- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Сократительная деятельность мышц презентация
Содержание
- 1. Сократительная деятельность мышц
- 2. План 1. Теории мышечного сокращения. 2.
- 4. Скелетные мышцы состоят из мышечных пучков, образованных
- 6. В миофибриллах скелетных
- 8. Саркоплазматический ретикулум состоит из системы поперечных, продольных
- 9. Саркомер ограничен с двух сторон так
- 11. В А-дисках миозиновые нити образуют гексагональную
- 15. В одной половине толстой протофибриллы (и
- 16. Иерархические уровни организации скелетной мышцы (по Bloom , 1968).
- 17. Хаксли и Хэнсон – теория скольжения нитей.
- 18. В ходе сокращения миозин становится способным
- 19. Схема сокращения 1 – актиновый
- 20. Сокращение мышцы
- 22. Главной «моторной» частью миозина скелетных мышц является
- 25. Предполагается, что поперечные мостики, отходящие от
- 26. Механизм сопряжения возбуждения и сокращения 1
- 27. ATP – АТФ ADP - АДФ Pi – ион фосфора
- 29. Actin Myosin Crossbridge 3D Animation*
- 30. Поступление потенциала действия в моторные концевые
- 31. Нервный импульс передается поперечными трубочкам, где он
- 32. Расслабление мышцы вызывается обратным переносом ионов
- 33. При одиночном мышечном сокращении различают 3
- 36. Одиночное сокращение и тетанус При воздействии импульсов,
- 37. Если период следования импульсов больше продолжительности одиночного
- 38. Used by permission of John W. Kimball
- 40. Происхождение зубчатого и гладкого тетануса. Б –

Слайд 2План
1. Теории мышечного сокращения.
2. Одиночное и тетаническое сокращение.
3. Теории тетануса.
4.
Формы и типы мышечного сокращения.


Слайд 4Скелетные мышцы состоят из мышечных пучков, образованных большим количеством мышечных волокон
Каждое волокно – это клетка цилиндрической формы диаметром 10-100 мкм и длиной от 5 до 400 мкм. Оно имеет клеточную мембрану – сарколемму.
В плазме мышечной клетки (саркоплазме) находится несколько ядер, митохондрии (30-35% от массы волокна), образования саркоплазматического ретикулума и сократительные элементы – миофибриллы.
Сама миофибрилла с свою очередь состоит из параллельно лежащих протофибрилл - актиновых и миозиновых нитей.


Слайд 6
В миофибриллах скелетных мышц наблюдается правильное чередование более светлых и более
темных участков. Поэтому скелетные мышцы называют поперечнополосатыми.
Миофибрилла состоит из одинаковых повторяющихся элементов, так называемых саркомеров.
Миофибрилла состоит из одинаковых повторяющихся элементов, так называемых саркомеров.


Слайд 8Саркоплазматический ретикулум состоит из системы поперечных, продольных трубочек и цистерн. Поперечные
трубочки – это впячивания саркоплазмы внутрь клетки. К ним примыкают продольные трубочки с цистернами. Благодаря этому, потенциал действия может распространятся от сарколеммы на систему саркоплазматического ретикулума.
Мембраны саркоплазматического ретикулума содержат кальций-АТФазные насосы, которые транспортируют ионы кальция.
Мембраны саркоплазматического ретикулума содержат кальций-АТФазные насосы, которые транспортируют ионы кальция.

Слайд 9
Саркомер ограничен с двух сторон так называемыми Z-дисками. К этим дискам
с двух сторон прикреплены тонкие актиновые волокна (более светлые) – I-зона.
В середине саркомера находятся более темные участки, образованные толстыми миозиновыми волокнами А – зона.
В середине саркомера находятся более темные участки, образованные толстыми миозиновыми волокнами А – зона.

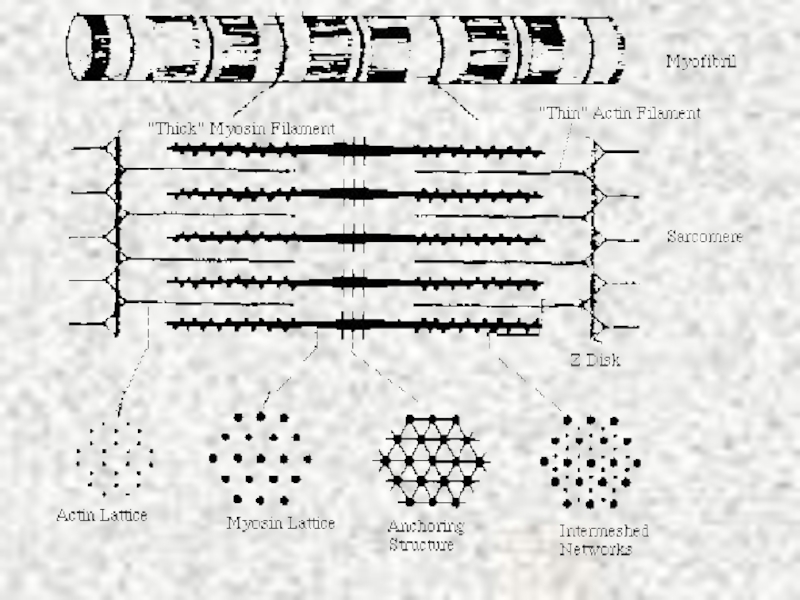
Слайд 11
В А-дисках миозиновые нити образуют гексагональную упаковку, именно они обусловливают высокую
оптическую плотность дисков. Актиновые нити прикрепляются с каждой стороны к узкой плотной мембране, т.называемой Z-мембране, которая пересекает I-диск . Расстояние между двумя Z-мембранами называется САРКОМЕРОМ.



