- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Сцепление генов и картирование хромосом презентация
Содержание
- 1. Сцепление генов и картирование хромосом
- 2. 1903 г. Вальтер Саттон – «единиц наследственности»
- 3. нарушение закона независимого комбинирования признаков (1906)
- 4. 1919 г. Дж. Холдейн такое расщепление
- 5. Сцепление и кроссинговер Т.Х. Морган и его
- 6. Сцепление генов соотношение фенотипов при сцеплении при
- 7.
- 8.
- 9. В результате контрольного скрещивания с мухами первого
- 10. Неполное сцепление, кроссинговер и хромосомное картирование Обычно
- 11. Работы Моргана по кроссинговеру Морган первым описал
- 12. При скрещивании по другим Х-сцепленным генам картина
- 13. (1) В чем причина разделения сцепленных генов?
- 14. при скрещиваниях по мутантным генам yellow-white происходит
- 15. Морган предположил, что частота формирования хиазм (рекомбинаций)
- 16. Работы Стертеванта по картированию генов Альфред Стертевант
- 17. гены располагаются в следующем порядке: yellow –
- 18. (1) yellow, white 0,5% (2) white, miniature 34,5% (3) yellow, miniature 35,4%
- 19. по частоте рекомбинаций между генами можно оценить
- 20. Позже Стертевант построил более подробную карту Х-хромосомы,
- 22. Одиночный кроссинговер Чем ближе расположены два локуса
- 23. Кроссинговер между двумя отдаленными локусами приводит к появлению рекомбинантных гамет.
- 24. Если между двумя несестринскими хроматидами происходит одиночный
- 25. Если два гена находятся на расстоянии более
- 26. Множественный кроссинговер В одной и той же
- 27. Вероятность единичного обмена между генами А и
- 28. Допустим, что обнаружено 20% кроссоверных гамет,
- 29. расстояние между генами А и В равно
- 30. Картирование генов у Drosophila и кукурузы Анализ
- 31. Трехлокусное картирование у Drosophila Для проведения экспериментального
- 32. В скрещивании анализируются три сцепленных с полом
- 33. В F1 все самки имеют фенотип дикого
- 34. У самок F1 гаметы могут нести нерекомбинантные
- 35. В потомстве можно выделить группы, состоящие из
- 36. Мухи с фенотипом yellow, echinus, white или
- 37. В F2 появляются и два реципрокных фенотипа:
- 38. Оставшиеся четыре фенотипических класса получаются в результате
- 39. Интерференция и коэффициент коинцедентности зная межгенное расстояние
- 40.
- 41. Зная коэффициент коинцедентности, легко определить величину
- 42. I – положительная величина, показывающая, что частота
- 43. Генетическая карта Drosophila У таких организмов, как
- 44. Гибридизация соматических клеток и картирование генов человека
- 45. метод гибридизации соматических клеток позволяет эффективно определять
- 46. В процессе культивирования гетерокарионов in vitro происходят
- 47. Если в синкарионе с 1 – 3
- 48. Проверка корреляции наличия или отсутствия определенной хромосомы
- 49. С помощью гибридизации соматических клеток и синтенного тестирования были локализованы сотни человеческих генов.
- 50. Митотический кроссинговер В 1936 г. К. Штерн
- 51. Появление таких двойных пятен К. Штерн объяснил
- 52. Сестринские хроматидные обмены Гомологичные хромосомы в соматических
- 53. СХО были обнаружены благодаря специальным методам окраски
- 54. Значение СХО до сих пор не совсем
- 55. Частота СХО повышается при некоторых наследственных заболеваниях
- 57. Хромосомные основы наследственности
- 58. Наследование, сцепленное с полом анализ отклонений от
- 59. красноглазые самки и белоглазые самцы в
- 60. При скрещивании белоглазых самок с красноглазыми самцами результаты были другими.
- 61. эти результаты можно объяснить, если предположить:
- 62. Морган пришел к заключению, что белый цвет
- 63. Нерасхождение Х-хромосом при скрещивании белоглазых самок дрозофилы
- 64. гипотеза Бриджеса: появляющиеся с частотой
- 65. Вторичное нерасхождение Бриджес скрещивал таких самок (ХХY)
- 66. Нерасхождение может быть следствием физического сцепления Х-хромосом,
- 67. Определение пола У D.melanogaster носители единственной Х-хромосомы,
- 68. Хромосомное определение пола у Drosophila у Drosophila
- 69. Ранние исследования Х- и Y-хромосом 1891 г.
- 70. Этот механизм получил название ХХ/Х0, или определение
- 71. Вильсон – определение пола у насекомого Lygaeus
- 72. У Protenor и Lygaeus самцы образуют неодинаковые
- 73. У мотыльков, бабочек, большинства птиц, некоторых рыб,
- 74. Определение пола у пчел и муравьев широко распространен механизм определения пола, называемый гаплодиплоидией
- 76. Сцепленное с полом наследование у человека и
- 77. пример сцепленного с полом наследования гемофилии представляет
- 78. Хромосомное определение пола у человека 1912 г.
- 79. 1956 г. Дж.Х. Тио и А. Ливен
- 80. Y-хромосома для генов, содержащихся только в Y-хромосоме,
- 81. Синдромы Клайнфельтера и Тернера В начале 40-х
- 82. У лиц с синдромом Клайнфельтера имеется лишняя
- 83. Пробанды с синдромом Тернера обычно имеют только
- 84. Анализ кариотипов и фенотипов у пробандов с
- 85. Синдром Клайнфельтера встречается примерно у 2-х из
- 86. Синдром Тернера может проявиться и у больных
- 87. Синдром 47,ХХХ Носительницы трех Х-хромосом (47,ХХХ) обнаруживаются
- 88. Синдром 47,ХYY Исследование этого синдрома было впервые
- 89. Оказалось, что среди уголовников частота мужчин с
- 90. Y-хромосома и мужской тип развития На концах
- 91. Некоторые гены Y-хромосомы кодируют продукт, запускающий развитие
- 92. ТДФ кодируется геном, локализованным на коротком плече
- 93. этот вывод был подкреплен результатами, полученных на

