д.м.н., проф. Марочков А.В.
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Разновидности мышечного волокна. Сократительные белки. Механизм сокращения презентация
Содержание
- 1. Разновидности мышечного волокна. Сократительные белки. Механизм сокращения
- 2. Типы и свойства мышечной ткани Возбудимость Сократимость Проводимость Эластичность
- 3. Макроскопическое строение мышц Брюшко Головка мышцы,
- 4. МЫШЦЫ
- 5. Микроскопическое строение мышц. Мышечный пучок Под какой
- 6. Схема строения мышечного волокна Саркомер - с
- 8. Саркомер в расслабленном состоянии Саркомер в состоянии сокращения
- 9. Саркоплазма В саркоплазме находится весь набор типичных
- 10. Актиновые миофиламенты Актиновые филаменты, скомпанованы из двух
- 11. Схема строения актиновых и миозиновых филаментов Миозиновые
- 13. миозин головка шейка
- 14. Актино-миозиновые мостики
- 15. Цикл работы головки миозина
- 16. Нервно-мышечный синапс 1 - пресинаптическая мембрана,
- 19. Строение нервно-мышечного синапса в разрезе
- 20. Электрохимическое преобразование ПД по мембране + дегидропиридина
- 21. Без ПД кальций в цитоплазму не выйдет!!
- 22. Механизм расслабления мышцы [Са+2] 107 М +
- 24. Затраты энергии АТФ во время мышечного сокращения
- 25. Двигательная или моторная единица Каждое мышечное волокно имеет только один синапс
- 26. Двигательная или моторная единица Мотонейрон и группа
- 27. Строение двигательной единицы
- 28. Распределение синапсов аксона мотонейрона в скелетной мышце
- 29. Классификация мышечных волокон
- 30. Медленные фазические волокна окислительного типа большое
- 31. Быстрые фазические волокна окислительного типа Содержат много
- 32. Быстрые фазические с гликолитическим типом окисления Мало
- 33. Тонические волокна Двигательный аксон образует множество синапсов
- 34. Закон «все или ничего» Одиночное мышечное волокно
- 35. Закон «все или ничего» Целая мышца данному
- 36. Режимы сокращения Изометрический – увеличение напряжения без
- 37. Виды мышечных сокращений Одиночное Тетанус Тонус
- 38. Виды раздражения мышцы в эксперименте Непрямое раздражение
- 39. Одиночное мышечное сокращение
- 40. Одиночное мышечное сокращение
- 41. Тетанус Это сильное и длительное сокращение
- 42. Суммация одиночных сокращений
- 43. Зубчатый тетанус Возникает в условиях когда каждый
- 46. Гладкий тетанус Возникает в условиях когда каждый
- 49. оптимум Частота раздражения, при которой наблюдается суммарное
- 50. пессимум Частота раздражения, при которй не наблюдается
- 52. 1 2 3 4 5 Е0 Екр 0 +30
- 53. 1 2 3 4 5
- 54. Сила мышц Зависит от толщины мышцы и её поперечного физиологического сечения
- 55. Работа мышцы Это энергия, затрачиваемая на перемещение
- 56. утомление Процесс временного снижения работоспособности мышцы. Возникает
- 58. Утомление в организме Утомление развивается вначале в
- 59. Гладкие мышцы
- 60. Висцеральные гладкие мышцы Все внутренние органы Большое количество нексусов (красные) Мало нервных окончаний (зеленые)
- 61. Мультиунитарные гладкие мышцы Представлены ресничной и мышцей
- 62. Иннервация ГМК
- 63. Особенности ГМК Веретенообразные клетки, тесно прилежат друг
- 64. Иннервация ГМК Иннервируют симпатические и парасимпатические нервные
- 65. Адекватные раздражители Нервный импульс Механическое растяжение Химические вещества
- 66. Особенности возбудимости Порог возбудимости ниже, чем у
- 67. Автоматия Способность клетки самостоятельно без внешнего раздражителя
- 68. Электрохимическое сопряжение в ГМК ПД открывает кальциевые
- 69. Механизм регуляции содержания кальция в ГМК
- 70. Особенности механизма сокращения в ГМК Кальций в
- 71. Спасибо за внимание!
Слайд 1Разновидности мышечного волокна. Сократительные белки. Анатомическое строение и физиологические свойства мышечной
ткани. Механизм сокращения.


