- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Мейоз. Первое деление мейоза презентация
Содержание
- 1. Мейоз. Первое деление мейоза
- 2. Мейоз Способ деления эукариот, при котором образуются
- 3. Первое деление мейоза
- 4. Мейоз - первое деление Интерфаза 1.
- 5. Мейоз - первое деление Профаза 1.
- 6. Профаза I Несестринские хроматиды гомологичных хромосом бивалента
- 7. Мейоз Не для каждой хромосомы можно обнаружить
- 8. Профаза I В первой профазе мейоза принято
- 9. Мейоз - первое деление Характерной особенностью профазы
- 10. Мейоз - первое деление Метафаза 1. Биваленты
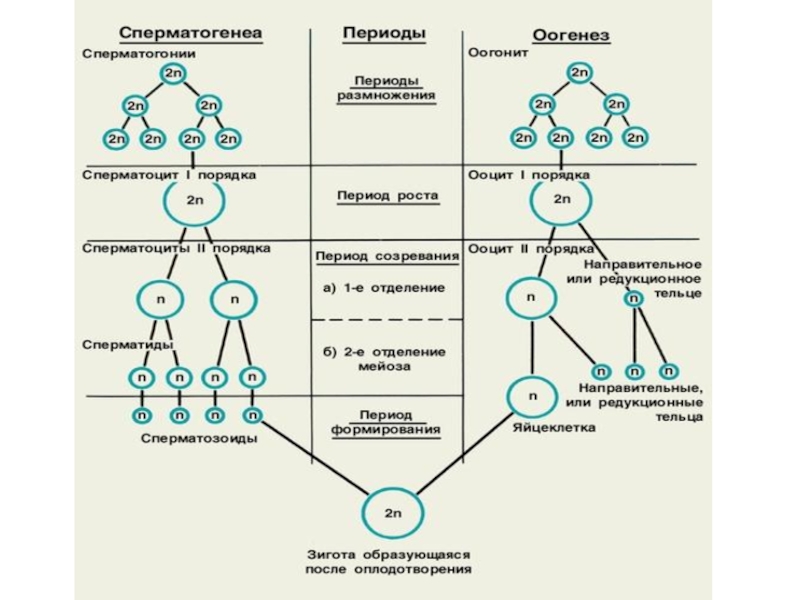
- 11. Второе деление мейоза
- 12. Мейоз - второе деление Интерфаза 2.
- 13. Мейоз - второе деление Профаза 2.
- 14. Мейоз В зависимости от места в жизненном
- 15. Механизм определение пола
- 16. Варианты определения пола Еще Мендель обратил
- 17. Варианты определения пола 1. При помощи
- 19. Хромосомное определение пола 1. ХY - тип.
- 20. Домашнее задание: § 26

Слайд 2Мейоз
Способ деления эукариот, при котором образуются клетки с гаплоидным набором хромосом,
В отличие от митоза, в мейозе два деления – первое (редукционное) и второе (обычный митоз без репликации). Первому делению мейоза предшествует интерфаза, перед вторым делением интерфаза не выражена. Оба деления мейоза состоят из тех же стадий, что и митоз: профаза, метафаза, анафаза и телофаза.


Слайд 4Мейоз - первое деление
Интерфаза 1.
Происходят процессы, аналогичные процессам в интерфазе
Удвоенные хромосомы при мейозе принято обозначать как 2n4с, где 2n показывает степень плоидности клетки, а 4с указывает на количество хромосомного материала (нитей ДНК).

Слайд 5Мейоз - первое деление
Профаза 1.
Занимает 90% всего времени мейоза.
Гомологичные
Процесс соединения гомологичных хромосом называется конъюгация.
Этот комплекс, состоящий из 4-х хроматид, называется бивалентом или тетрадой.
Бивалент

Слайд 6Профаза I
Несестринские хроматиды гомологичных хромосом бивалента тесно соединяются между собой и
Благодаря кроссинговеру происходит перекомбинация генов, и образовавшиеся в результате этого процесса хромосомы отличаются от исходных родительских.

Слайд 7Мейоз
Не для каждой хромосомы можно обнаружить пару. Например, в кариотипе мужских
Эту пару назвали парой половых хромосом (короткую — Y-хромосомой, а среднюю — Х-хромосомой) в противоположность 22 парам аутосом. Аутосомы одинаковы у представителей обоих полов, половые хромосомы у одного пола — одинаковые, у другого — разные.
Тем не менее половые хромосомы также образуют бивалент и в первом делении мейоза расходятся в разные клетки.

Слайд 8Профаза I
В первой профазе мейоза принято выделять отдельные стадии:
В лептотене (стадия
В зиготене (стадия сливающихся нитей) гомологичные хромосомы соединяются в биваленты;
В пахитене (стадия толстых нитей) происходит кроссинговер.
Диплотена (стадия двойных нитей) начинается взаимным отталкиванием гомологов и образованием хиазм (точка, в которой две гомологичные несестринские хроматиды обмениваются генетическим материалом). Целостность бивалента в это время сохраняется, в частности благодаря связи между сестринскими хроматидами в районе хиазм. У подавляющего большинства организмов в диплотене происходит дальнейшая конденсация хромосом. Лишь в ооцитах животных, накапливающих много желтка (некоторые рыбы, земноводные, птицы, млекопитающие), хромосомы, наоборот, деконденсируются и приобретают вид «ламповых щёток». Это наиболее длительный период профазы I, у человека может длиться 12-50 лет.
Для диакинеза (стадия обособленных двойных нитей) характерно уменьшение числа хиазм и значительная компактность бивалентов, конденсация хромосом завершается.
 происходит конденсация")
Слайд 9Мейоз - первое деление
Характерной особенностью профазы 1 мейоза, в отличие от
В ядрах созревающих женских половых клеток можно отчетливо видеть хромосомы с отходящими от них деконденсированными петлями хроматина, на которых идет транскрипция. Это хромосомы типа «ламповых щеток».