Слайд 21903 г. Вальтер Саттон – «единиц наследственности» у большинства организмов значительно
Горох n = 2
Дрозофила n = 4
Аскарида n = 1
Каждая хромосома должна быть детерминантом не одного, а нескольких элементарных признаков.
Графическое представление нормального человеческого кариотипа в виде идеограмм всех его хромосом

Слайд 3нарушение закона независимого комбинирования
признаков (1906) – У.Бэтсон, Р.Пеннет
душистый горошек
окраска цветка – пурпурная (Р) или красная (р)
форма пыльцевых зерен – удлиненная (L) или круглая (l)
пурпурные цветки, удлиненная пыльца (PP LL) х красные цветки, круглая пыльца (pp ll)
F1: пурпурные цветки, удлиненная пыльца (Pp Ll).
самоопыление
F2: Пурпурные цветки, удлиненная пыльца (P- L-) 4831 (69,5 %)
Пурпурные цветки, круглая пыльца (P- ll) 390 (5,6 %)
Красные цветки, удлиненная пыльца (pp L-) 393 (5,6 %)
Красные цветки, круглая пыльца (pp ll) 1338 (19,3 %)
 – У.Бэтсон, Р.Пеннет душистый горошек – Lathyrus odoratus.окраска цветка")
Слайд 41919 г. Дж. Холдейн
такое расщепление может получиться, если четыре типа
0,44 PL : 0,06 Pl : 0,06 pL : 0,44 pl
родительские сочетания аллелей исследованных генов PL и pl предпочтительно попадают в одни и те же гаметы, в то время как их новые рекомбинантные сочетания (pL и Pl) встречаются гораздо реже.
-сцепление генов.

Слайд 5Сцепление и кроссинговер
Т.Х. Морган и его сотрудники в экспериментах с D.

Слайд 6Сцепление генов
соотношение фенотипов при сцеплении при двух тесно сцепленных рецессивных мутаций
brown (bw) – коричневые глаза
heavy (hv) – утолщенные жилки крыла.
Оба аллеля bw+ и hv+ доминантны и проявляются в фенотипе как красные глаза и тонкие жилки на крыльях.