Слайд 3Макроскопическое строение мышц
Брюшко
Головка мышцы,
Хвост мышцы,
Мышцы-антагонисты,
Мышцы-синергисты,
Апоневроз?

Слайд 4
МЫШЦЫ
СТРОЕНИЕ
КЛАССИФИКАЦИЯ
БРЮШКО
ФАСЦИЯ
ПОВЕРХНОСТНАЯ
ГЛУБОКАЯ
МЫШЕЧНЫЕ ПУЧКИ
ВСПОМОГА-ТЕЛЬНЫЙ АПАРАТ
СЫНОВИАЛЬНАЯ СУМКА
СЕСАМОВИДНЫЕ КОСТИ
ЗА
ФОРМОЙ
КВАДРАТНАЯ, ТРИУГОЛЬНАЯ, КОЛОВАЯ, ОДНОПЕРИСТАЯ, ДВОПЕРИСТАЯ:
ЗА КОЛИЧЕСТВОМ ГОЛОВОК
Одно-, дво-, три-, четырехглавая
ЗА НАПРАВЛЕНИЕМ ВОЛОКОН
Прямая, косая, поперечная, продольная
ЗА РОСПОЛОЖЕНИЕМ
Повекрхностные, глубокие, латеральные, медиальные
КВАДРАТНАЯ, ТРИУГОЛЬНАЯ, КОЛОВАЯ, ОДНОПЕРИСТАЯ, ДВОПЕРИСТАЯ:
ЗА КОЛИЧЕСТВОМ ГОЛОВОК
Одно-, дво-, три-, четырехглавая
ЗА НАПРАВЛЕНИЕМ ВОЛОКОН
Прямая, косая, поперечная, продольная
ЗА РОСПОЛОЖЕНИЕМ
Повекрхностные, глубокие, латеральные, медиальные

Слайд 5Микроскопическое строение мышц. Мышечный пучок
Под какой буквой обозначены гладкая и поперечнополосатая
мускулатура? А-; Б-.
Что обозначено цифрами
1-; 2-; 3-; 4-.
Что обозначено цифрами
1-; 2-; 3-; 4-.

Слайд 6Схема строения мышечного волокна
Саркомер - с двух сторон ограничен
Z – мембранами.
Толстые – миозиновые,
Тонкие – актиновые нити.
Состояния:
1 - расслабленное,
2 – сокращенное.
Длина саркомера в покоящейся мышце около
2 мкм, а в сократившейся c максимальной силой - несколько более 1 мкм.
Толстые – миозиновые,
Тонкие – актиновые нити.
Состояния:
1 - расслабленное,
2 – сокращенное.
Длина саркомера в покоящейся мышце около
2 мкм, а в сократившейся c максимальной силой - несколько более 1 мкм.

