- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Метаболизм ДНК презентация
Содержание
- 1. Метаболизм ДНК
- 2. Метаболизм ДНК Репликация – процесс точного копирования
- 3. Репликация Процесс самовоспроизведения макромолекул нуклеиновых кислот, обеспечивающий
- 4. Репликация ДНК происходит в определенной фазе
- 5. Основные принципы репликации ДНК Полуконсервативный механизм -
- 7. Субстратами для синтеза ДНК являются дезоксирибонуклеозидтрифосфаты
- 8. Точка начала репликации – origin (ориджин).
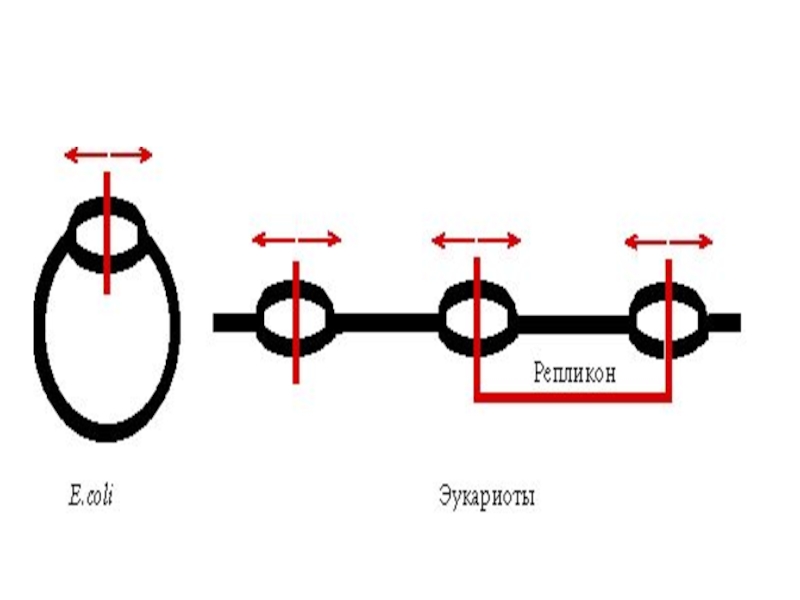
- 10. Репликон – это участок ДНК между двумя «ориджинами» репликации.
- 11. Родительская ДНК Репликационный глазок Дочерние молекулы ДНК Репликативные вилки
- 12. Репликация идет в двух направлениях
- 13. Синтез новых цепей ДНК может протекать
- 14. 5’ 3’ 3’ 5’
- 15. Синтез ДНК
- 16. Вся сложность ферментативного аппарата репликации связана
- 17. Ферменты репликации Синтез новой цепи ДНК осуществляется
- 18. Синтез ДНК У прокариот найдено 3 типа
- 19. ДНК-полимераза I выполняет функции проверки поддержания
- 20. ДНК-полимераза II – участвует в процессах репарации ДНК при повреждении ДНК ультрафиолетовым облучением.
- 21. ДНК-полимераза III – главный фермент репликации
- 22. Для создания Матрицы – одноцепочечной ДНК,
- 23. Хеликазы – это ферменты, способные расплетать
- 24. Топоизомеразы – ферменты, изменяющие степень сверхспирализации
- 25. Топоизомеразы находятся перед репликативной вилкой, разрезают
- 26. Антибиотики – ингибиторы топоизомеразы: Хинолоны, фторхинолоны Норфлоксацин
- 27. белки, стабилизирующие разделенные нити ДНК – SSB (single strand binding)
- 28. Роль SSB -белков заключается в том,
- 29. ДНК-лигаза – фермент катализирующий сшивание одноцепочечных
- 30. Синтез ДНК Синтез ДНК включает в себя три этапа: инициация элонгация терминация
- 31. Инициация инициация синтеза ДНК у E.coli происходит
- 32. Процесс инициации начинается с присоединения к
- 33. DnaА белки Dna A белки
- 34. Это приводит к разделению цепей и
- 35. Хеликазы Хеликазы разделяют ДНК в двух направлениях в районе двух вилок вилка вилка
- 37. Синтез ДНК В результате действия хеликазы,
- 38. Синтез ДНК ДНК-полимераза III не способна начинать
- 39. Синтез ДНК Праймер - короткий сегмент РНК,
- 40. Синтез ДНК от 3’ –конца праймера начинается
- 41. Синтез ДНК ДНК-полимераза III удлиняет РНК-затравку, присоединяя
- 42. Элонгация Синтез лидирующей цепи. Синтез отстающей цепи.
- 43. Синтез лидирующей цепи начинается с синтеза
- 44. Синтез отстающей цепи Синтез осуществляется
- 45. ДНК-полимераза III связывается с праймером и
- 46. Реплисома присоединяет по 1000 нуклеотидов в
- 47. Оставшийся разрыв «сшивает» ДНК-лигаза. ДНК-лигаза
- 49. Терминация У прокариот есть специальные терминаторы (ter)
- 50. Комплекс Ter-Tus может задержать репликативную вилку,
- 51. Для терминации репликации линейных эукариотических хромосом
- 52. Это многократно повторяющиеся последовательности нуклеотидов (
- 53. Теломеры присоединяются к концам эукариотических хромосом с помощью фермента теломеразы.
- 54. Одна из причин старения — закон
- 55. есть клетки, которые могут делиться бесконечно,
- 56. Источники повреждения ДНК УФ излучение
- 57. Репарация ДНК Репарация- процесс восстановления повреждений ДНК.
- 58. Репарация осуществляется с помощью: специфического набора
- 59. У бактерий имеются 2 ферментные системы,
- 60. Прямая репарация ДНК Фотореактивация. Расщепление пиримидиновых димеров
- 61. фермент ДНК – фотолиаза -
- 62. Репарация ДНК
- 63. Репарация ДНК Темновая эксцизионная репарация Не нуждается в энергии видимого света
- 64. Каждая из систем репарации включает следующие компоненты:
- 65. Эксцизионная репарация Base excision repair – BER
- 66. Фосфодиэстераза отщепляет от ДНК сахарофосфатную
- 67. Nucleotide excision repair - NER Узнавание
- 68. Освобождение олигонуклеотида, содержащего
- 69. При некоторых типах повреждений ДНК (двухнитевые
- 70. SOS-репарация SOS – белки всегда присутствуют в
- 71. В геноме типичной клетки млекопитающих за
- 72. Все дефекты генов белков, участвующих в
- 73. У человека нет ДНК-фотолиазы и эксцизионная
- 74. Молекулярные мутации Анемия Фанкони – генетические отклонения,
- 75. У 60—75 % больных также встречаются врожденные
- 76. Спасибо за внимание