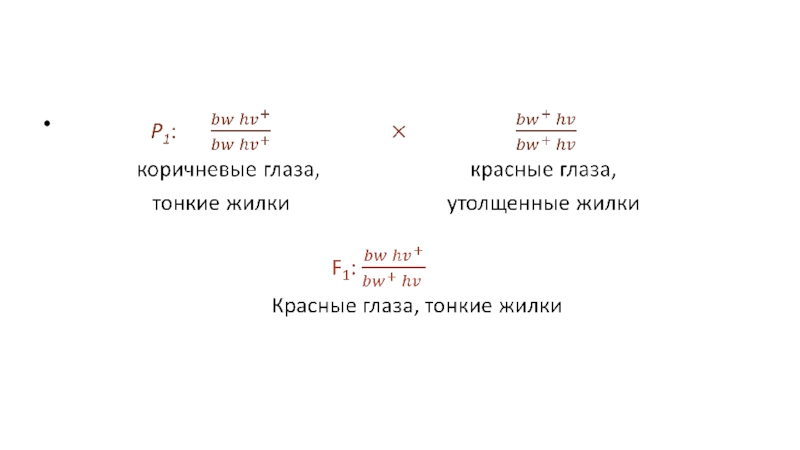

Слайд 9В результате контрольного скрещивания с мухами первого поколения получаются коричневоглазые мухи
Анализ большого числа мутантных генов, локализованных на одной и той же хромосоме, обнаруживает, что эти гены входят в одну группу сцепления.

Слайд 10Неполное сцепление, кроссинговер и хромосомное картирование
Обычно при скрещиваниях по двум сцепленным
Частота рекомбинаций между этими генами зависит от меженного расстояния.
Эту закономерность впервые исследовали в 1911 г. Томас Х. Морган и Альфред Х. Стертевант.

Слайд 11Работы Моргана по кроссинговеру
Морган первым описал Х-сцепленные гены у Drosophila, анализируя
При скрещивании мутантных мух с желтым телом (y – yellow) и белыми глазами (w – white) с мухами дикого типа (серое тело красные глаза).
F1 все самки дикого типа,
у самцов проявляются оба мутантных признака.
F2 98,7% мух имеют родительские фенотипы:
желтое тело, белые глаза или серое тело, красные глаза.
1,3% мух имеют либо желтое тело, красные глаза,
серое тело, белые глаза.

Слайд 12При скрещивании по другим Х-сцепленным генам картина оказалась похожей, но пропорции
Во втором поколении от скрещивания мутантных белоглазых мух с миниатюрными крыльями
62,8% мух имели родительские фенотипы,
37,2% мух имели другие комбинации родительских признаков.

Слайд 13(1) В чем причина разделения сцепленных генов?
(2) Почему частота рекомбинаций варьирует
уже были известны результаты цитологических наблюдений Ф.А. Янссенса, который в 1909 г. обнаружил хиазмы (точки перекреста) во время синапса гомологичных хромосом в мейозе у земноводных.
Морган предположил, что хиазмы как раз и являются точками генного обмена между хромосомами.
 В чем причина разделения сцепленных генов?(2) Почему частота рекомбинаций варьирует в зависимости от изучаемых")
Слайд 14при скрещиваниях по мутантным генам yellow-white происходит 1,3% рекомбинаций,
при скрещиваниях

Слайд 15Морган предположил, что частота формирования хиазм (рекомбинаций) между двумя тесно сцепленными
Для описания процесса обмена между хроматидами, приводящего к рекомбинации генов Морган ввел понятие кроссинговера.
 между двумя тесно сцепленными генами ниже, чем между")
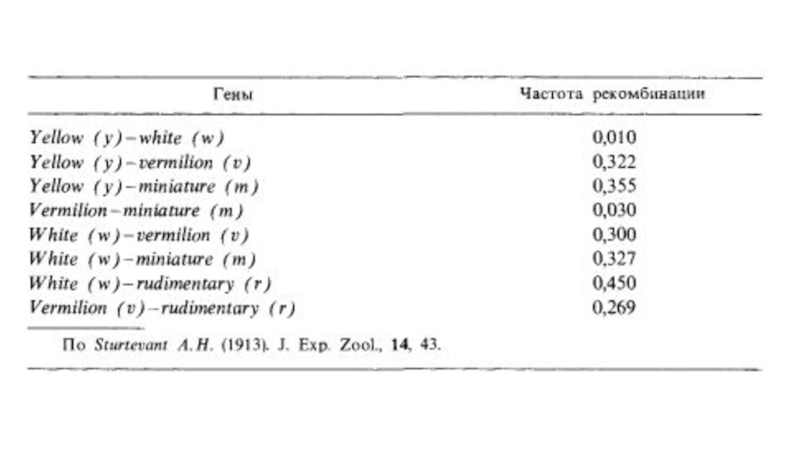
Слайд 16Работы Стертеванта по картированию генов
Альфред Стертевант – картирования генов на хромосомах.
исследование
частоты кроссинговера между отдельными генами в разных скрещиваниях:
(1) yellow, white 0,5%
(2) white, miniature 34,5%
(3) yellow, miniature 35,4%