Слайд 10Мейоз - первое деление
Метафаза 1. Биваленты располагаются в экваториальной части клетки
Анафаза 1. Гомологичные хромосомы (а не хроматиды, как при митозе!), каждая из которых все еще состоит из 2-х хроматид, разъединяются и расходятся к полюсам клетки (2n4с).
Телофаза 1. Происходит образование 2-х дочерних клеток с гаплоидным набором хромосом, хотя каждая хромосома остается удвоенной, т.е. состоящей из 2-х хроматид. Такую клетку можно обозначить как 1n2с.
. Центромерные районы каждой")

Слайд 12Мейоз - второе деление
Интерфаза 2.
Эта стадия обычно непродолжительна по времени

Слайд 13Мейоз - второе деление
Профаза 2.
Ядрышки и ядерные мембраны вновь разрушаются,
Метафаза 2.
Удвоенные хромосомы выстраиваются на экваторе (1n2с).
Анафаза 2.
Хроматиды разъединяются и движутся к полюсам клетки (2n2с).
Телофаза 2.
Вокруг хромосом на полюсах клетки образуется ядерная оболочка, и начинается цитокинез.
В результате 2-х делений мейоза образуются 4 дочерних клетки-гаметы с гаплоидным набором хромосом (1n1с).

Слайд 14Мейоз
В зависимости от места в жизненном цикле организма различают три типа
- зиготный (у многих грибов и водорослей) – происходит в зиготе сразу после оплодотворения и приводит к образованию гаплоидного мицелия или таллома, а затем спор и гамет;
- гаметный (у всех многоклеточных животных и ряда низших растений) – происходит в половых органах и приводит к образованию гамет;
- споровый (у высших растений) – происходит при образовании спор, из которых прорастает гаплоидный гаметофит, в котором позднее образуются гаметы.


Слайд 16Варианты определения пола
Еще Мендель обратил внимание на то, что соотношение
Однако, существуют некоторые виды покрытосеменных растений, у которых пол действительно определяется одним геном, который наследуется по законам Менделя.
У растения, известного, как «бешеный огурец», серия множественных аллелей определяет пол цветков: ам – мужской пол, аж – женский пол, аг - обоеполые цветки, способные к самоопылению (ам > аг > аж).
Таким образом, пол растения зависит от одного гена.

Слайд 17Варианты определения пола
1. При помощи одного гена (бешеный огурец).
2.
3. После оплодотворения (у морского кольчатого червя Bonellia viridis, крокодила, мидии) в зависимости от условий среды.
4. В зависимости от факта оплодотворения (пчелы, муравьи - особи женского пола развиваются из диплоидных оплодотворенных яйцеклеток, а мужские особи из неоплодотворенных (партеногенез).
5. У большинства живых организмов пол формируется в момент оплодотворения и зависит от баланса хромосом. Это хромосомное определение пола.
. 2. До оплодотворения в зависимости")
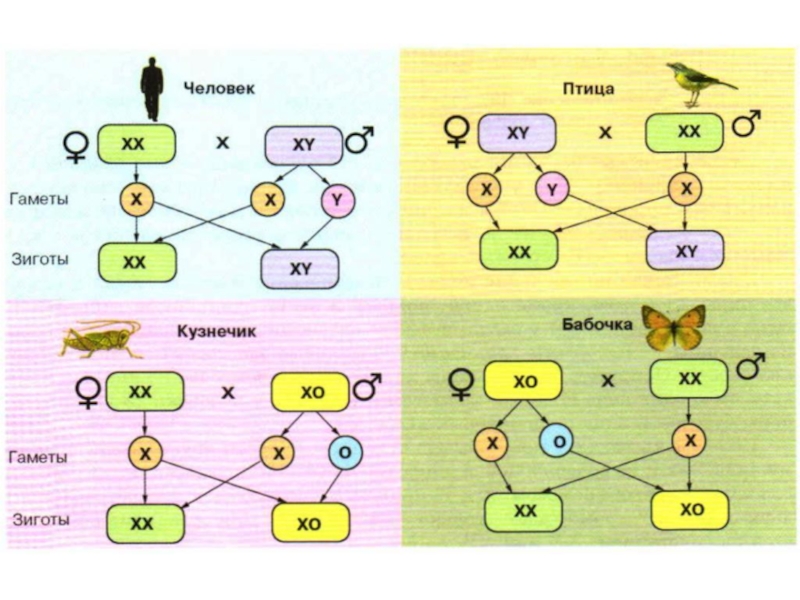
Слайд 19Хромосомное определение пола
1. ХY - тип. У мухи-дрозофилы и некоторых других
2. ZW – тип. У птиц, пресмыкающихся и некоторых насекомых (чешуекрылые) женский пол – гетерогаметен (ZW), а мужской – гомогаметен (ZZ).
3. ХО – тип. У некоторых мух, клопов, жуков, пауков и кузнечиков у самцов только одна половая хромосома (ХО), самки гомогаметны (ХХ).
4. ZО – тип. У моли, наоборот, самцы гомогаметны (ZZ), а самки имеют только одну половую хромосому (ZО).
5. 2n - 1n – тип. У пчел и муравьев самки диплоидны, самцы гаплоидны.