Система дыхание
Лекция № 2
2016 В.М.
Система дыхание
Лекция № 2
2016 В.М.

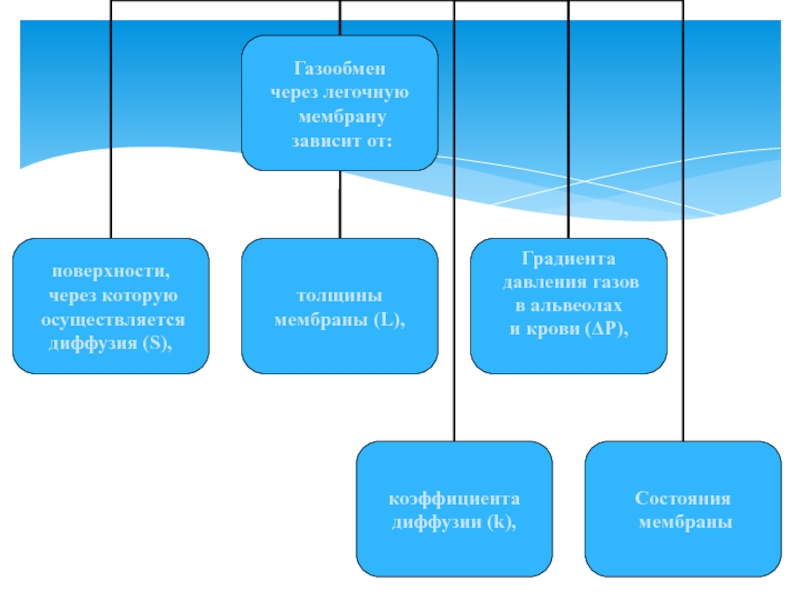
Закон Фика
 зависит от природы газа,")








Доставка О2 тканям
.")

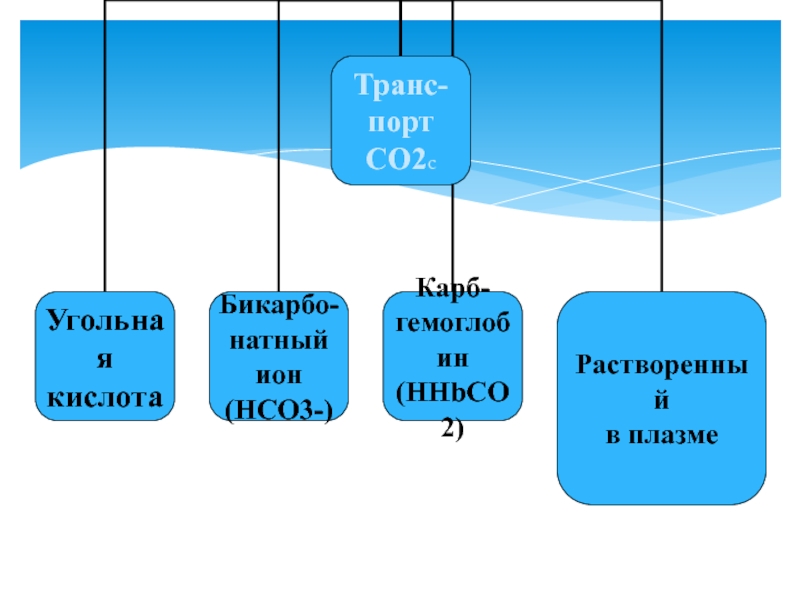
Обмен СО2 между тканями и кровью


Диссоциация СО2

*а) Н++НСО3-<==> H2СО3<==>Н2О+СО2
б) ННbCO2 + O2 <==> HHbO2 + CO2 <==> HbO2 + H+ + CO2
* - участие карбоангидразы
На нижней кривой представлена зависимость газообмена в легких от времени: СО2 выводится быстро, а О2 поступает более медленно. Поэтому при увеличении скорости тока крови (физ. работа) эритроцит может недополучать О2.
*а) Н++НСО3- H2СО3Н2О+СО2б) ННbCO2")
РЕГУЛЯЦИЯ ДЫХАНИЯ
 спинного мозга.Поэтому дыхание можно")

 несколько отличаются.Это обусловлено: а) при")
 автоматия обусловлена лишь свойствами водителей ритма, то в дыхательном")
 состоит из:2 -дорсального ядра (ведущее), 3 – вентрального ядра, 4")

Автоматия дыхательного центра

Межнейронные взаимодействия дыхательного центра
/ГЕНЕРАТОР РИТМА ДЫХАНИЯ/
I-α
I-β
Спинной мозг
Мотонейроны вспомогательных мышц вдоха и выдоха
Спинной мозг
Мотонейроны основных мышц вдоха
Возбуждение
Торможение


Регуляция дыхания при одышке


Центральные хеморецепторы
. Нахождение их")
Периферические хеморецепторы


Рецепторы легких и дыхательных путей
 Р. растяжения легких – расположены в гладких мышцах воздухоносных путей (активируя Iβ-нейроны, тормозящие активность")
Другие рецепторы легких
 Рецепторы дыхательных мышц . Функционируют по принципу гамма-петли межреберных и мышц стенок живота (при")
Ирритантные рецепторы

J-рецепторы (юкстамедуллярные рецепторы)

Схема всех механизмов, которые могут участвовать в регуляции дыхания

Если не удалось найти и скачать презентацию, Вы можете заказать его на нашем сайте. Мы постараемся найти нужный Вам материал и отправим по электронной почте. Не стесняйтесь обращаться к нам, если у вас возникли вопросы или пожелания:
Email: Нажмите что бы посмотреть
Это сайт презентаций, докладов, проектов, шаблонов в формате PowerPoint. Мы помогаем школьникам, студентам, учителям, преподавателям хранить и обмениваться учебными материалами с другими пользователями.