Слайд 17гены располагаются в следующем порядке: yellow – white – miniature.
(1) yellow,
(2) white, miniature 34,5%
(3) yellow, miniature 35,4%
 yellow, white 0,5%(2) white, miniature 34,5%(3) yellow, miniature 35,4%")
 yellow, white 0,5%(2) white, miniature 34,5%(3) yellow, miniature 35,4%")
Слайд 19по частоте рекомбинаций между генами можно оценить меженное расстояние.
Исходя из частот
1 единица карты соответствует 1% рекомбинации между генами.

Слайд 20Позже Стертевант построил более подробную карту Х-хромосомы, включающую 5 генов.
В результате

Слайд 22Одиночный кроссинговер
Чем ближе расположены два локуса на хромосоме, тем вероятность кроссинговера
и наоборот:
чем дальше эти локусы друг от друга, тем вероятность кроссоверов между ними выше.


Слайд 24Если между двумя несестринскими хроматидами происходит одиночный кроссинговер, то две другие
теоретически, частота рекомбинантных гамет не может превышать 50%.

Слайд 25Если два гена находятся на расстоянии более 50 сМ, то кроссинговер

Слайд 26Множественный кроссинговер
В одной и той же тетраде может произойти два, три
Двойной кроссинговер – двойные обмены.
многократный кроссинговер между гомологичными хромосомам

Слайд 27Вероятность единичного обмена между генами А и В или В и
чем ближе они расположены, тем меньше вероятность рекомбинации между двумя из этих генов.
В случае двойного кроссинговера одновременно происходят два независимых межгенных обмена.
Вероятность двух независимых событий равна произведению вероятностей каждого из них.
Двойной кроссинговер

Слайд 28Допустим, что обнаружено
20% кроссоверных гамет, возникших в результате рекомбинации между
30% кросоверных гамет – в результате рекомбинации между генами В и С (р = 0,30).
вероятность двойного кросинговера между генами А и В, В и С, приводящего к появлению кросоверных гамет, равна
(0,20)(0,30) = 0,06 = 6%.

Слайд 29расстояние между генами А и В равно 3 единицам карты,
расстояние
частота двойного кроссовера равна
(0,03)(0,02) = 0,0006, то есть всего 0,06%, или
6 кроссоверов на 10 000.
В данном случае, двойной кроссинговер трудно выявить даже среди 1000 потомков, поскольку его частота крайне низкая.

Слайд 30Картирование генов у Drosophila и кукурузы
Анализ последствий двойного кроссинговера позволяет картировать

Слайд 31Трехлокусное картирование у Drosophila
Для проведения экспериментального скрещивания необходимы следующие условия:
1. Родитель,
2. В потомстве должны точно определяться рекомбинантные фенотипы, поскольку генотипы гамет недоступны для прямого анализа.
3. Для выявления всех кроссоверных фенотипов и картирования генов нужно проанализировать достаточное количество потомков.

Слайд 32В скрещивании анализируются три сцепленных с полом рецессивных мутантных гена:
yellow
white– белые глаза
echinus – ежевидные глаза.

Слайд 33В F1 все самки имеют фенотип дикого типа, поскольку они гетерозиготны
В результате кроссинговера между этими генами самки F1 могут давать рекомбинантные гаметы.

Слайд 34У самок F1 гаметы могут нести нерекомбинантные Х-хромосомы, которые попадают к
Если одна из Х-хромосом несет один мутантный аллель, а вторая – два других мутантных аллеля, то образуются реципрокные классы гамет и фенотипов.

