- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Цитологические основы наследственности презентация
Содержание
- 1. Цитологические основы наследственности
- 2. Значение цитологического метода Развитие клеточной теории во
- 3. Уже в 1855 г. Р.Вирхов выдвинул фундаментальное
- 4. В 1862 г. Французская Академия удостоила премии
- 5. Тем самым было положено начало пристальному изучению
- 6. В 1884 г. Э. Страсбургер выделили такие
- 7. Цитологический метод широко используется в генетике для
- 8. Клеточный цикл Клеточный цикл состоит из двух
- 9. Схема регуляции клеточного цикла Клеточный цикл
- 10. Фазы клеточного цикла и митоза Временной
- 11. Митоз
- 12. Метафаза Анафаза
- 13. Мейоз и формирование гамет Мейо́з (от др.-греч. μείωσις — уменьшение) или редукционное
- 14. В зависимости от места в жизненном цикле
- 16. БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ МЕЙОЗА Биологическое значение мейоза впервые
- 17. Поведение хромосом в мейозе сопоставимо с поведением
- 18. Хромосомная теория наследственности — теория, согласно которой
- 19. Спасибо за внимание

Слайд 2Значение цитологического метода
Развитие клеточной теории во второй половине XIX века создало

Слайд 3Уже в 1855 г. Р.Вирхов выдвинул фундаментальное положение “Omnis cellula e
Ру́дольф Людвиг Карл Ви́рхов (1821 — 1902) — немецкий учёный и политический деятель второй половины XIX столетия, врач, патологоанатом, гистолог, физиолог, один из основоположников клеточной теории в биологии и медицине, основоположник теории клеточной патологии в медицине; был известен также как археолог, антрополог и палеонтолог.

Слайд 4В 1862 г. Французская Академия удостоила премии мемуар Л. Пастера «О
Так была обоснована невозможность самозарождения жизни в настоящий геологический период.
Так стали утверждаться представления о том, что жизнь – это непрерывный ряд воспроизведения клеток
Луи́ Пасте́р (правильно Пастёр, 1822 — 1895) —французский микробиолог и химик, член Французской академии (1881). Пастер, показав микробиологическую сущность брожения и многих болезней человека, стал одним из основоположников микробиологии и иммунологии. Его работы в области строения кристаллов и явления поляризации легли в основу стереохимии. Также Пастер поставил точку в многовековом споре о самозарождении некоторых форм жизни в настоящее время, опытным путем доказав невозможность этого. Его имя широко известно в ненаучных кругах благодаря созданной им и названной позже в его честь технологии пастеризации.

Слайд 5Тем самым было положено начало пристальному изучению процесса клеточного деления –
Флемминг обнаружил, что при митозе хромосомы «делятся» вдоль, а Э. ван Бенеден (1883) обратил внимание на то, что дочерние хромосомы, распределяющиеся между дочерними клетками, до мельчайших деталей повторяют строение материнской хромосомы
Вальтер Флемминг (1843— 1905)
Генрих Вильгельм Вальдейер
(1836—1921)
Мари Жозеф Эдуард ван Бенеден (1846-1910)

Слайд 6В 1884 г. Э. Страсбургер выделили такие стадии митоза, как профаза
Эдуард Адольф Страсбургер
(1844—1912)
Гертвиг, Оскар
(1849 – 1922).
УИЛСОН (Wilson) (Вильсон) Эдмунд Бичер (1856-1939) - американский цитолог, один из основоположников цитогенетики, президент Нью-Йоркской АН (с 1904). На примере насекомых выяснил (1905-12) хромосомный механизм определения пола. Внес значительный вклад в изучение строения и физиологии клетки, предложил (1925) коллоидную теорию протоплазмы. Фундаментальная монография по цитологии (в рус. переводе - "Клетка и ее роль в развитии и наследственности", т. 1-2, 1936-40).

Слайд 7Цитологический метод широко используется в генетике для непосредственного изучения клеточных структур
Рис. Вверху: схема поворота ротора (показан красным цветом) ДНК-мотора (то есть перехода из B-формы в Z-форму) при увеличении концентрации ионов Mg2+ и обратного перехода при уменьшении концентрации ионов. Внизу: изображения, полученные при помощи атомной силовой микроскопии и подтверждающие, что переход произошел. Рисунок из обсуждаемой статьи в Journal of American Chemical Society
Группа японских ученых сконструировала наномотор, работающий за счет смены конформации молекулой ДНК — перехода двойной спирали из правозакрученной (B-форма) в левозакрученную (Z-форма). «Запуск» мотора осуществляется путем изменения концентрации ионов в растворе. Закрепив мотор в жесткой рамке, которая фиксирует его на подложке, не мешая при этом вращению ротора, ученые смогли при помощи атомной силовой микроскопии проследить за работой мотора в режиме реального времени. Они зафиксировали два оборота ротора, что соответствует одному B-Z-переходу.