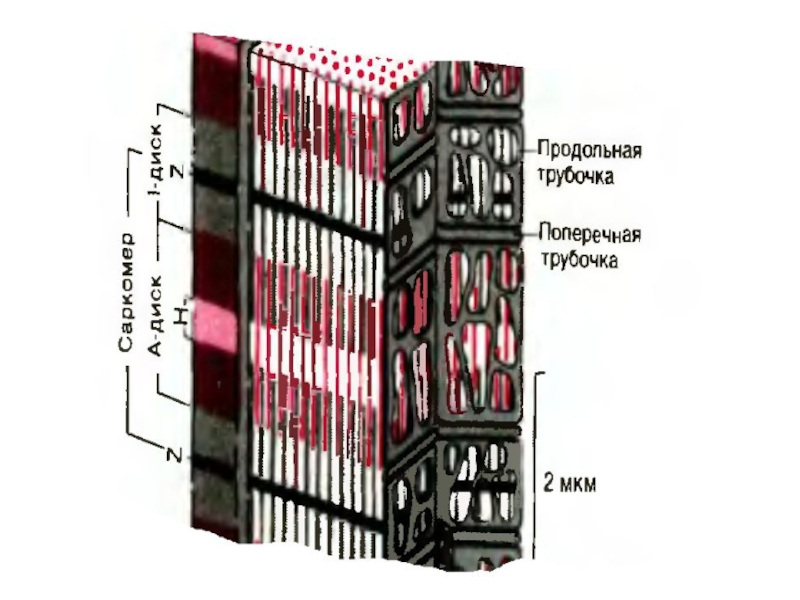

Слайд 9Саркоплазма
В саркоплазме находится весь набор типичных для любой клетки органоидов.
Особо следует
подчеркнуть наличие:
- саркоплазматического ретикулума,
- миоглобина,
- большого количества митохондрий,
Кроме того здесь есть сократимые миофиламенты.
- саркоплазматического ретикулума,
- миоглобина,
- большого количества митохондрий,
Кроме того здесь есть сократимые миофиламенты.

Слайд 10Актиновые миофиламенты
Актиновые филаменты, скомпанованы из двух актиновых нитей, представляющих собой как
бы бусинки глобулярных молекул актина. Тонкие нити имеют активные центры, расположенные друг от друга на расстоянии 40 нм, к которым могут прикрепляться головки миозина. Кроме актина в тонких нитях имеются и другие белки - тропомиозин, тропонины (I, T, C). Тропониновый комплекс располагается над активными центрами, прикрывая их, что препятствует соединению актина с миозином.

Слайд 11Схема строения актиновых и миозиновых филаментов
Миозиновые филаменты образуются более чем двумястами
молекулами миозина. Каждая из них скручена попарно и имеет выступающий отросток, называемый головкой. Головки направлены под углом от центра в сторону тонких нитей (напоминают «ерш» для мытья посуды). В основании головки миозина имеется фермент АТФаза, а на самой головке располагаются легкие цепи и молекула АТФ.





Слайд 16Нервно-мышечный синапс
1 - пресинаптическая мембрана,
2 - пузырьки с ацетилхолином,
3
- митохондрии,
4 - синапттическая щель,
5 - постсинаптическая мембрана,
7 - миофибриллы.
Ширина синаптической щели 20-30 нм
4 - синапттическая щель,
5 - постсинаптическая мембрана,
7 - миофибриллы.
Ширина синаптической щели 20-30 нм

Слайд 17
Ах
Аксонная терминаль
Аксонная бляшка
Ацетилхолин
Пресинаптическая
мембрана
Постсинаптическая
мембрана
Синаптическая
щель
Рецептор
Хемозависимый
канал
Электроза-
висимый
канал



Слайд 20Электрохимическое преобразование
ПД по мембране
+ дегидропиридина
в триадах
+ рианодина
в СПР
Открытие каналов
для кальция
в
СПР
Выход кальция
в цитоплазму
[Са+2] повышается с 105 до 107 М




Слайд 24Затраты энергии АТФ во время мышечного сокращения
На поддержание ионной асимметрии (натрий-калиевй
насос)
Образование мостиков («приклеивание» миозиновых головок к нитям актина) расщепляется 1 молекула АТФ на образование 1-го мостика
На работу кальциевого насоса!
Образование мостиков («приклеивание» миозиновых головок к нитям актина) расщепляется 1 молекула АТФ на образование 1-го мостика
На работу кальциевого насоса!
Образование мостиков («приклеивание» миозиновых")