Слайд 35В потомстве можно выделить группы, состоящие из двух реципрокных фенотипических классов,
при участии некроссоверных гамет (NCO),
гамет с одной рекомбинацийе между этими генами (SCO)
дважды кросоверных гамет (DCO).

Слайд 36Мухи с фенотипом yellow, echinus, white или мухи дикого типа составляют

Слайд 37В F2 появляются и два реципрокных фенотипа:
с нормальной окраской глаз,
c нормальной окраской тела и формой глаз, но белоглазых (мутация white).
В сумме эти два класса составляют только 0,06% всего потомства F2.

Слайд 38Оставшиеся четыре фенотипических класса получаются в результате одиночных кроссоверов:
реципрокные фенотипы
Фенотипы классов, возникающих вследствие одиночных кросоверов между локусами white и echinus составляют 4,00% мух второго поколения.
Расстояние между локусами у и w или w и ес равно частоте всех рекомбинаций, обнаруженных между этими локусами, включая одиночные и двойные кроссоверы
В случае локусов у и w следует сложить частоты рекомбинантных фенотипов:
1,50 + 0,06 = 1,56 единиц карты.
Аналогично, расстояние между локусами ec и w равно частоте всех рекомбинаций между этими локусами:
4,00 + 0,06 = 4,06 ед. карты.

Слайд 39Интерференция и коэффициент коинцедентности
зная межгенное расстояние можно оценить частоту множественных рекомбинаций,
расстояние между генами v и pr у кукурузы равно 22,3 ед. карты,
между генами pr и bm – 43,4 ед. карты.
Если два одиночных кроссовера происходят независимо и одновременно, то ожидаемая частота двойного кроссовера равна:
DCOexp = (0,223) х (0,434) = 0,997 = 9,7%.


Слайд 41
Зная коэффициент коинцедентности, легко определить величину интерференции
I = 1 – C
В
I = 1,000 – 0,804 = 0,196.

Слайд 42I – положительная величина, показывающая, что частота наблюдаемых двойных кроссоверов на
При полной интерференции двойных кроссоверов в данном районе хромосомы вообще не наблюдается, то есть I = 1,0.
Чем ближе расположены гены, тем выше положительная интерференция.
У Drosophila полная интерференция наблюдается на расстоянии 10 ед. карты и на участке такой длины уже не происходит двойных или множественных кроссоверов.

Слайд 43Генетическая карта Drosophila
У таких организмов, как дрозофила, кукуруза, мышь, известно множество

Слайд 44Гибридизация соматических клеток и картирование генов человека
У человека невозможны контрольные скрещивания
 с многочисленным потомством,")
Слайд 45метод гибридизации соматических клеток
позволяет эффективно определять сцепление генов с определёнными хромосомами
Георгий Барский
в культуре можно индуцировать слияние двух разнородных клеток в одну гибридную.
Сначала Барский использовал две мышиных клеточных линии, но вскоре убедился, что к слиянию способны и клетки, принадлежащие разным организмам, в результате образуется гетерокарион.

Слайд 46В процессе культивирования гетерокарионов in vitro происходят два важных события.
Ядра родительских
По мере дальнейшего культивирования гибридных клеток хромосомы одного из родителей постепенно утрачиваются.

Слайд 47Если в синкарионе с 1 – 3 человеческими хромосомами синтезируется продукт
Можно создавать панели гибридных клеток, каждая линия которых содержит только одну из 23 хромосом человека, что позволяет картировать любой ген по наличию или отсутствию его продукта.

Слайд 48Проверка корреляции наличия или отсутствия определенной хромосомы с наличием или отсутствием

Слайд 49С помощью гибридизации соматических клеток и синтенного тестирования были локализованы сотни

Слайд 50Митотический кроссинговер
В 1936 г. К. Штерн исследовал мух D. melanogaster генотипа
Мутация у – желтое тело, sn (singed) – опаденные щетинки.
Оба гена находятся в Х-хромосоме, а центромера расположена справа от гена sn.
Дигетерозиготы – дикий фенотип по обоим признакам, однако изредка не теле некоторых мух появлялись двойные пятна:
половина пятна - желтая с нормальными щетинками,
другая половина – нормального серого цвета, но покрытая опаленными щетинками.