Слайд 8Клеточный цикл
Клеточный цикл состоит из двух периодов: митоз (М-фаза) включает разделение предварительно удвоенного
стадию роста G1,
фазу репликации ДНК (S),
фазу подготовки к делению G2.
 включает разделение предварительно удвоенного ядерного материала, деление ядра,")
Слайд 9Схема регуляции клеточного цикла
Клеточный цикл регулируется генами и белками-ферментами двух
В делении дрожжевой клетки основные роли играют один Cdk и девять циклинов, которые образуют девять разных циклин-Cdk комплексов. У гораздо более сложно организованных млекопитающих изучено шесть Cdk и полтора десятка циклинов. Контроль выхода клетки из G1-, и G2-фаз осуществляют промотор-фактор S-фазы (SPF) и промотор-фактор M-фазы (MPF), представляющие собой гетеродимеры. Cуществует особая контрольная точка клеточного цикла (Start), с которой заканчивается рост (G1-фаза) и начинается процесс синтеза ДНК. Постулируется существование фактора транскрипции SBF, который может быть в активной Sa- и пассивной Si -форме. Он переходит в активную форму под действием циклина Cln (N) и Start-киназы (Cdc28-Cln3) (A) и инактивируется другим веществом (Е). Циклин продуцируется путем активации SBF и деградирует. SBF активируется Chu и Start-киназой и инактивируется фосфатазой
")
Слайд 10Фазы клеточного цикла и митоза
Временной ход митоза и цитокинеза, типичный для
Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой (от слова «митоз»). М-фазу условно подразделяют на шесть стадий, постепенно и непрерывно переходящих одна в другую. Первые пять — профаза, прометафаза (метакинез), метафаза, анафаза и телофаза (или цитотомия) — составляют митоз, а берущий своё начало в анафазе процесс разделения цитоплазмы клетки, или цитокинез, протекает вплоть до завершения митотического цикла и, как правило, рассматривается в составе телофазы.
Только в растительных клетках профазе предшествует дополнительная стадия, известная как препрофаза.
Длительность отдельных стадий различна и варьируется в зависимости от типа ткани, физиологического состояния организма, внешних факторов.



Слайд 13Мейоз и формирование гамет
Мейо́з (от др.-греч. μείωσις — уменьшение) или редукционное деление клетки — деление ядра эукариотической клетки с уменьшением числа хромосом в
 или редукционное деление клетки — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в")
Слайд 14В зависимости от места в жизненном цикле организма различают три осн.
Общая схема последовательных стадий мейоза: а — лептотена; б — зиготена; в — пахитена; г — диплотена; д — диакинез; е — метафаза I; ж — анафаза I; з — телофаза I; и — интеркинез; к — метафаза II; л — анафаза II; м — телофаза II.

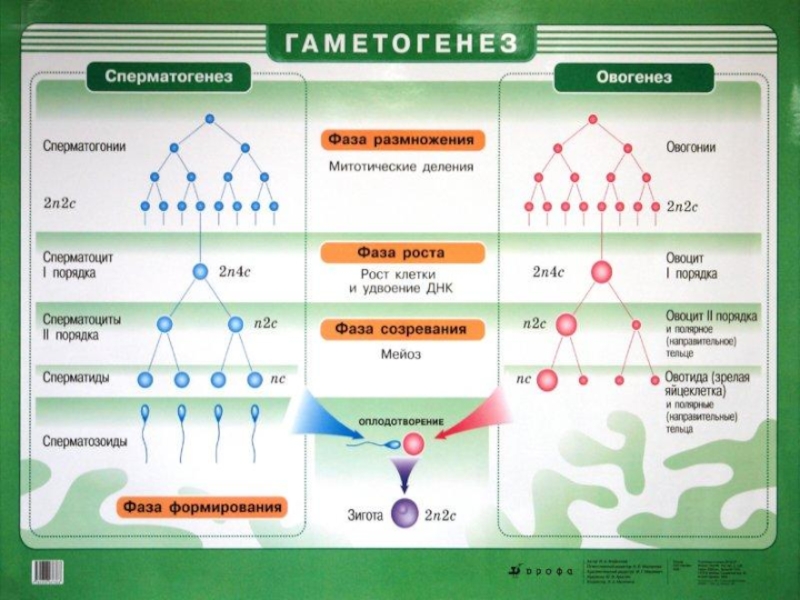
Слайд 16БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ МЕЙОЗА
Биологическое значение мейоза впервые оценил А. Вейсман, который отметил, что
Август Вейсман
(1834-1914, Германия)

Слайд 17Поведение хромосом в мейозе сопоставимо с поведением наследственных факторов (генов) в
Действительно, гаплоидные гаметы содержат по одной из каждой пары хромосом и соответственно по одному менделевско- му фактору, или аллеломорфу.
Слияние гаплоидных гамет приводит к объединению в ядре по одной гомологичной хромосоме от каждого родителя, а если они различались по аллелям одного гена, то и к установлению гетерозиготности (Аа).
При мейозе происходит редукция числа хромосом, при этом гомологи из разных бивалентов расходятся в анафазе I независимо и также независимо ведут себя разные пары менделевских факторов (АаВЬ), образуя в итоге 4 типа гамет: АВ, Ab, аВ, аЬ.
F1
Ab
 в открытых Г. Менделем закономерностях")
Слайд 18
Хромосомная теория наследственности — теория, согласно которой хромосомы, заключённые в ядре клетки,