Слайд 26Двигательная или моторная единица
Мотонейрон и группа мышечных волокон, иннервируемых разветвлениями аксона
этого мотонейрона
Число мышечных волокон в единице зависит от функции, которую выполняет данная мышца (от 10 до 30 000)
Число мышечных волокон в единице зависит от функции, которую выполняет данная мышца (от 10 до 30 000)




Слайд 30Медленные фазические волокна окислительного типа
большое содержание миоглобина и митохондрий
красного
цвета
медленно утомляются
В одной моторной единице их очень много (до 30 000)
Входят в состав мышц, поддерживающих позу
медленно утомляются
В одной моторной единице их очень много (до 30 000)
Входят в состав мышц, поддерживающих позу

Слайд 31Быстрые фазические волокна окислительного типа
Содержат много митохондрий
Способны синтезировать АТФ путем окислительного
фосфорилирования
Выполняют быстрые сокращения
Утомляются медленно
В составе моторной единицы меньше, чем медленных
Выполняют быстрые сокращения
Утомляются медленно
В составе моторной единицы меньше, чем медленных

Слайд 32Быстрые фазические с гликолитическим типом окисления
Мало митохондрий
АТФ образуется за счет гликолиза
Миоглобина
нет (белый цвет)
Быстро сокращаются и быстро утомляются
В моторной единице небольшое количество волокон
Быстро сокращаются и быстро утомляются
В моторной единице небольшое количество волокон
Быстро сокращаются")
Слайд 33Тонические волокна
Двигательный аксон образует множество синапсов
Медленно сокращаются и медленно расслеабдяются
Низкая АТФ-азная
активность миозина
У человека входят в состав наружных мышц глаз
У человека входят в состав наружных мышц глаз

Слайд 34Закон «все или ничего»
Одиночное мышечное волокно подчиняется этому закону:
Подпороговое раздражение не
вызывает сокращение, а пороговое – вызвает максимально возможное сокращение, т.о. амплитуда мышечного сокращения не зависит от силы раздражения

Слайд 35Закон «все или ничего»
Целая мышца данному закону не подчиняется потому, что
состоит из множества моторных единиц, обладающих разным порогом деполяризации

Слайд 36Режимы сокращения
Изометрический – увеличение напряжения без изменения длины мышцы
Изотонические – уменьшение
длины мышцы без изменения её напряжения
Смешанный - ауксотонический
Смешанный - ауксотонический


Слайд 38Виды раздражения мышцы в эксперименте
Непрямое раздражение – импульс воздействует на нервное
окончание мотонейрона
Прямое раздражение – импульс воздействует непосредственно на мышцу
Прямое раздражение – импульс воздействует непосредственно на мышцу


Слайд 40Одиночное мышечное сокращение
1
2
3
1 – латентный период
2 – период напряжения
3 – период
расслабления

Слайд 41Тетанус
Это сильное и длительное сокращение мышцы в ответ на серию
раздражений.
Происходит за счет суммации одиночных сокращения в следствии увеличения концентрации кальция в цитоплазме
Происходит за счет суммации одиночных сокращения в следствии увеличения концентрации кальция в цитоплазме