Слайд 51Появление таких двойных пятен К. Штерн объяснил митотическим кроссинговером на стадии
если такой обмен произойдет, то при расхождении хромосом в митозе в половине случаев должны образовываться двойные пятна.
Частота митотического кроссинговера значительно ниже (на 2 – 3 порядка) мейотического.

Слайд 52Сестринские хроматидные обмены
Гомологичные хромосомы в соматических клетках обычно не образуют синапса
Оказалось, что между сестринскими хроматидами возможен реципрокный обмен, сходный с кроссинговером.
Хотя сестринские хроматидные обмены (СХО) не приводят к появлению новых аллельных комбинаций, они имеют важное значение.
, и")
Слайд 53СХО были обнаружены благодаря специальным методам окраски хромосом.
репликация ДНК в присутствии
После двух циклов репликации, когда БУдР встраивается во вновь синтезированную цепь ДНК, одна хроматида в каждой паре сестринских хроматид оказывается «меченой» БудР.
С помощью флуоресцентных красителей можно отличить хроматиды друг от друга и легко обнаружить СХО.
На них видны многочисленные межхроматидные обмены, так что некоторые хромосомы выглядят мозаичными, их называют хромосомами типа арлекин.
.После двух циклов")
Слайд 54Значение СХО до сих пор не совсем понятно.
агенты, индуцирующие повреждение ДНК
Вирусы
рентгеновское и ультрафиолетовое излучение
химические мутагены

Слайд 55Частота СХО повышается при некоторых наследственных заболеваниях
синдром Блума, ассоциированный с
наследуется по рецессивному типу
сопровождается
пренатальной задержкой роста
высокой чувствительностью кожи лица к ультрафиолету
Иммунодефицитом
предрасположенностью к злокачественным и доброкачественным опухолям
аномальным поведением.
метафазные хромосомы в культуре клеток костного мозга, лейкоцитов и фибробластов полученных от гомозигот по этой мутации, несут множество ломких сайтов и по сравнению с хромосомами гетерозигот или здоровых людей чаще вступают в СХО или перестройки типа разрыв-воссоединение.
Джеймс Герман с коллегами обнаружил, что ген BLM кодирует ДНК-геликазу
репликация ДНК.



Слайд 58Наследование, сцепленное с полом
анализ отклонений от ожидаемых результатов скрещиваний.
Морган использовал для

Слайд 59красноглазые самки и белоглазые самцы
в F1 все мухи имели красные глаза
в F2 все самки были красноглазыми, а среди самцов половина имела красные глаза, а половина – белые.


Слайд 61эти результаты можно объяснить, если предположить:
ген, определяющий цвет глаз, расположен
половая хромосома самцов (Y-хромосома) не содержит этот ген.
Клетки самок дрозофилы содержат по две Х-хромосомы, а клетки самцов – две разные хромосомы, Х и У.
Самцы же получают Х-хромосому от матери и передают ее лишь дочерям
")
Слайд 62Морган пришел к заключению, что белый цвет глаз у дрозофилы –
ген, определяющий этот признак, расположен в Х-хромосоме.

Слайд 63Нерасхождение Х-хромосом
при скрещивании белоглазых самок дрозофилы с красноглазыми самцами дочери оказываются
Бриджес предположил, что появление редких мух возможно при нерасхождении Х-хромосом;
Х-хромосомы не расходятся в мейозе к разным полюсам, а направляются вместе к одному из полюсов, в результате чего образуются яйцеклетки с двумя Х-хромосомами и яйцеклетки без Х-хромосом.

Слайд 64гипотеза Бриджеса:
появляющиеся с частотой 1 : 2000 белоглазые самки получают
а такие же редкие красноглазые самцы получают лишь одну отцовскую хромосому (и, следовательно, вовсе лишены Y-хромосомы).

Слайд 65Вторичное нерасхождение
Бриджес скрещивал таких самок (ХХY) с нормальными красноглазыми самцами (ХY).
4%
Бриджес предположил, что эти 4% составляют самки и самцы, возникающие снова в результате нерасхождения Х-хромосом в мейозе у самок.
Он назвал такое нерасхождение вторичным, поскольку оно происходит в потомстве самок, появившихся в результате первичного нерасхождения Х-хромосом (и поэтому обладающих двумя Х-хромосомами и одной Y-хромосомой).
 с нормальными красноглазыми самцами (ХY).4% самок имеют белые глаза,")
Слайд 66Нерасхождение может быть следствием физического сцепления Х-хромосом, в таком случае нерасхождение
Желтый цвет тела у Drosophila melanogaster определяется сцепленным с полом аллелем yellow (у), рецессивным по отношению к аллелю дикого типа (у+).
Морган обнаружила существование темных самок, которые при скрещивании с нормальными самцами давали в потомстве лишь желтых дочерей и нормальных сыновей, а не нормальных дочерей и желтых сыновей, как подавляющее большинство.