Слайд 2Метаболизм ДНК
Репликация – процесс точного копирования молекулы ДНК.
Репарация – поддержание целостности
биологического материала в ДНК.
Рекомбинация –перестройка заключенной в молекуле ДНК генетической информации.
Рекомбинация –перестройка заключенной в молекуле ДНК генетической информации.

Слайд 3Репликация
Процесс самовоспроизведения макромолекул нуклеиновых кислот, обеспечивающий точное копирование генетического материала и
передачу его от поколения к поколению.
В основе репликации лежит понятие матрицы – это макромолекулярная структура для синтеза комплементарной копии макромолекулы.
В основе репликации лежит понятие матрицы – это макромолекулярная структура для синтеза комплементарной копии макромолекулы.

Слайд 4
Репликация ДНК происходит в определенной фазе клеточного цикла.
Основные свойства процесса репликации
ДНК и каталитические механизмы этого процесса практически идентичны у всех видов организмов.

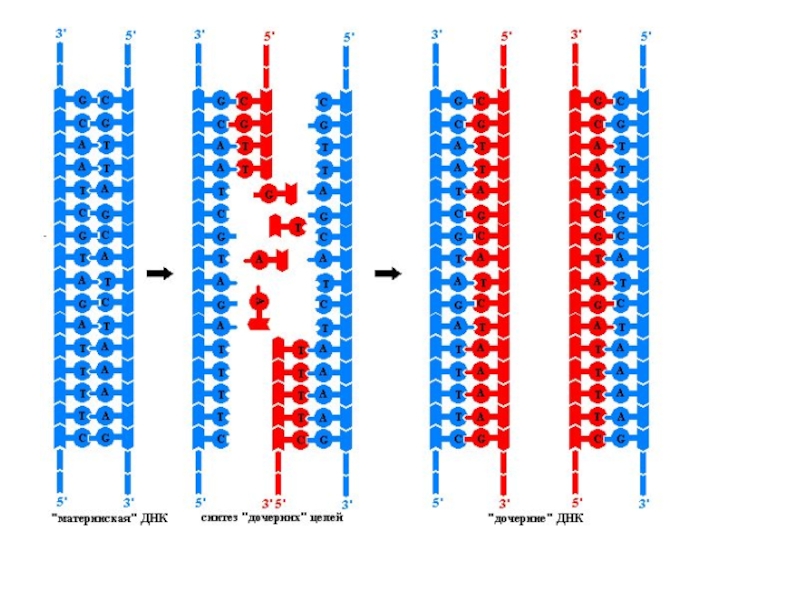
Слайд 5Основные принципы репликации ДНК
Полуконсервативный механизм - каждая из 2 цепей ДНК
служит матрицей для образования новой цепи. Образуются две новые двухцепочечные молекулы ДНК, каждая из которых состоит из одной новой и одной старой цепей.

