- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Регуляция слияния мембран. Везикулярный транспорт (лекция 4) презентация
Содержание
- 1. Регуляция слияния мембран. Везикулярный транспорт (лекция 4)
- 3. NSF- гексамер (6 x
- 4. SNAREs – 18-47 кДа Гетерогенная группа мембранных
- 5. Исходная гипотеза Слияние происходит за счет
- 6. Но: Скорость слияния мембран гораздо выше, чем
- 7. Структура SNARE Белковые мембранные участки Остатки жирных кислот мембрана цитозоль
- 8. SNARE- комплекс наиболее стабилен, если образован «пучком»
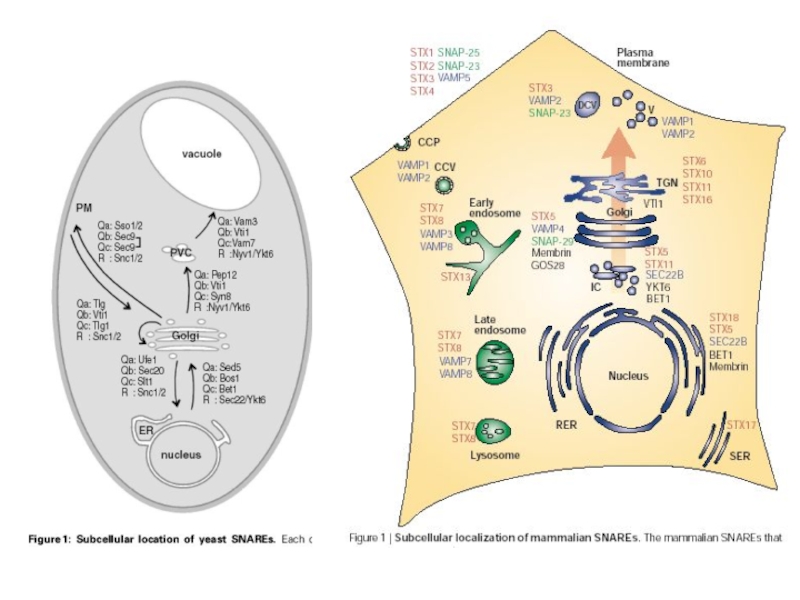
- 9. Как правило: R – v-SNARE, Q - t-SNARE Примеры SNARE-комплексов
- 10. Q R
- 12. Таким образом, специфичность слияний поддерживается за
- 13. Еще один уровень контроля слияний мембран: Способность
- 14. Действующая модель регуляции слияния с помощью SNARE-комплексов
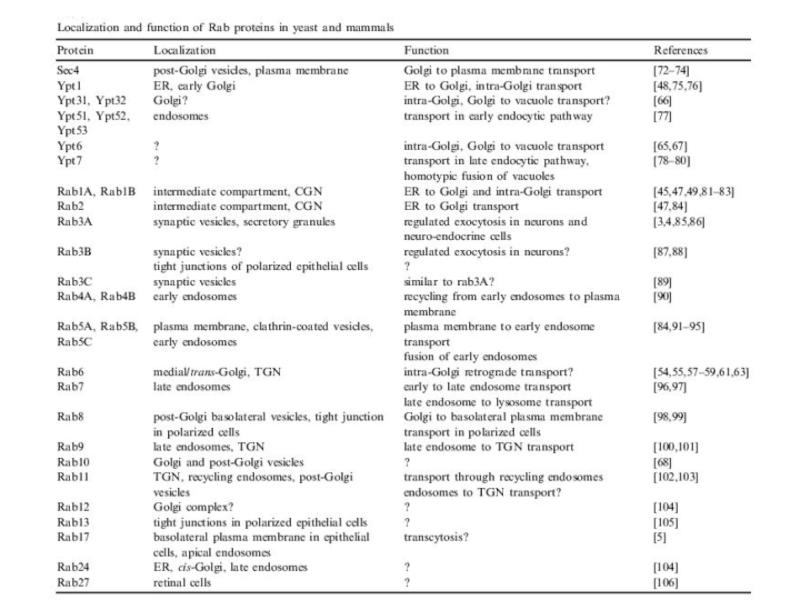
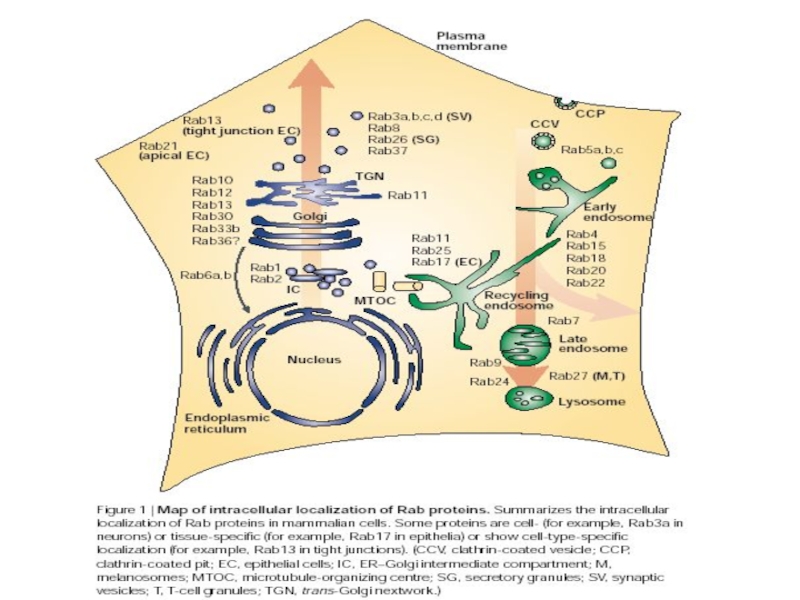
- 15. Rab-белки (Ras-related proteins from brain) Rab-белки,
- 16. Rab-белки (Ras-related proteins from brain) Малые ГТФазы
- 17. Каждый Rab-белок может иметь более одного GEF
- 20. Первый цикл Rab-белка после синтеза
- 21. Рабочий цикл Rab-белка Гидролиз ГТФ не нужен для слияния мембран
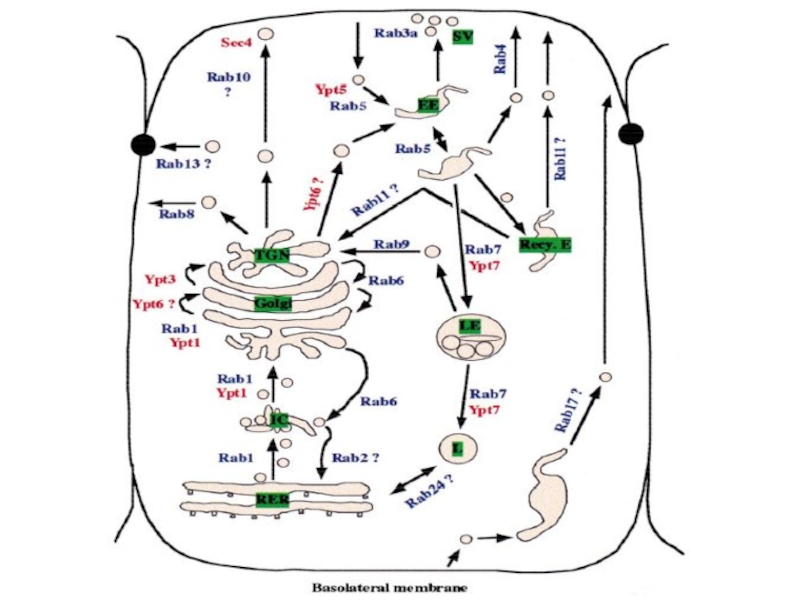
- 22. Но как же конкретно они
- 23. Множество партнеров Rab-белков представляют собой комплексы белков,
- 24. Секреция: Sec4 и Exocyst Слияние секреторных
