- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Общий план строения эукариотической клетки презентация
Содержание
- 1. Общий план строения эукариотической клетки
- 2. Источники информации О.В. Быков В.Л. Цитология и
- 3. Эукариотическая животная клетка
- 4. Немембранные
- 5. Схема строения цитолеммы. 1 - углеводные цепи
- 6. Морфофункциональные особенности клеточных мембран Все клеточные
- 7. Функции мембран в животной клетке
- 8. *Функции некоторых цитоплазматических и внутриклеточных мембран в животной клетке
- 9. Мембрана состоит из двойного слоя липидов и
- 10. Состав мембран
- 11. Химическая структура молекулы фосфолипида Холин Фосфат Глицерол
- 12. Липосома из одного биослоя липидов Липосома из бимолекулярых слоев СПОСОБЫ УПАКОВКИ ФОСФОЛИПИДОВ
- 13. МЕМБРАННЫЕ БЕЛКИ 1. Структурные белки: фибронектин, спектрин,
- 14. Схема переноса ионов и молекул через плазматическую
- 16. 3. Белки, обеспечивающие непосредственные межклеточные взаимодействия. 4. Белки, участвующие в передаче сигналов.
- 17. Эндоцитоз - процесс обволакивания внеклеточного материала плазмалеммой
- 18. Цитоплазма Переходы гель-золь характерны для гиалоплазмы.
- 19. Эндоплазматическая сеть Эндоплазматическая сеть –
- 20. Электронные микрофотографии участков клетки с мощно развитой
- 21. Синтетический аппарат клетки
- 22. Строение рибосомы. А – сборка субъединиц рибосом.
- 23. Полная прокариотическая рибосома имеет коэффициент седиментации 70S
- 24. Синтез белка на гранулярной ЭПС. РФ –
- 25. Модель динамической работы рибосомы в элонгационном цикле.
- 26. Пластинчатый комплекс или комплекс Гольджи Микрофотография клеток
- 27. Комплекс Гольджи представлен мембранными структурами, собранными вместе
- 28. По мере созревания белков в цистернах комплекса
- 29. Таким образом, в комплексе Гольджи происходит не
- 30. Лизосомы Схема образования лизосом 1 –
- 31. Митохондрии Впервые митохондрии были обнаружены Р.
- 32. На электронной микрофотографии панкреатической клетки крысы видна
- 33. Строение и работа митохондрий: а – митохондрии,
- 34. Каждый комплекс (I, III, IV) образован многими
- 35. Схема цикла трикарбоновых кислот - цикла Кребса

Слайд 2Источники информации
О.В. Быков В.Л. Цитология и общая гистология (функциональная морфология клеток
Леви А., Сикевиц Ф. Структура и функции клетки. – М.: Мир, 1971.
Меркулов Г.А. Курс патологогистологической техники. – Изд-во Медицина, Ленинградское отделение, 1969.
Ченцов Ю.С. Общая цитология. – М.: Изд-во МГУ, 1995
Ченцов Ю.С. Введение в клеточную биологию. – М.: ИКЦ «Академкнига», 2004
Щелкунов С.И. Клеточная теория и учение о тканях. – Медгиз, Ленинградское отделение, 1958.
Джеральд М. Фаллер, Денис Шилдс. Молекулярная биология клетки. Руководство для врачей. Пер. с англ. М.: «Издательство БИНОМ»2006 – 256 с., ил
А.Г. Гунин Атлас микрофотографий histol@mail.ru
. –")


Слайд 5Схема строения цитолеммы.
1 - углеводные цепи гликокаликса;
2 – гликолипид; 3
7 - молекула холестерина; 8 – микрофиламенты;
9 - микротрубочки
Клетки прокариот (кроме Микоплазм) имеют клеточную стенку из муреина или целлюлозы (Цианобактерии).
Поверхностный аппарат клетки
Все растительные клетки (кроме Диатомовых водорослей) имеют клеточную оболочку из целлюлозы, которая может быть пропитана различными веществами (например: лигнин, суберин…).
Клетки грибов тоже имеют клеточную оболочку, у многих грибов в её состав входит хитин.
Клетки животных клеточной стенки не имеют.
Все и прокариотические и эукариотические клетки имеют плазматическую мембрану - плазмалемму.

Слайд 6Морфофункциональные особенности клеточных мембран
Все клеточные мембраны представляют собой сложную смесь
1. Мембраны не однородны. Мембраны, окружающие внутриклеточные органеллы, и плазматическая мембрана отличаются по составу.
2. Многие компоненты мембран находятся в состоянии непрерывного движения. Мембрана напоминает постоянно меняющуюся мозаику. Некоторые части мембраны изменяются быстрее, чем другие.
3. Компоненты мембран чрезвычайно асимметричны. Между наружным и внутренним слоями мембран имеется различие по относительному количеству и качественному составу липидов. Белки располагаются среди липидов асимметрично и имеют хорошо различимые вне- и внутриклеточные домены.