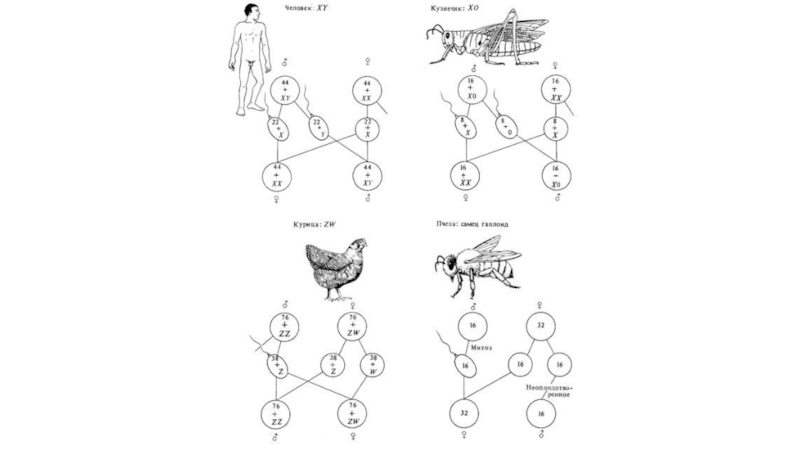
Слайд 67Определение пола
У D.melanogaster носители единственной Х-хромосомы, не имеющие Y-хромосомы, обладают нормальным
Фенотип по полу D.melanogaster определяется соотношением между числом Х-хромосом и числом наборов аутосом (А).
Если отношение Х/А = 1, муха имеет фенотип самки
если Х/А = ½ - фенотип самца
если Х/А больше 1/2, но меньше единицы, то развивается особь с фенотипом, промежуточным между мужским и женским, т.е. «интерсекс».
.Фенотип по")
Слайд 68Хромосомное определение пола у Drosophila
у Drosophila Y-хромосома не детерминирует мужской тип

Слайд 69Ранние исследования Х- и Y-хромосом
1891 г. Х. Хенкинг обнаружил в ядрах
Кларенс МакКланг – в некоторых спермиях кузнечиков содержится необычная структура, которую назвали гетерохромосомой, но в других спермиях ее не находили.
1906 г. Эдмунд Б. Вильсон – соматические клетки у самок насекомого (Protenor) содержат 14 хромосом, в том числе две Х-хромосомы.
При слиянии двух гамет, несущих Х-хромосомы зигота развивается в самку, а при оплодотворении яйцеклетки спермием без Х-хромосомы – в самца.

Слайд 70Этот механизм получил название ХХ/Х0, или определение пола типа Protenor
такая же

Слайд 71Вильсон – определение пола у насекомого Lygaeus turicus, оба пола у
У самок имеется 2 Х-хромосомы, а у самцов – 1 Х-хромосома и 1 маленькая гетерохромасома, которую он назвал Y-хромосомой.
Этот тип определения пола получил название тип Lygaeus, или ХХ/ХY.

Слайд 72У Protenor и Lygaeus самцы образуют неодинаковые по хромосомному составу гаметы,
Самки относятся к гомогаметному полу, дающему одинаковые по хромосомному составу гаметы.

Слайд 73У мотыльков, бабочек, большинства птиц, некоторых рыб, у рептилий, земноводных и,
половые хромосомы самцов и самок обозначают – ZZ/ZW
самки несут ZW-хромосомы.

Слайд 74Определение пола
у пчел и муравьев широко распространен механизм определения пола, называемый

Слайд 76Сцепленное с полом наследование у человека и других видов
Самцы называются гемизиготными
1911 год – Эдмунд Вильсон:
все известные факты дальтонизма можно объяснить, предположив, что дальтонизм является рецессивным сцепленным с полом аллелем, а мужской пол у людей гетерогаметен.