- 25. Заякоривание СОРI-везикул (транспорт между цистернами Гольджи): Rab1- GM130/p115 Rab1
- 26. Гомотипичекое слияние ранних эндосом: Rab5 – EEA1 (early endosome’s autoantigene 1)
- 27. Эти белки или белковые комплексы получили
- 28. Rab-белки и/или их факторы дистанционного взаимодействия
- 29. Роль Rab-белков не ограничивается регуляцией слияния. 1.
- 30. 2. Rab-белки взаимодействуют с цитоскелетом в процессе
- 31. 3. В процессе заякоривания Rab-белок (GTP-связанный) способен
- 32. Rab-белки: Участвуют в формировании транспортной везикулы (за
- 34. Rab-белки в дрожжах:
- 35. (docking) ? Слияние происходит
- 36. Для полного слияния необходима реорганизация липидных бислоев
- 37. В качестве «носителя» для реорганизации липидных бислоев может выступать Vo-субъединица везикулярной протонной помпы V1/ V0
- 38. Не все слияния в клетке опосредуются NSF-SNAP-SNARE-системой:
- 40. Ca2+ и слияние мембран
- 41. Кальций участвует в регуляции слияния
- 42. BAPTA и EGTA – «быстрый» и «медленный»
- 43. Тотальное повышение уровня кальция Локальное повышение уровня
- 44. В синапсах: Синаптотагмин – имеет 2
- 45. 2. На эндосомах: HRS – (компонент
- 46. 3. На биосинетическом пути компонентами транспортной машинерии,
- 47. Еще одна функция Rab-белков – организация функциональных доменов на мембране органеллы
- 48. Эффекторы Rab5 – Рабаптин5, рабаптин5а, -
- 49. Множество эндосомных Rab-белков, собранных в субдомены,
- 51. 1