Слайд 9Мембрана состоит из двойного слоя липидов и связанных с ним протеинов.
Жидкостно-мозаичная модель мембраны.
1 - интегральный белок; 2 - билипидный слой мембраны; 3 - гидрофильные головки фосфолипидов; 4 - гидрофобные хвосты фосфолипидов; 5 - периферический белок; 6 - рецепторная часть мембраны..
При раскалывании замороженной мембраны она расщепляется на два липидных слоя наименее прочного соединения, где контактируют гидрофобные части липидов.


Слайд 11Химическая структура молекулы фосфолипида
Холин
Фосфат
Глицерол
Образование липидных мембран, мицелл, эмульсий и липосом

Слайд 12Липосома из одного биослоя липидов
Липосома из бимолекулярых слоев
СПОСОБЫ УПАКОВКИ ФОСФОЛИПИДОВ

Слайд 13МЕМБРАННЫЕ БЕЛКИ
1. Структурные белки: фибронектин, спектрин, гликофорин.
В плазмалемме эритроцита около 360

Слайд 14Схема переноса ионов и молекул через плазматическую мембрану
1 – простая диффузия;
4 – белок переносчик
(K+ Na+) -насос
1 – участок связывания Na+;
2 – участок связывания K+;
3 – мембрана
Транспорт ионов через плазмалемму проходит за счет участия в этом процессе мембранных транспортных белков - пермеаз. Эти белки могут вести транспорт в одном направлении одного вещества (унипорт), или нескольких веществ одновременно (симпорт), или же вместе с импортом одного вещества выводить из клетки другое (антипорт). Так, например, глюкоза может входить в клетки симпортно вместе с ионом Na+.
В клетках существуют мембранные белковые переносчики, которые работают против градиента концентрации, затрачивая при этом энергию АТФ. Такой тип работы носит название активного транспорта, и он осуществляется с помощью белковых ионных насосов.
2. Транспортные белки. Межмембранный транспорт
В плазматической мембране находится двухсубъединичная молекула (K+ Na+)-насоса, которая одновременно является и АТФазой. Этот насос при работе откачивает за один цикл 3 иона Na+ и закачивает в клетку 2 иона K+ против градиента концентрации.

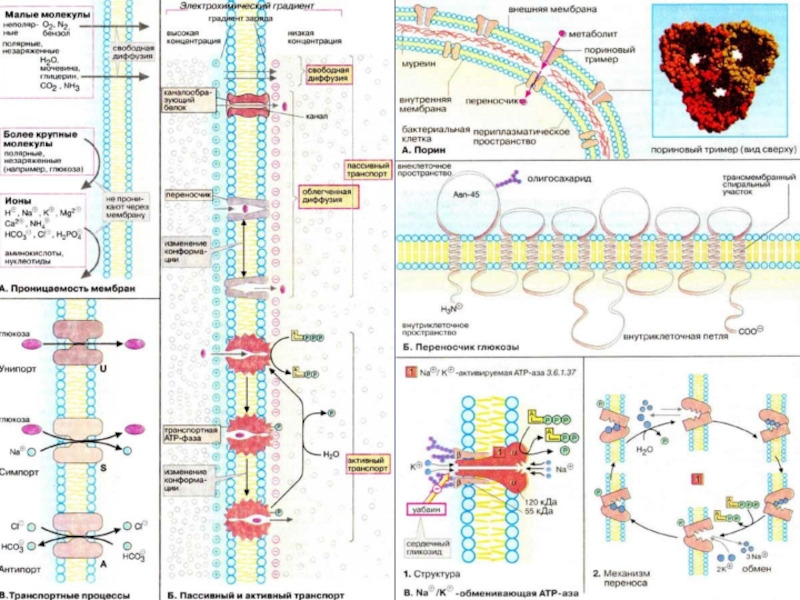
Слайд 163. Белки, обеспечивающие непосредственные межклеточные взаимодействия.
4. Белки, участвующие в передаче сигналов.

Слайд 17Эндоцитоз - процесс обволакивания внеклеточного материала плазмалеммой и заключение его в
Фагоцитоз, способность захватывать клеткой крупные частицы, встречается среди клеток животных как одноклеточных (например, амебы, некоторые хищные инфузории), так и для специализированных клеток многоклеточных животных - фагоцитов.
Эндоцитоз
Пиноцитоз вначале определялся как поглощение клеткой воды или водных растворов разных веществ. Сейчас известно, что как фагоцитоз, так и пиноцитоз протекают очень сходно, и поэтому употребление этих терминов может отражать лишь различия в объемах, массе поглощенных веществ. Общее для этих процессов то, что поглощенные вещества на поверхности плазматической мембраны окружаются мембраной в виде вакуоли - эндосомы, которая перемещается внутрь клетки.
Клатрин – сигнал и форма пузырьков

Слайд 18Цитоплазма
Переходы гель-золь характерны для гиалоплазмы.
В гиалоплазме могут возникать и
В гиалоплазме находятся ферменты, участвующие в синтезе аминокислот, нуклеотидов, жирных кислот, сахаров.
В гиалоплазме происходит синтез и отложение гликогена, накопление запасных жировых капель. Здесь же происходят процессы гликолиза и синтез части АТФ.