Слайд 7
Субстратами для синтеза ДНК являются дезоксирибонуклеозидтрифосфаты – dАТФ, dГТФ, dТТФ, dЦТФ,
выполняющие роль строительного материала и источников энергии.

Слайд 8
Точка начала репликации – origin (ориджин).
У бактерий в кольцевом геноме имеется
только одна точка «origin», тогда как у эукариотических хромосом их множество.
У человека репликация начинается в точках, удаленных друг от друга от 30 до 300тыс.п.н.
У человека репликация начинается в точках, удаленных друг от друга от 30 до 300тыс.п.н.
.У бактерий в кольцевом геноме имеется только одна точка «origin»,")



Слайд 13
Синтез новых цепей ДНК может протекать только в направлении
5’
→ 3’, так как достраивается 3’-конец цепи ДНК.



Слайд 16
Вся сложность ферментативного аппарата репликации связана с требованиями высокой точности синтеза.
Весь этот комплекс называется ДНК-репликативной системой, или
Реплисомой.

Слайд 17Ферменты репликации
Синтез новой цепи ДНК осуществляется при помощи ДНК-полимеразы.
Фермент катализирует
синтез дочерних цепей на матрице ДНК по принципу комплементарности.

Слайд 18Синтез ДНК
У прокариот найдено 3 типа этих ферментов:
ДНК-полимераза I
ДНК-полимераза II
ДНК-полимераза III
ДНК-полимераза III

Слайд 19
ДНК-полимераза I выполняет функции проверки поддержания порядка во время репликации, репарации
и рекомбинации.
Обладает 5’ → 3’ экзонуклеазной активностью, может заменить участок ДНК (или РНК), что используется в методах по гибридизации нуклеиновых кислот.
Обладает 5’ → 3’ экзонуклеазной активностью, может заменить участок ДНК (или РНК), что используется в методах по гибридизации нуклеиновых кислот.

Слайд 20
ДНК-полимераза II – участвует в процессах репарации ДНК при повреждении ДНК
ультрафиолетовым облучением.

Слайд 21
ДНК-полимераза III – главный фермент репликации у E.coli.
Состоит из субъединиц 10
разных типов.
Обладает полимеразной активностью (присоединяет 250-1000 нуклеотидов/с).
Корректирующей активностью, т.е. 3’ → 5’ экзонуклеазной активностью, благодаря которой каждый нуклеотид после присоединения проверяется дважды.
Обладает полимеразной активностью (присоединяет 250-1000 нуклеотидов/с).
Корректирующей активностью, т.е. 3’ → 5’ экзонуклеазной активностью, благодаря которой каждый нуклеотид после присоединения проверяется дважды.

Слайд 22
Для создания Матрицы – одноцепочечной ДНК, необходимы хеликазы – ферменты, разделяющие
цепи двухцепочечной ДНК на одинарные цепи.

Слайд 23
Хеликазы – это ферменты, способные расплетать две комплементарные нити в ДНК
с использованием энергии, полученной при гидролизе АТФ.

Слайд 24
Топоизомеразы – ферменты, изменяющие степень сверхспирализации ДНК, путем внесения одноцепочечных
или двухцепочечных разрывов в ДНК.

Слайд 25
Топоизомеразы находятся перед репликативной вилкой, разрезают молекулу ДНК для облегчения ее
расплетания и раскручивания молекулы ДНК, после чего непрерывность ее восстанавливается.