Слайд 3NSF- гексамер (6 x 75 кДа)
Является АТФазой, при
NSF-related proteins:
P97 – постмитотическая сборка аппарата Гольджи; не требует SNAP (in vitro)
Cdc48 – гомотипическое слияние при сборке ER и ядерной оболочки
Является АТФазой, при гидролизе АТФ «разворачивается»NSF-related proteins:P97 – постмитотическая")
Слайд 4SNAREs – 18-47 кДа
Гетерогенная группа мембранных белков,
способных образовывать высокостабильные
компартмент-специфические
Заякорены в мембране с помощью коротких хвостов (белковых или изопренильных)
t-SNARE
первоначальная классификация:
v-SNAREs - VAMP (vesicle-attached membrane proteins)
t-SNAREs - syntaxins;
SNAP25

Слайд 5Исходная гипотеза
Слияние происходит
за счет гидролиза АТФ на NSF,
энергия которого нужна
(priming) связи t- и v-SNARE
“20S fusion particle”
 связи")
Слайд 6Но:
Скорость слияния мембран гораздо выше, чем скорость гидролиза АТФ на NSF
Но:
1. некоторые стадии обслуживаются более чем 2-мя SNARE
2. некоторые SNARE обеспечивают более чем одну стадию транспортного процесса
Как же обеспечивается
специфичность слияния?


Слайд 8SNARE- комплекс наиболее стабилен, если образован «пучком» из 4 суперскрученных участков
Q-R-гипотеза: R+3Q



Слайд 12
Таким образом, специфичность слияний поддерживается за счет того, что каждой стадии
Например, Sed5p/Syn5 входит в состав всех комплексов, регулирующих слияния на биосинтетическом пути:
Yeast
Mammals
Yeast
Mammals
Yeast

Слайд 13Еще один уровень контроля слияний мембран:
Способность синтаксинов к слиянию подавлена в
n-Sec1 – t-SNARE протектор; подавляет скорость слияния в 2000 раз; вз-ет с N-концом синтаксинов

Слайд 14Действующая модель регуляции слияния с помощью SNARE-комплексов
Энергия гидролиза АТФ на NSF
Рециклирование v-SNAREs
hairpin

Слайд 15Rab-белки (Ras-related proteins from brain)
Rab-белки, несомненно, участвуют в регуляции слияния мембран:
ГДФ-связанные их формы блокируют транспорт, а ГТФ-γS-связанные –стимулируют образование сильно увеличенных компарментов
Rab-белки, несомненно, участвуют в регуляции слияния мембран: ГДФ-связанные их формы")
Слайд 16Rab-белки (Ras-related proteins from brain)
Малые ГТФазы
Компартмент-специфичны:
НО: большинство компартментов имеют более чем
Rab- белок
Идентифицировано ок. 60 генов у млекопитающих,
11 – в дрожжах
высококонсервативны
Малые ГТФазыКомпартмент-специфичны:НО: большинство компартментов имеют более чем один Rab- белок Идентифицировано")
Слайд 17Каждый Rab-белок может иметь более одного GEF или GAP, что позволяет

Слайд 20Первый цикл Rab-белка после синтеза
Rab
REP (Rab escort protein),
субъединица геранил-геранил-трансферазы
α-
мембрана
, субъединица геранил-геранил-трансферазыα- и β-субъединицы геранил-геранил-трансферазымембрана")