Слайд 19Эндоплазматическая сеть
Эндоплазматическая сеть – ЭПС (ретиклум ЭР) - сквозная сеть
Эта внутриклеточная мембранная структура была открыта в 1945 году К. Р. Портером.
В 50-х гг., при использовании метода ультратонких срезов удалось выяснить структуру эндоплазматического ретикулюма и обнаружить его неоднородность. Электронно-микроскопический анализ позволил выделить два типа ЭПС: гранулярную (шероховатую) и агранулярную или гладкую.
 - сквозная сеть двухконтурных мембран, соединенных друг")
Слайд 20Электронные микрофотографии участков клетки с мощно развитой системой каналов ЭПС. 1
Гранулярная ЭПС представлена замкнутыми мембранами, которые образуют на сечениях вытянутые мешки, цистерны или же имеют вид узких (около 20 нм) каналов. Отличительной особенностью гранулярной ЭПС, является то, что ее мембраны со стороны гиалоплазмы покрыты мелкими (около 20 нм) темными, почти округлыми частицами, гранулами - рибосомами.
На мембранах рибосомы расположены в виде полисом (множество рибосом, объединенных одной информационной РНК). Это синтезирующие белок рибосомы прикрепляются к мембранам большой субъединицей.
Важнейшей функцией гранулярной ЭПС, вне зависимости от специализации или тканевой принадлежности клеток, является функция образования и построения клеточных мембран.


Слайд 22Строение рибосомы.
А – сборка субъединиц рибосом. 1 – малая и 2
Рибосомы
Рибосома представляет собой элементарную клеточную машину синтеза любых белков клетки. Рибосомы в клетке многочисленны: за клеточный цикл их образуется 1х107. Рибосомы - это сложные рибонуклеопротеидные частицы, в состав которых входит множество молекул индивидуальных белков и несколько молекул РНК. Полная, работающая рибосома, состоит из двух неравных субъединиц: большой и малой. Размер полной прокариотической рибосомы составляет 20 х 17 х 17 нм, эукариотической - 25 х 20 х 20.

Слайд 23Полная прокариотическая рибосома имеет коэффициент седиментации 70S и диссоциирует на две

Слайд 24Синтез белка на гранулярной ЭПС. РФ – рибофовины, СРЧ – сигнал-распознающая

Слайд 25Модель динамической работы рибосомы в элонгационном цикле. Размыкание рибосомных субъединиц открывает
1 – связывание аминоацетил-тРНК с рибосомой и фактор элонгации открывает рибосому;
2 – фактор элонгации покидает рибосому, аминокислотный конец аминоацетил-тРНК взаимодействует с пептидилтрансферазным центром большой субъединицы рибосомы и способствует замыканию субъединиц; 3 – реакция транспептидации происходит в закрытой рибосоме; 4 – размыкание транслокационной рибосомы осуществляется вторым фактором элонгации, что приводит к выходу тРНК и смещению молекулы пептидил-тРНК вместе с мРНК; 5 – рибосома вновь смыкается.

Слайд 26Пластинчатый комплекс или комплекс Гольджи
Микрофотография клеток с выявленными каналами комплекса Гольджи
Электронограмма
В 1898 году итальянский ученый Камилло Гольджи, используя свойства связывания тяжелых металлов (осмия и серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал «внутренним сетчатым аппаратом». Обычно элементы комплекса Гольджи расположены около ядра, вблизи клеточного центра. Участки комплекса Гольджи, имеют в некоторых клетках вид сложных сетей, где ячейки связаны друг с другом или представлены в виде отдельных темных участков, лежащих независимо друг от друга, имеющих вид палочек, зерен, вогнутых дисков.
 пластинчатого комплекса Гольджи")
Слайд 27Комплекс Гольджи представлен мембранными структурами, собранными вместе в небольшой зоне. Отдельная
В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены в виде стопки плоские мембранные мешки, или цистерны. Каждая отдельная цистерна имеет диаметр около 1 мкм; на периферии цистерны имеют расширения, ампулы. Количество таких мешков в стопке обычно не превышает 5-10.
Кроме плотно расположенных плоских цистерн в зоне комплекса Гольджи наблюдается множество вакуолей. Мелкие вакуоли встречаются главным образом в периферических участках; иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн.
Принято различать в зоне диктиосомы проксимальный или формирующийся, цис-участок, и дистальный или зрелый, транс-участок. Между ними располагается средний или промежуточный участок комплекса Гольджи.
В секретирующих клетках обычно комплекс Гольджи поляризован: его проксимальная часть обращена к цитоплазме и ядру, а дистальная - к поверхности клетки.
Комплекс Гольджи является промежуточным звеном между собственно синтезом секретируемого белка и выведением его из клетки.