")
Слайд 28
Роль SSB -белков заключается в том, что они связываются с однонитчатой
ДНК, выпрямляют ее и блокируют образование шпилечных двухнитчатых структур.
Участие SSB в репликации абсолютно необходимо. Они удерживают матричные цепи ДНК в репликативной вилке в одноцепочечном состоянии, а также защищают одноцепочечную ДНК от действия нуклеаз.
Участие SSB в репликации абсолютно необходимо. Они удерживают матричные цепи ДНК в репликативной вилке в одноцепочечном состоянии, а также защищают одноцепочечную ДНК от действия нуклеаз.

Слайд 29
ДНК-лигаза – фермент катализирующий сшивание одноцепочечных фрагментов ДНК.
ДНК-праймаза — это
фермент РНК-полимераза, синтезирующий короткий фрагмент РНК, называемый праймером, комплементарный одноцепочечной матрице ДНК.


Слайд 31Инициация
инициация синтеза ДНК у E.coli происходит в oriC (245п.н.)
В точках начала
репликации отмечено большое количество А=Т пар и участки связывания ключевого инициаторного белка DnaA.
В точках начала репликации отмечено большое количество")
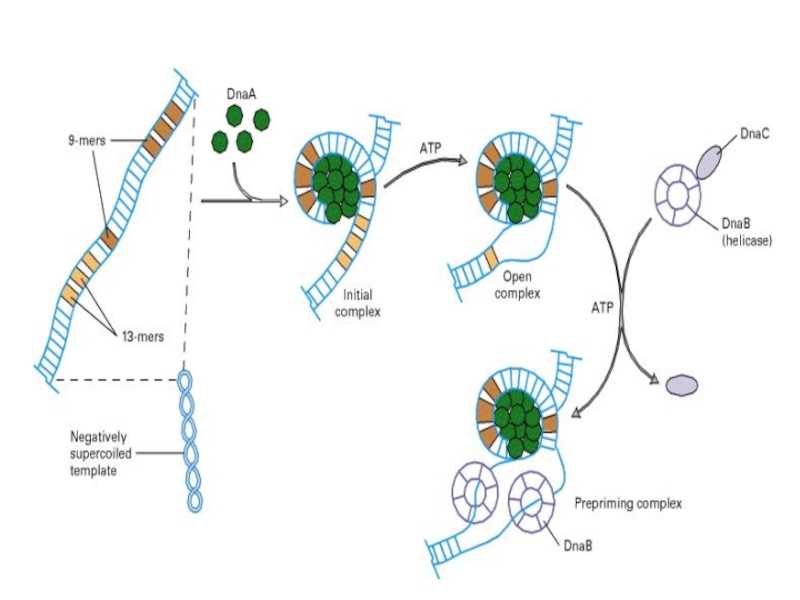
Слайд 32
Процесс инициации начинается с присоединения к хромосоме белка DnaA (в origin
области).
8 молекул DnaA, каждая из которых связана с АТФ, образуют спиральный комплекс (положительная суперспираль). В соседних участках ДНК это вызывает денатурацию А=Т богатого участка.
8 молекул DnaA, каждая из которых связана с АТФ, образуют спиральный комплекс (положительная суперспираль). В соседних участках ДНК это вызывает денатурацию А=Т богатого участка.
. 8 молекул DnaA,")
Слайд 33
DnaА белки
Dna A белки садятся на oriC – участок начала репликации
Индуцируется
расплетение ДНК
Расплетенный участок индуцирует прикрепление SSB-белков и хеликаз (Dna B)
Хеликазы
SSB - белки

Слайд 34
Это приводит к разделению цепей и способствует работе основного расплетающего белка
- хеликазы (DnaB).
С образовавшейся одноцепочечной ДНК связываются белки SSB, которые стабилизируют вилку репликации.
С образовавшейся одноцепочечной ДНК связываются белки SSB, которые стабилизируют вилку репликации.
. С")