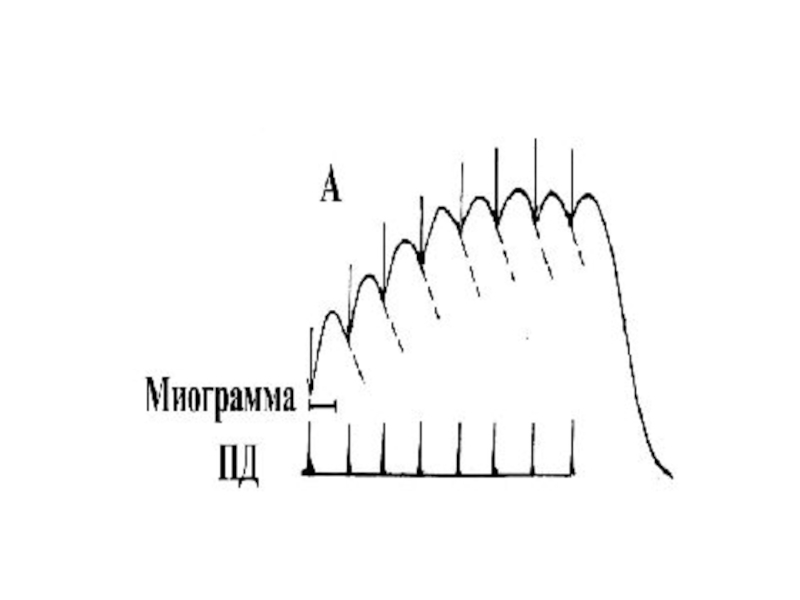
Слайд 43Зубчатый тетанус
Возникает в условиях когда каждый последующий импульс попадает в период
расслабления
или интервал между импульсами меньше чем длительность одиночного сокращения, но больше чем период укорочения
или интервал между импульсами меньше чем длительность одиночного сокращения, но больше чем период укорочения


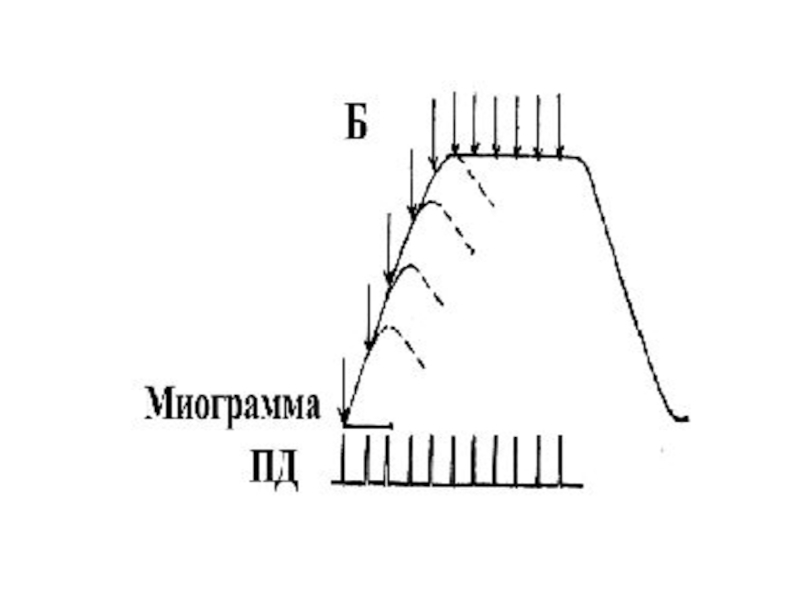
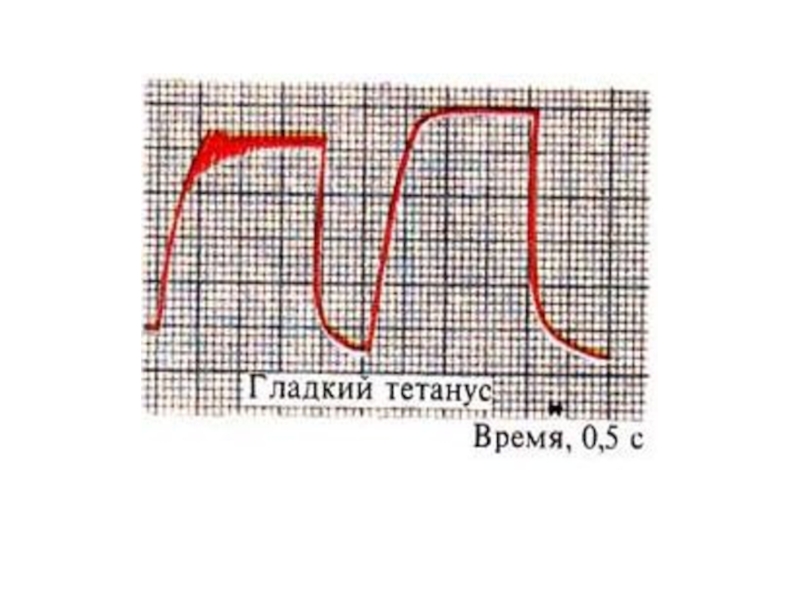
Слайд 46Гладкий тетанус
Возникает в условиях когда каждый последующий импульс попадает в период
укорочения
Или интервал между импульсами меньше чем длительность периода укорочения, но больше чем латентный период .
Или интервал между импульсами меньше чем длительность периода укорочения, но больше чем латентный период .