Слайд 22
Но как же конкретно они работают?
Ответ был получен по мере
данных по идентификации белков,
способных взаимодействовать с
активированными Rab-белками

Слайд 23Множество партнеров Rab-белков представляют собой комплексы белков, выстроенные линейно, или белки,

Слайд 24Секреция: Sec4 и Exocyst
Слияние секреторных пузырьков с ПМ происходит только

:Rab1- GM130/p115Rab1")
")
Слайд 27 Эти белки или белковые комплексы получили название
tethers (от «привязь», «коновязь»)
Работают как арканы, осуществляя заякоривание (tethering) мембраны на первой стадии слияния, когда расстояние между пузырьком и мишенью еще велико (около 25 нм) – т.е. являются факторами дистанционного взаимодействия
Стабилизируют мембраны для дальнейшего сближения (< 10 нм)
 Работают как арканы, осуществляя")
Слайд 28
Rab-белки и/или их факторы дистанционного взаимодействия могут узнавать соответствующие SNARE, тем

Слайд 29Роль Rab-белков не ограничивается регуляцией слияния.
1. Rab-белки, как правило, встраиваются в
Но ассоциация активированного Rab-белка может происходить и позже, уже после отделения транспортной везикулы от донорной мембраны (Rab5)
ПМ

Слайд 302. Rab-белки взаимодействуют с цитоскелетом в процессе передвижения транспортной везикулы
Rab27a-GTP
Rab27a-GDP
(or
МТ
МТ")
Слайд 313. В процессе заякоривания Rab-белок (GTP-связанный) способен снимать протекторный белок n-Sec1
 способен снимать протекторный белок n-Sec1 с t-SNARE и активировать")
Слайд 32Rab-белки:
Участвуют в формировании транспортной везикулы (за счет взаимодействия со SNARE, окаймлениями
Участвуют в ее перемещении к мембране-мишени (за счет взаимодействия с цитоскелетом)
Опосредуют 1-ю фазу слияния, стабилизируя везикулу напротив мембраны-мишени
Активируют t-SNARE, удаляя протекторный белок, могут взаимодействовать со SNARE (как v-, так и t-)
т.е. только 3+4 – участие в регуляции слияния
Участвуют в ее")

Слайд 35(docking)
?
Слияние происходит в 3 фазы:
Tethering (> 20 nm), Rabs
Docking (
3. lipid bilayers reorganization
?Слияние происходит в 3 фазы:Tethering (> 20 nm), RabsDocking (")
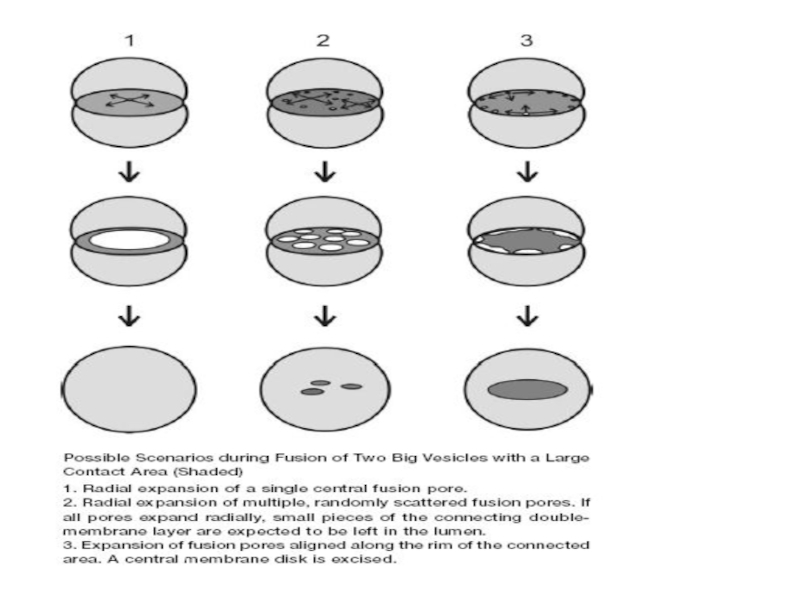
Слайд 36Для полного слияния необходима реорганизация липидных бислоев обеих мембран
Это промежуточное состояние,
flickering

Слайд 37В качестве «носителя» для реорганизации липидных бислоев может выступать Vo-субъединица везикулярной