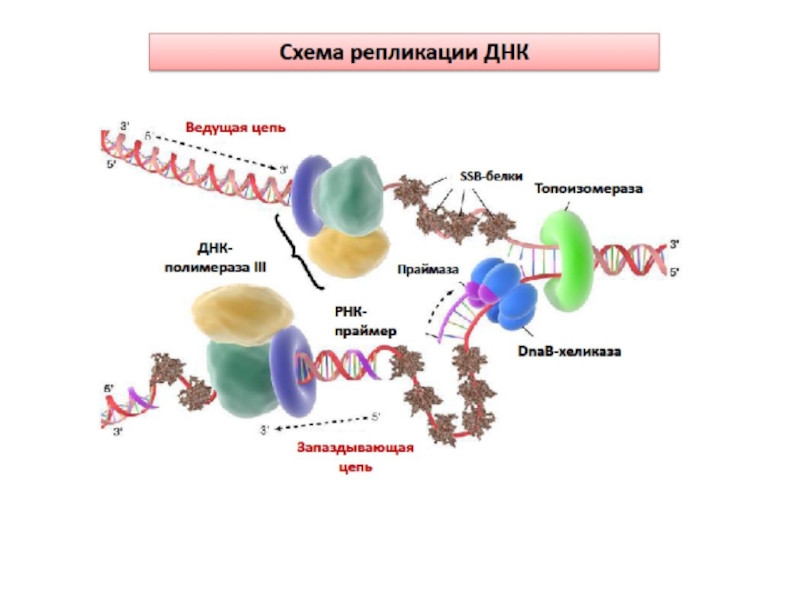
Слайд 37 Синтез ДНК
В результате действия хеликазы, топоизомеразы, SSB- белков, ионов Mg+2
образуется репликативная вилка - участок ДНК, в пределах которого спираль раскручена и разделена на отдельные цепи.

Слайд 38Синтез ДНК
ДНК-полимераза III не способна начинать синтез новой цепи с ее
первого нуклеотида. Поэтому репликация начинается с синтеза праймера (РНК-затравки) на обеих цепях расплетенной ДНК.

Слайд 39Синтез ДНК
Праймер - короткий сегмент РНК, комплементарный матричной цепи ДНК.
Праймер
синтезируется при участии ДНК- зависимой РНК-полимеразы (праймазы).

Слайд 40Синтез ДНК
от 3’ –конца праймера начинается синтез новой цепи ДНК при
помощи ДНК- полимераза III. Синтез идет в направлении 5’ → 3’

Слайд 41Синтез ДНК
ДНК-полимераза III удлиняет РНК-затравку, присоединяя к ней один за другим
нуклеотиды, комплементарные матричной цепи


Слайд 43
Синтез лидирующей цепи начинается с синтеза праймера, дезоксирибонуклеотиды добавляются к этому
праймеру ДНК-полимеразой III, связанной с DnaВ- хеликазой.
Синтез идет непрерывно со скоростью, соответствующей скорости раскручивания ДНК в репликативной вилке.
Синтез идет непрерывно со скоростью, соответствующей скорости раскручивания ДНК в репликативной вилке.

Слайд 44
Синтез отстающей цепи
Синтез осуществляется в виде коротких фрагментов ОКАЗАКИ.
Каждый фрагмент Оказаки
состоит примерно из 1000 нуклеотидов
(у прокариот).
Праймаза синтезирует РНК-праймер.
(у прокариот).
Праймаза синтезирует РНК-праймер.

Слайд 45
ДНК-полимераза III связывается с праймером и присоединяет дезоксирибонуклеотиды.
Синтез фрагментов Оказаки происходит
с участием ферментативного аппарата – праймасомы ( DnaB-хеликаза и DnaG-праймаза) в составе репликативного комплекса.

Слайд 46
Реплисома присоединяет по 1000 нуклеотидов в секунду к каждой цепи (лидирующей
и отстающей).
После завершения сборки одного фрагмента Оказаки его РНК-праймер удаляется и замещается последовательностью ДНК с помощью ДНК-полимеразы I.
После завершения сборки одного фрагмента Оказаки его РНК-праймер удаляется и замещается последовательностью ДНК с помощью ДНК-полимеразы I.
.После завершения сборки")
Слайд 47
Оставшийся разрыв «сшивает» ДНК-лигаза.
ДНК-лигаза катализирует образование фосфодиэфирной связи между 3’-гидроксильной
группой на конце одной цепи ДНК и 5’-фосфатом на конце другой цепи.
Для этой реакции используется НАД-кофактор в качестве источника АМФ (у прокариот).
Для этой реакции используется НАД-кофактор в качестве источника АМФ (у прокариот).

Слайд 49Терминация
У прокариот есть специальные терминаторы (ter) – специальные последовательности нуклеотидов, прекращающие
синтез цепи ДНК
Ter-последовательности служат участками связывания белка Tus (terminus utilization substance)
Ter-последовательности служат участками связывания белка Tus (terminus utilization substance)
 – специальные последовательности нуклеотидов, прекращающие синтез цепи ДНКTer-последовательности служат")
Слайд 50
Комплекс Ter-Tus может задержать репликативную вилку, движущуюся только в одном направлении,
т.е. останавливает одну из вилок с которой сталкивается. Другая вилка останавливается, когда встречается с первой задержанной вилкой.