Слайд 49оптимум
Частота раздражения, при которой наблюдается суммарное сокращение (тетанус) наибольшей амплитуды
При этом
каждый последующий импульс попадает в период супернормальности, т.е. сразу после ПД
Это приводит к тому, что в цитоплазме поддерживается наибольшая концентрация кальция (насос не успевает включится)
Это приводит к тому, что в цитоплазме поддерживается наибольшая концентрация кальция (насос не успевает включится)
 наибольшей амплитудыПри этом каждый последующий импульс попадает")
Слайд 50пессимум
Частота раздражения, при которй не наблюдается суммации сокращения
При этом каждый
последующий импульс попадает в период рефрактерности (в период развития ПД)
В результате на мембране все каналы для натрия остаются инактивированными и невозможно возникновение нового ПД
В результате на мембране все каналы для натрия остаются инактивированными и невозможно возникновение нового ПД





Слайд 55Работа мышцы
Это энергия, затрачиваемая на перемещение тела с определенной силой на
определенное расстояние:
A = F x S
Если F=0, то и работа А=0
Если S=0, то и работа А=0
Максимальная работа совершается при средних нагрузках
Amax= Fср x Smax
A = F x S
Если F=0, то и работа А=0
Если S=0, то и работа А=0
Максимальная работа совершается при средних нагрузках
Amax= Fср x Smax

Слайд 56утомление
Процесс временного снижения работоспособности мышцы.
Возникает в связи с уменьшением энергетических запасов
(АТФ) в мышечном волокне или уменьшением медиатора в нервно-мышечном синапсе
В нервно-мышечном препарате утомление раньше развивается в синапсе!
В нервно-мышечном препарате утомление раньше развивается в синапсе!
 в мышечном волокне")
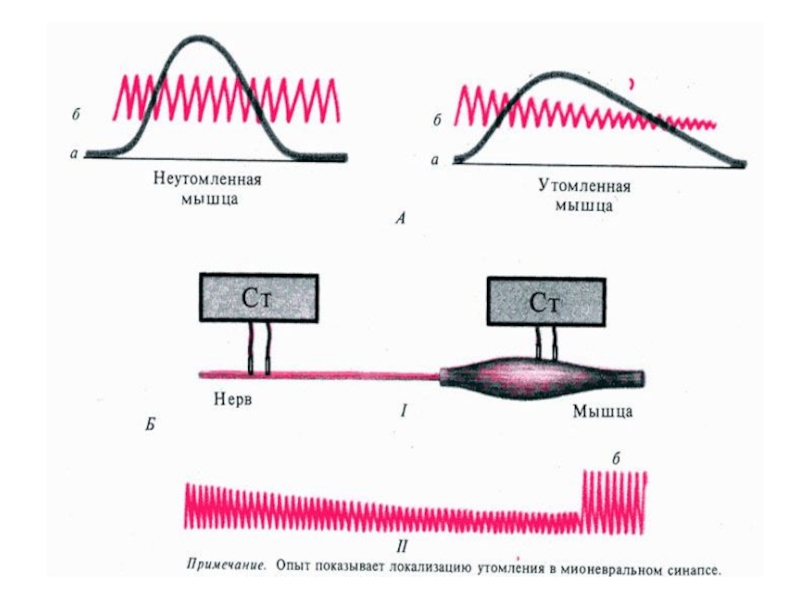
Слайд 58Утомление в организме
Утомление развивается вначале в нервных центрах – это защитный
механизм
При этом в мышцах остается «аварийный» запас энергии
Доказательство – опыты Сеченова И.М. с активным отдыхом
При этом в мышцах остается «аварийный» запас энергии
Доказательство – опыты Сеченова И.М. с активным отдыхом