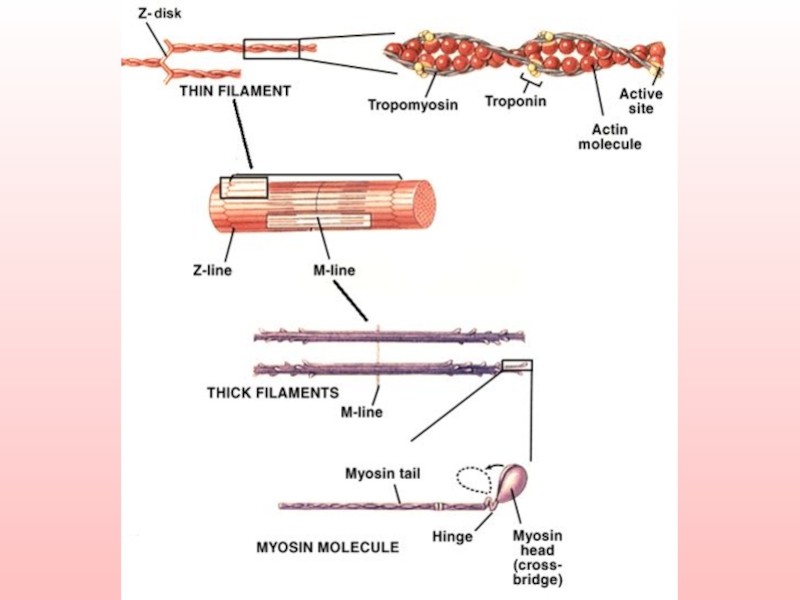
Слайд 15
В одной половине толстой протофибриллы (и в А-диске) все молекулы ориентированы
в одну сторону, а во второй половине - в противоположную
 все молекулы ориентированы в одну сторону, а")
.")

Слайд 18
В ходе сокращения миозин становится способным взаимодействовать с актином (нити актина
тянет к центру). При этом уменьшается длина каждого саркомера и мышцы в целом – происходит генерация силы.
Важно, что в процессе такого взаимного скольжения не меняется длина актиновых и миозиновых волокон.
Важно, что в процессе такого взаимного скольжения не меняется длина актиновых и миозиновых волокон.
. При")
Слайд 19
Схема сокращения
1 – актиновый филламент,
2 – центр связывания,
3
– миозиновый филламент,
4 – головка миозина,
5 – Z -диск саркомера.



Слайд 22Главной «моторной» частью миозина скелетных мышц является головка тяжелой цепи миозина.
Эти головки могут дотягиваться до нитей актина и контактировать с ними. При замыкании таких контактов образуются так называемые поперечные мостики, генерирующие тянущее усилие и обеспечивающие скольжение нитей актина относительно миозина.



Слайд 25
Предполагается, что поперечные мостики, отходящие от толстой нити прикрепляются к тонкой
нити под определенным углом, а затем, по-видимому, поворачиваются, что приводит к смещению толстых нитей относительно тонких.
Для того чтобы могло произойти значительное укорочение каждый поперечный мостик должен действовать циклически.
Для того чтобы могло произойти значительное укорочение каждый поперечный мостик должен действовать циклически.

Слайд 26Механизм сопряжения возбуждения и сокращения
1 – поперечная трубочка саркоплазматической мембраны,
2 –саркоплазматический ретикулум,
3 – ион Са++ ,
4 – молекула тропонина,
5 – молекула тропомиозина




Слайд 30
Поступление потенциала действия в моторные концевые пластинки аксона приводи к высвобождению
ацетилхолина из запасных пузырьков в синаптическую щель.
Ацетилхолиновые молекулы связываются с рецепторами в постсинаптических мембранах, что ведет к существенному увеличению проницаемости мембран и деполяризации мембран.
Потенциал действия, возникший в нейромышечном соединении быстро распространяется по поверхности волокна ( примерно 5 м в сек) и в поперечные трубочки.
Возбуждение поперечных трубочек высвобождает кальций, который в свою очередь запускает мышечное сокращение.
Ацетилхолиновые молекулы связываются с рецепторами в постсинаптических мембранах, что ведет к существенному увеличению проницаемости мембран и деполяризации мембран.
Потенциал действия, возникший в нейромышечном соединении быстро распространяется по поверхности волокна ( примерно 5 м в сек) и в поперечные трубочки.
Возбуждение поперечных трубочек высвобождает кальций, который в свою очередь запускает мышечное сокращение.