Слайд 51
Для терминации репликации линейных эукариотических хромосом на концах каждой хромосомы синтезируются
специальные структуры, называемые теломерами.
Иначе с каждым клеточным делением хромосомы становились бы короче и короче.
Иначе с каждым клеточным делением хромосомы становились бы короче и короче.

Слайд 52
Это многократно повторяющиеся последовательности нуклеотидов ( у одноклеточных эукариот от 20
до100, у млекопитающих превышает 1500).
У человека такая последовательность (TTAGGG)n .
У человека такая последовательность (TTAGGG)n .


Слайд 54
Одна из причин старения — закон делимости клеток, открытый американским биологом
Леонардом Хейфликом (Leonard Hayflick) в 1961 году. Суть его в том, что клетки человеческого организма не могут делиться бесконечно. Максимально возможное количество делений в среднем составляет 50±10 (так называемый предел или лимит Хейфлика).
Вызвано это тем, что при делении клетки репликация ДНК происходит "с потерями" — молекула копируется не до конца. До определенного момента в этом ничего страшного нет — кончики хромосом защищены теломерами, которые и теряются при копировании..
Вызвано это тем, что при делении клетки репликация ДНК происходит "с потерями" — молекула копируется не до конца. До определенного момента в этом ничего страшного нет — кончики хромосом защищены теломерами, которые и теряются при копировании..
")
Слайд 55
есть клетки, которые могут делиться бесконечно, например кроветворные или раковые. В
этих случаях механизм ограничения количества делений не срабатывает благодаря теломеразе, которая "чинит" ДНК, достраивая теломеры.

Слайд 56
Источники повреждения ДНК
УФ излучение
Радиация
Химические вещества
Ошибки репликации ДНК
Апуринизация -
отщепление азотистых оснований от сахарофосфатного остова
Дезаминирование - отщепление аминогруппы от азотистого основания
Дезаминирование - отщепление аминогруппы от азотистого основания

Слайд 57Репарация ДНК
Репарация- процесс восстановления повреждений ДНК.
Распознавание дефекта сопровождается непосредственно при репликации,
все ДНК-полимеразы обладают 3’ → 5’ экзонуклеазной активностью.

Слайд 58
Репарация осуществляется с помощью: специфического набора ферментов, постоянно присутствующих в нормально
функционирующих клетках (фотореактивационная, эксцизионная)
активации группы генов, контролирующих различные клеточные функции –
SOS-репарация.
активации группы генов, контролирующих различные клеточные функции –
SOS-репарация.
")

Слайд 60Прямая репарация ДНК
Фотореактивация. Расщепление пиримидиновых димеров (приУФ облучении) осуществляется ферментом
ДНК - фотолиазой.
Реакция расщепления связей зависит от видимого света. У человека отсутствует.
Реакция расщепления связей зависит от видимого света. У человека отсутствует.
 осуществляется ферментом ДНК - фотолиазой. Реакция")
Слайд 61
фермент ДНК – фотолиаза - мономерный флавин-зависимый фермент и 2
кофактора (хромофоры).
5,10-метенилтетрагидрофолат (5,10-MTГФ) –поглощает фотоны синего цвета (300-500 нм) и передает энергию возбуждения на FADH- .
Возбужденный FADH- отдает электрон пиримидиновому димеру, устраняя повреждение.
5,10-метенилтетрагидрофолат (5,10-MTГФ) –поглощает фотоны синего цвета (300-500 нм) и передает энергию возбуждения на FADH- .
Возбужденный FADH- отдает электрон пиримидиновому димеру, устраняя повреждение.
. 5,10-метенилтетрагидрофолат (5,10-MTГФ)")


Слайд 64Каждая из систем репарации включает следующие компоненты:
фермент, «узнающий» химически изменённые участки
в цепи ДНК и осуществляющий разрыв цепи вблизи от повреждения.
фермент, удаляющий повреждённый участок.
фермент, синтезирующий соответствующий участок цепи ДНК взамен удалённого.
фермент, замыкающий последнюю связь в полимерной цепи и тем самым восстанавливающий её непрерывность.
фермент, удаляющий повреждённый участок.
фермент, синтезирующий соответствующий участок цепи ДНК взамен удалённого.
фермент, замыкающий последнюю связь в полимерной цепи и тем самым восстанавливающий её непрерывность.