Слайд 60Висцеральные гладкие мышцы
Все внутренние органы
Большое количество нексусов (красные)
Мало нервных окончаний (зеленые)
Мало нервных окончаний (зеленые)")
Слайд 61Мультиунитарные гладкие мышцы
Представлены ресничной и мышцей радужки глаза
Отмечается большая плотность нервных
синаптических (варикозных) контактов (зеленый цвет)
 контактов (зеленый цвет)")

Слайд 63Особенности ГМК
Веретенообразные клетки, тесно прилежат друг к другу
Между клетками – плотные
контакты – нексусы
Актин и миозин расположены неупорядоченно
СПР развит меньше, чем в скелетных
Актин и миозин расположены неупорядоченно
СПР развит меньше, чем в скелетных

Слайд 64Иннервация ГМК
Иннервируют симпатические и парасимпатические нервные волокна
Нет синапсов, нервные окончания образуют
варикозные расширения
Медиатор выделяется в межтканевую жидкость
Возбуждается 1 клетка и передает ПД по нексусам (электрические синапсы)
Вся мышца сокращаяется одномоментно (функциональный синцитий)
Медиатор выделяется в межтканевую жидкость
Возбуждается 1 клетка и передает ПД по нексусам (электрические синапсы)
Вся мышца сокращаяется одномоментно (функциональный синцитий)


Слайд 66Особенности возбудимости
Порог возбудимости ниже, чем у поперечно-полосатых мышц
Мембрана более проницаема для
натрия, поэтому МП меньше (-50, -60 мв)
Амплитуда ПД ниже, а длительность больше
Деполяризация открывает потенциал-зависимые кальциевые каналы, вход кальция замедляет реполяризацию
Амплитуда ПД ниже, а длительность больше
Деполяризация открывает потенциал-зависимые кальциевые каналы, вход кальция замедляет реполяризацию

Слайд 67Автоматия
Способность клетки самостоятельно без внешнего раздражителя генерировать ПД
ГМК в состоянии покоя
пропускает натрий, который медленно деполяризует клетку до критического уровня и возникает ПД

Слайд 68Электрохимическое сопряжение в ГМК
ПД открывает кальциевые каналы и в клетку входит
кальций.
Лиганд - активирует фосфолипазу С на мембране клетки – активация инозитолтрифосфата и диацилглицерола.
инозитолтрифосфат освобождает из цистерн кальций и он инициирует сокращение
Лиганд - активирует фосфолипазу С на мембране клетки – активация инозитолтрифосфата и диацилглицерола.
инозитолтрифосфат освобождает из цистерн кальций и он инициирует сокращение


Слайд 70Особенности механизма сокращения в ГМК
Кальций в цитоплазме связывается с кальмодулином и
активирует протеинкиназу .
Протеинкиназа фосфорилирует головку миозина
Образуются мостики между миозином и актином
Уменьшение концентрации кальция в миоплазме вызывает дефосфорилирование головки миозина – мостики распадаются
Мышца расслабляется
Протеинкиназа фосфорилирует головку миозина
Образуются мостики между миозином и актином
Уменьшение концентрации кальция в миоплазме вызывает дефосфорилирование головки миозина – мостики распадаются
Мышца расслабляется