Слайд 38Не все слияния в клетке опосредуются NSF-SNAP-SNARE-системой:
Пероксисомы и митохондрии не имеют
Вирусы имеют свою машинерию для образования пор слияния
, но")
Слайд 40
Ca2+ и слияние мембран
ЭПР
мит
Существует множество ионных каналов (как регулируемых, так и
В результате даже массированный вход Са2+ может быть быстро нивелирован за счет его выброса в окружающую среду и обратной закачки в депо.
 и транспортеров,")
Слайд 41Кальций участвует в регуляции
слияния синаптических пузырьков с пресинаптической мембраной
слияния
гомотипического слиянии СОРII-везикул
слияния эндосом друг с другом и с лизосомами
Сложилось представление о том, что все процессы слияния являются кальций-зависимыми
Однако, такого механизма до сих пор не обнаружено

Слайд 42BAPTA и EGTA – «быстрый» и «медленный» хелаторы кальция
в концентрации 10
EGTA - за 1,2 миллисекунды
быстрый хелатор подавляет слияния эндосом и COPII-везикул, тогда как медленный – нет
для эффективного слияния повышение уровня Са2+ требуется на время меньше 1 миллисекунды.
источник кальция (т.е. открытый канал) должен находиться на расстоянии порядка 20 нм от объекта влияния
Слияния идут за счет локального повышения уровня кальция
объект влияния - машинерия, связанная со слиянием;

Слайд 43Тотальное повышение уровня кальция
Локальное повышение уровня могут обеспечивать сами везикулы благодаря
При действии ростовых факторов, запускающих механизм опустошения внутриклеточных депо, «все должно слиться со всем», чего не происходит в действительности

Слайд 44В синапсах:
Синаптотагмин –
имеет 2 кальций-связывающих С2-домена,
стабилизирует SNARE-комплекс до момента
после чего связывает Са2+ и изменяет свою конформацию таким образом, что частично погружается в мембрану, способствуя быстрой реорганизации липидов.
Сам он после этого диссоциирует, что и прекращает дальнейшие слияния.
Позитивная регуляция слияния
Мишени и механизмы действия кальция?
Гипотеза: докинг комплекса SNARE стимулирует выброс Са2+, а он, в свою очередь, активирует некие Са2+-связывающие белки, которые и регулируют слияние.

Слайд 452. На эндосомах:
HRS – (компонент сортирующего комплекса ESCRT0, необходимого для направления
напрямую взаимодействует с Q-SNARE SNAP-25 за счет своего SNARE-подобного домена, препятствуя формированию комплекса слияния с синтаксином-13 и подавляя связывание c R-SNARE VAMP2.
Это ингибиторное взаимодействие существует до тех пор, пока не происходит выброс кальция из эндосом, в результате чего HRS высвобождается из комплекса, освобождая путь для SNARE-опосредуемого слияния.
Время выброса Са2+ из эндосом, в свою очередь, определяется моментом достижения определенного уровня рН (6,2 – 6,7) за счет работы вакуолярной протонной помпы и осуществляется через рН-чувствительный кальциевый канал
Таким образом, в данном случае кальциевый сенсор скорее является протектором несанкционированного слияния, которое могло бы произойти и в отсутствие кальция.
Негативная регуляция слияния
 напрямую")
Слайд 463. На биосинетическом пути компонентами транспортной машинерии, зависимой от Са2+, оказались
Связывание кальция стабилизирует эти окаймления.
Как это может повлиять на слияния?
Во-первых, стабилизация окаймлений может способствовать формированию транспортных пузырьков или препятствовать их обратным слияниям.
Во-вторых, даже на стадии взаимодействия с ERGIC везикулы могут сохранять часть окаймления, однако и в этом случае нельзя сказать, ингибиторное или стимулирующее действие на слияния будет оказывать Са2+.


Слайд 48Эффекторы Rab5 –
Рабаптин5, рабаптин5а, - стабилизируют связь Rab5 с Rabex5,
RBD5
Таким образом рабаптин 5 может регулировать гимотипическое слияние ранних эндосом
ЕЕ
ЕЕ

Слайд 49
Множество эндосомных Rab-белков, собранных в субдомены, отражает многообразие сортирующих функций эндосом,

Слайд 50
Rab5*
Rab4*
Rab7*
EE
EE
Гомотипическое слияние
Взаимодействие с цитоскелетом
Сортировка на путь рециклирования
Сортировка на путь деградации
МТ