Слайд 65Эксцизионная репарация
Base excision repair – BER
ДНК гликозилазы, распознают аномальные основания ДНК
и катализируют гидролитическое расщепление N-glycosyl связи, между основанием и сахаром.
Образуется AP-сайт (apurinic/apyrimidinic).
АП-сайт распознается АП-эндонуклеазой,
которая вводит в нить ДНК разрыв.
Образуется AP-сайт (apurinic/apyrimidinic).
АП-сайт распознается АП-эндонуклеазой,
которая вводит в нить ДНК разрыв.

Слайд 66
Фосфодиэстераза отщепляет от ДНК
сахарофосфатную группу, к которой не
присоединено основание.
Брешь размеров в 1 н. застраивается
ДНК-полимеразой I и концы ДНК
соединяются ДНК-лигазой.

Слайд 67
Nucleotide excision repair - NER
Узнавание повреждений.
Связывание мультисубъединичного комплекса с
поврежденным сайтом.
Двойное надрезание поврежденной цепи на несколько нуклеотидов.
от поврежденного сайта в обоих направлениях
5' и 3'.
Двойное надрезание поврежденной цепи на несколько нуклеотидов.
от поврежденного сайта в обоих направлениях
5' и 3'.

Слайд 68
Освобождение олигонуклеотида, содержащего повреждение между двумя
надрезами.
Заполнение образовавшейся бреши ДНК полимеразой.
Лигирование.
Заполнение образовавшейся бреши ДНК полимеразой.
Лигирование.

Слайд 69
При некоторых типах повреждений ДНК (двухнитевые разрывы,поперечные сшивки).
Репликативная вилка наталкивается
на нерепарированное повреждение ДНК.
Результат воздействия ионизирующей радиации.
Окислительные реакции.
Результат воздействия ионизирующей радиации.
Окислительные реакции.
. Репликативная вилка наталкивается на нерепарированное повреждение ДНК.Результат")
Слайд 70SOS-репарация
SOS – белки всегда присутствуют в клетке, но при запуске SOS-ответа
их уровень значительно повышается.
Мутации, возникающие в результате такой репликации, приводят к гибели одних клеток и опасны для других, но репликация не прекращается, позволяет выжить некоторым мутантным дочерним клеткам.
Мутации, возникающие в результате такой репликации, приводят к гибели одних клеток и опасны для других, но репликация не прекращается, позволяет выжить некоторым мутантным дочерним клеткам.

Слайд 71
В геноме типичной клетки млекопитающих за 24 ч аккумулируется много тысяч
повреждений. Благодаря репарации менее одного повреждения из 1000 становится мутацией. Изменения в генах репарации значительно повышают чувствительность к раку.

Слайд 72
Все дефекты генов белков, участвующих в эксцизионной репарации связаны с онкологическими
заболеваниями,
генетическими заболеваниями, например, пигментная ксеродерма.
генетическими заболеваниями, например, пигментная ксеродерма.

Слайд 73
У человека нет ДНК-фотолиазы и эксцизионная репарация оснований – единственный способ
репарации пиримидиновых димеров.
Инактивация этой системы связана с развитием рака кожи, который индуцируется солнечным светом.
Инактивация этой системы связана с развитием рака кожи, который индуцируется солнечным светом.

Слайд 74Молекулярные мутации
Анемия Фанкони – генетические отклонения, возникающие при репарации ДНК.
Болезнь
названа в честь швейцарского педиатра, Гвидо Фанкони, впервые описавшего это заболевание.

Слайд 75
У 60—75 % больных также встречаются врожденные дефекты, такие как низкорослость, ненормальная
пигментация, маленькая голова, аномалии скелета
Ряд неврологических расстройств (косоглазие, недоразвитие одного или обоих глаз, опущение века, глазное дрожание, глухота, умственная отсталость),
Поражения половых органов (недоразвитие половых органов)
Почечные аномалии
Врождённые пороки сердца.
Средняя продолжительность жизни у больных анемией Фанкони составляет около 30 лет.
Ряд неврологических расстройств (косоглазие, недоразвитие одного или обоих глаз, опущение века, глазное дрожание, глухота, умственная отсталость),
Поражения половых органов (недоразвитие половых органов)
Почечные аномалии
Врождённые пороки сердца.
Средняя продолжительность жизни у больных анемией Фанкони составляет около 30 лет.