Слайд 28По мере созревания белков в цистернах комплекса Гольджи, они с помощью
Синтезированный на рибосомах экспортируемый белок отделяется и накапливается внутри цистерн ЭПС, по которым он транспортируется к зоне мембран комплекса Гольджи. Здесь от гладких участков ЭПС отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей в проксимальной части диктиосомы. В этом месте вакуоли могут сливаться друг с другом и с плоскими цис-цистернами диктиосомы. Происходит перенесение белкового продукта уже внутри полостей цистерн КГ.

Слайд 29Таким образом, в комплексе Гольджи происходит не просто «перекачка» продуктов из
1. Гликозелирование белков и липидов.
2. Гликозелирование и сборка протеогликанов.
3. Добавление манозо-6-фосфата к ферментам лизосом.
4. Сортировка для транспорта.
*Биохимические процессы в комплексе Гольджи.

Слайд 30Лизосомы
Схема образования лизосом
1 – комплекс Гольджи; 2 – первичная лизосома;
5 – пиноцитоз; 6 – остаточное тельце;
7 – слияние лизосомы с митохондрией;
8 - аутосома
Лизосомы – главные пищеварительные органеллы клетки были открыты Де Дюв в 1955году. Лизосомы представляют собой пузырьки, содержащие около 40 гидролитических ферментов: протеиназы, нуклеазы, гликозидазы, фосфорилазы, фосфатазы, сульфитазы и др. Под электронным микроскопом видно, что лизосомы представлены пузырьками (0,2-0,4 мкм), ограниченными одиночной мембраной, с очень разнородным содержимым внутри. Среди лизосом можно выделить четыре типа: первичные лизосомы, вторичные лизосомы, аутофагосомы и остаточные тельца.
Первичные лизосомы - пузырьки размером около 100 нм, которые трудно отличить от мелких вакуолей комплекса Гольджи. Первичные лизосомы сливаются с фагоцитарными или пиноцитозными вакуолями и образуют вторичную лизосому или внутриклеточную пищеварительную вакуоль. Содержимое первичной лизосомы сливается с полостью эндоцитозной вакуоли, и гидролазы начинают расщеплять полимеры. Расщепление макромолекул внутри лизосом может идти не до конца - в полостях лизосом происходит накопление непереваренных продуктов, и вторичные лизосомы превращаются в телолизосомы, или остаточные тельца.
Аутолизосомы содержат фрагменты или даже целые цитоплазматические структуры, такие, как митохондрии, пластиды, элементы ЭПС, рибосомы, гранулы гликогена.

Слайд 31Митохондрии
Впервые митохондрии были обнаружены Р. Альтманом, который описал эти клеточные
Внутренняя мембрана ограничивает собственно внутреннее содержимое митохондрии, ее матрикс. Характерной чертой внутренней мембраны митохондрий является их способность образовывать многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид плоских гребней, или крист.
Митохондрии ограничены двумя мембранами. Наружная митохондриальная мембрана отделяет ее от гиалоплазмы. Наружную мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм.
Схема строения митохондрии.
1 – наружная мембрана; 2 – внутренняя мембрана; 3- кристы;
4 – матрикс; 5 – митохондриальные рибосомы

Слайд 32На электронной микрофотографии панкреатической клетки крысы видна целая митохондрия. Ясно видны
Различные формы хондриома: а – разрозненные митохондрии, б - группа митохондрий в клетках сетчатки, в - митохондрильный ретикулум одноклеточной водоросли хлореллы
Матрикс митохондрий имеет тонкозернистое гомогенное строение, в нем иногда выявляются тонкие собранные в клубок нити (около 2-3 нм) и гранулы около 15-20нм. Теперь стало известно, что нити матрикса митохондрий представ-ляют собой молекулы ДНК в составе митохондриального нуклеоида, а мелкие гранулы - митохондриальные рибосомы.
Хондриом – это совокупность всех митохондрий в одной клетке.

Слайд 33Строение и работа митохондрий: а – митохондрии, б- ультраструктура митохондрий, общая

Слайд 34Каждый комплекс (I, III, IV) образован многими белками. Белок цитохром с
Структурная организация компонентов дыхательной цепи и схема связывания энергии в молекулах АТФ на мембранах митохондрий.
 образован многими белками. Белок цитохром с (обозначено с) в окисленном")






