «малые ядерные
Во время сплайсинга комплексы из гяРНК и мяРНП образуют сплайсому. Полагают, что мяРНК в сплайсоме образуют канонические пары друг с другом и с гяРНК и таким образом фиксируют и ориентируют их реакционные группы. Собственно катализ обусловлен РНК-составляющей сплайсомы Такие каталитические РНК носят название рибозимов.
В. 5'- и 3'-Концевые модификации мРНК
У эукариот после завершения собственно транскрипции 5'-конец растущей молекулы РНК блокируется структурой, которая называется кэп (от англ. cap). В случае мРНК кэп состоит из 7'-метил-ГТФ и защищает РНК от гидролиза 5'-экзонуклеазами. В конце транскрипции к 3'-концу присоединяется полиадениловая последовательность, которая может включать до 200 звеньев АМФ (AMP). Только после этого созревшая мРНК (mRNA) покидает ядро.
.")
Слайд 72Генетический код, активация аминокислот
Большая часть генетической информации, содержащейся в ДНК, кодирует
Поскольку в биосинтезе участвуют 20 аминокислот, называемых протеиногенными, «язык» нуклеиновых кислот должен содержать по крайней мере 20 слов (кодонов) Однако в аминокислотном «алфавите» имеется только четыре «буквы» (А, Г, Ц и У или Т [или в англ. транскрипции: A, G, С и U или Т*]), так что для получения 20 различных слов каждое должно состоять по крайней мере из трех букв. Кодоны действительно включают три азотистых основания (триплет нуклеотидов). На схеме 1 представлен стандартный код ДНК (последовательность триплетов в некодирующей цепи), изображенный в виде круга. Схема читается от центра наружу, так что, например, триплет CAT кодирует аминокислоту гистидин. ДНК-кодоны идентичны таковым в мРНК (mRNA), за исключением того, что в мРНК вместо урацила (U), характерного для ДНК, стоит тимин (Т).
В триплетном генетическом коде для 20 аминокислот потенциально существует 43 = 64 кодона. Таким образом, большинство аминокислот записывается несколькими кодонами, т. е. генетический код является вырожденным. Кроме того, имеются три триплета, которые обозначают конец транскрипции (стоп-кодоны). Еще один специальный кодон, стартовый (инициирующий) кодон, маркирует начало трансляции. Генетический код, показанный на рисунке, является почти универсальным. Этому стандарту не полностью соответствуют только митохондрии и некоторые микроорганизмы.

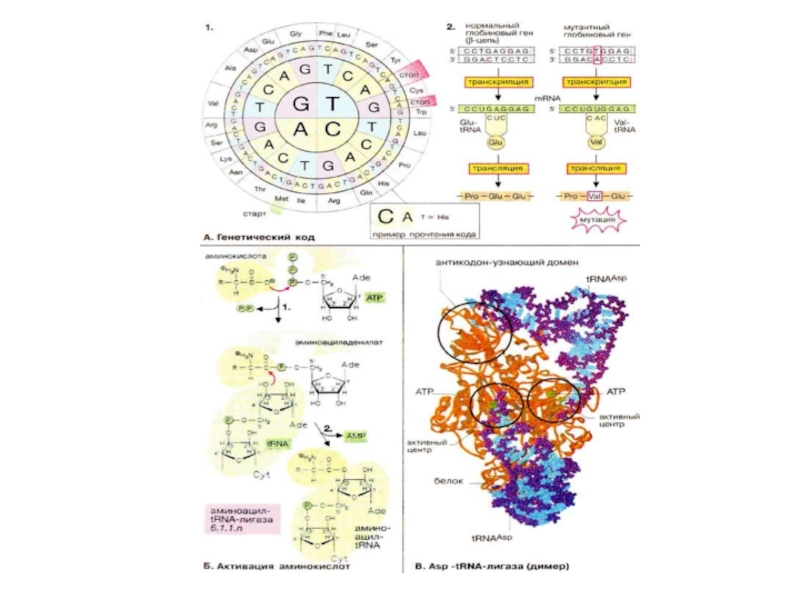
Слайд 74Активация аминокислот
Для каждой из 20 аминокислот имеется соответствующая аминоацил-тРНК-лигаза, которая в
Точность трансляции зависит, прежде всего, от субстратной специфичности аминоацил-тРНК-лигаз. Корректирующий механизм активного центра лигазы обеспечивает немедленное удаление ошибочно присоединенных аминокислотных остатков. В среднем встречается только одна ошибка на 1300 аминокислотных остатков — поразительно высокая точность «работы», если представить, насколько близки структуры некоторых аминокислот.
Процесс активации аминокислот представлен на примере лигазы, специфичной для аспарагиновой кислоты. Молекулы фермента (окрашены в оранжевый цвет) связаны между собой в димер, причем каждая субъединица ассоциирована с одной молекулой тРНК (окрашены в голубой цвет). В активном центре присутствует остаток АТФ (окрашен в зеленый цвет), связанный с 3'-концом тРНК Другой домен белка (слева вверху) отвечает за «узнавание» антикодона тРНК.


Слайд 76Классификация генов у эукариот
1. Акцепторные – регулируют деятельность структурных генов
2. Структурные
1) по структуре организации:
а) уникальные гены. Представлены в единственном числе и не имеют копий. Если в них происходят мутации. То они сразу проявляются фенотипически
б) умеренно повторяющиеся, имеет до 70 копий, кодируют синтез специфических белков (тубулина, миозина)
в) высокоповторяющиеся более 70 копий, входит в состав сателлитной ДНК.
2) псевдогены – в ходе мутаций утратили свою информативность, но так как они нейтральны, то не исчезают.
3) спейсеры –это гены связывающие акцепторные и структурные гены между собой.

Слайд 77Классификация генов у эукариот
1) домашнего хозяйства
2) роскошного синтеза
3) гамиозисные гены –активируются
4) гены стресса – в норме неактивны, функционируютв экстремальной ситуации
5) прото-онкогены – в каждой клетке до 70 онкогенов
 домашнего хозяйства2) роскошного синтеза3) гамиозисные гены –активируются в период эмбриогенеза4) гены")