Слайд 31Нервный импульс передается поперечными трубочкам, где он трансформируется во взаимодействия между
дигидропиридиновыми и рианодиновыми рецепторами, что ведет к высвобождению кальция из саркоплазматического ретикулума (SR). Увеличение кальция запускает движение сократительных филамент
Схематическое изображение структур, связанных с возбуждением-контракцией мышечного волокна.

Слайд 32
Расслабление мышцы вызывается обратным переносом ионов Са2+ посредством кальциевого насоса в
каналы саркоплазматического ретикулума.
По мере удаления Са2+ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы
По мере удаления Са2+ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы

Слайд 33
При одиночном мышечном сокращении различают 3 фазы:
латентный период продолжительностью
около 5 мс – промежуток между воздействием стимула и началом видимого укорочения мышцы;
период сокращения продолжительностью 40 мс, в течении которого мышца укорачивается и производит работу;
период расслабления – самый длительный период (50 мс) во время которого мышца возвращается к своей исходной длине.
период сокращения продолжительностью 40 мс, в течении которого мышца укорачивается и производит работу;
период расслабления – самый длительный период (50 мс) во время которого мышца возвращается к своей исходной длине.

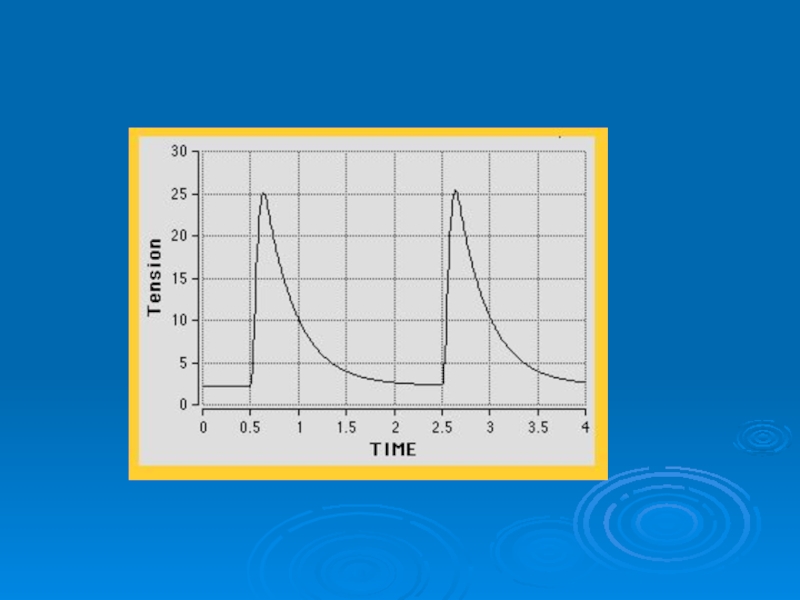

Слайд 36Одиночное сокращение и тетанус
При воздействии импульсов, наносимых с частотой более 20
в 1 с, в здоровой мышце в результате суммации нервно-мышечным аппаратом отдельных возбуждений происходит тетаническое сокращение, при котором полное расслабление между последовательными одиночными сокращениями произойти не успевает. Форма тетанического сокращения зависит от частоты импульсов.

Слайд 37Если период следования импульсов больше продолжительности одиночного сокращения (100 мс), то
возникает серия одиночных мышечных сокращений (а). Если же период будет меньше, сокращение приобретает форму зубчатого тетануса (b). При дальнейшем уменьшении периода следования импульсов возникают непрерывные сокращения и развивается гладкий тетанус ©.
, то возникает серия одиночных мышечных")
Слайд 38Used by permission of John W. Kimball
Н.Е.Введенский проводя опыты на
нервно-мышечном препарате лягушки установил, что нарастание до некоторого предела частоты или силы раздражений усиливает тетанус (т.н. оптимум). При дальнейшем увеличении частоты или усилении силы раздражения наступает резкое расслабление мышцы (пессимум).


Слайд 40Происхождение зубчатого и гладкого тетануса. Б – фазы и периоды мышечного сокращения, Б
– режимы мышечного сокращения, возникающие при разной частоте стимуляции мышцы.
Изменение длины мышцы показано синим цветом, потенциал действия в мышце - красным, возбудимость мышцы - фиолетовым







