Проспективные потенции области зародыша: множество возможных вариантов судьбы области (динамис по Аристотелю)
ЗАКОН ДРИША: проспективные потенции области шире ее проспективного значения. Это эмбриональные регуляции при изменении условий развития, а в неизменных условиях – изменчивость нормального развития
ЭКВИПОТЕНЦИАЛЬНОСТЬ ПО ДРИШУ: каждая часть развивающейся системы равнозначна системе как целому. Это возможно только при равенстве начальных условий развития (Dyctiostelium)
 значение области зародыша: судьба данной области при нормальном ходе развития (энтелехия по")
 и многоклеточной (плазмодий,")







: модель, показывающая как формируется нормальное распределение: «детерминированный хаос»Для возникновения изменчивости достаточно")


")
 эксцесс означает положительную (стабилизирующую), (-)")







, развитие которых считается детерминированным, случайный выбор смещается из эмбриогенеза в оогенез")
 и геометрический (геометрия движения клеток)")


 не просто существует, а активно поддерживается за счет замкнутой")


, или же оказаться в медианной зоне")
, образующей арку над желточной")
")
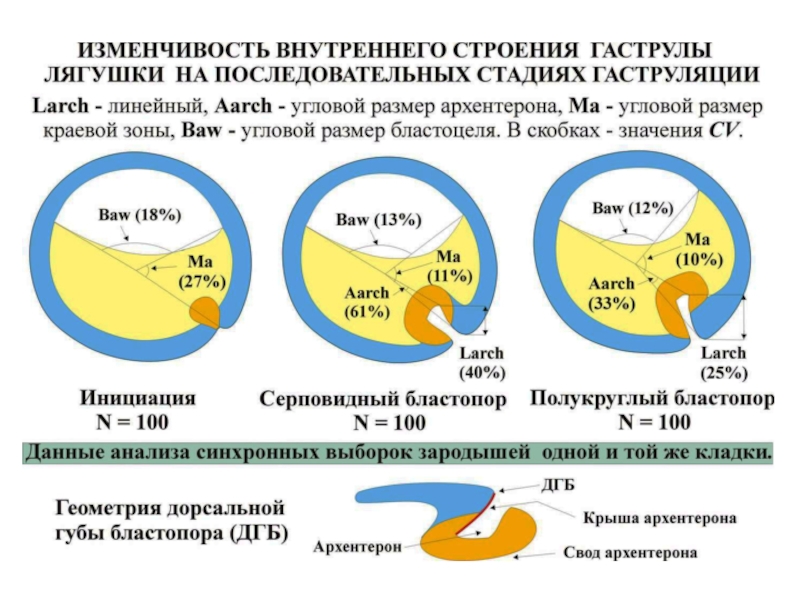

: Компоненты")
 образуют «детерминированный хаос»: анализировать можно")
")


 структур A – ротовой диск")
")

 упорядочивается по мере формирования борозды гастрального впячивания (Б)АБТо же самое видно")

: при эпиболии")





 вытесняет исходную кривизну окружности краевой зоны (тонкие стрелки) подобно тому,")
: простейшая (гаплоидная) модельОтбор максимизирует среднюю приспособленность W* со скоростью,")


 изменчивы не меньше, чем морфологияИзменчивость")



Гетерохронии между входом и выходом клеток из краевого валика")
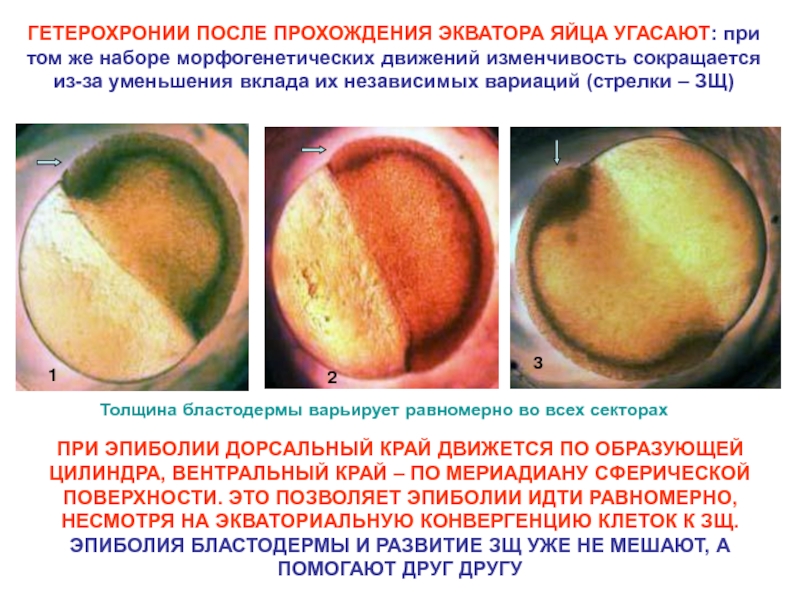


 – это прямая и обратная связь формы с организацией ее изменчивости на")
 новые МК возникают на последовательных стадиях развития")



 гена eve, маркирующего переднюю половину каждого сегмента,")