Слайд 77пример сцепленного с полом наследования гемофилии представляет собой родословное дерево некоторых

Слайд 78Хромосомное определение пола у человека
1912 г. Х. Винивартер обнаружил в метафазных
1920 г. Теофил Пейнер обнаружил в ткани семенников маленькую Y-хромосому и, в общем, около 45 – 48 хромосом.
Схема половых хромосом человека и сцепленных с ними генов:
1 — Х-хромосома; 2 — Y-хромосома.

Слайд 791956 г. Дж.Х. Тио и А. Ливен усовершенствовали метод приготовления хромосомных
Позже Л.Е. Форд и Дж.Л. Хамертон также исследовали ткань семенников и подтвердили этот вывод.

Слайд 80Y-хромосома
для генов, содержащихся только в Y-хромосоме, характерно голандрическое наследование, т.е. они

Слайд 81Синдромы Клайнфельтера и Тернера
В начале 40-х годов XX века у человека
У пробандов с синдромом Клайнфельтера имеются мужские половые гениталии и семявыносящие канальцы, но семенники рудиментарны, и спермы не образуется.
Кариотип 47, (XXY)

Слайд 82У лиц с синдромом Клайнфельтера имеется лишняя Х-хромосома и большинство из

Слайд 83Пробанды с синдромом Тернера обычно имеют только одну Х-хромосому и кариотип
У пробандов с синдромом Тернера имеются женские наружные и внутренние гениталии, однако яичники недоразвиты.
Причиной обоих заболеваний стало нерасхождение Х-хромосом во время мейоза.

Слайд 84Анализ кариотипов и фенотипов у пробандов с синдромами Клайнфельтера и Тернера
Наличие Y-хромосомы при синдроме Клайнфельтера обеспечивает мужской фенотип, хотя и с неполным проявлением признаков.
Однако без этой хромосомы в кариотипе у пробандов проявляются женские половые признаки, как при синдроме Тернера.

Слайд 85Синдром Клайнфельтера встречается примерно у 2-х из 1000 новорожденных мальчиков.
Синдром Тернера

Слайд 86Синдром Тернера может проявиться и у больных с мозаицизмом соматических клеток.
Таким

Слайд 87Синдром 47,ХХХ
Носительницы трех Х-хромосом (47,ХХХ) обнаруживаются примерно у 1 из 1200
Недоразвитие вторичных половых признаков
Стерильность
Умственная отсталость
 обнаруживаются примерно у 1 из 1200 новорожденных девочек.Недоразвитие вторичных половых")
Слайд 88Синдром 47,ХYY
Исследование этого синдрома было впервые предпринято в 1965 г. Патрицией
В одной из шотландских тюрем строгого режима среди 315 мужчин она обнаружила 9 носителей кариотипа 47,ХУУ.
Выше среднего роста
Асоциальное поведение
Уголовные преступления
Семь из этих девяти заключённых страдали умственными, и все девять – психическими расстройствами.
Кариотип мужчины с XYY-синдромом

Слайд 89Оказалось, что среди уголовников частота мужчин с ростом выше 1,8 м

Слайд 90Y-хромосома и мужской тип развития
На концах У-хромосомы находятся псевдоаутосомные районы (ПАР),
Оставшиеся негомологичные участки У-хромосомы называют нерекомбинирующими районами У-хромосомы (НРУ).
, которые гомологичны участкам Х-хромосомы")
Слайд 91Некоторые гены Y-хромосомы кодируют продукт, запускающий развитие недифференцированных зародышевых гонад в
Этот продукт назвали фактором, детерминирующим семенники, или тестис-детерминирующим фактором (ТДФ).

Слайд 92ТДФ кодируется геном, локализованным на коротком плече Y-хромосомы и названным пол-детерминирующим
SRY позволяет идентифицировать последовательности Y-хромосомы у индивидов, чей фенотип не соответствует кариотипу.
содержащий SRY-участок Y-хромосомы присоединен в результате хромосомной перестройки к Х-хромосоме.
.SRY позволяет")
Слайд 93этот вывод был подкреплен результатами, полученных на трансгенных мышах.
У мышей был
Если последовательности мышиного Sry инъецировать в яйцеклетки мыши с геномом ХХ, то большинство развившихся их них мышат будет самцами.






