- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Матричные биосинтезы презентация
Содержание
- 1. Матричные биосинтезы
- 2. Направление роста цепи 5'→3' Поступающий нуклеотид он
- 3. Биосинтез ДНК
- 4. Полуконсервативность биосинтеза ДНК
- 6. Особенности репликации
- 7. Этапы репликации 1. Инициация: Топоизомераза находит точку
- 8. Этапы репликации ДНК-связывающие белки (SSB-белки) стабилизируют репликативную
- 9. Инициация репликации ДНК-полимераза α ДНК-полимераза δ ДНК-полимераза ε
- 10. Ориджин репликации репликон Инициация репликации
- 11. 2. Элонгация ДНК-полимераза δ продолжает удлинять нить
- 12. Элонгация репликации
- 13. 3. Терминация ДНК-полимераза β (фермент репарации)
- 14. Функции ДНК-полимеразы β
- 15. Репарация ДНК
- 16. Для биосинтеза РНК (транскрипции) необходимы: МАТРИЦА –
- 17. Биосинтез РНК
- 18. Транскрибируемая нить Нетранскрибируемая нить РНК-полимераза ДНК пре-РНК (первичный транскрипт) Биосинтез РНК мононуклеотиды Сайт терминации
- 19. Процессинг РНК (1. сплайсинг)
- 20. м-РНК 7-метилгуанозин полиаденилат 5' 3' Процессинг (2. модификация концов м-РНК)
- 21. Состав зрелой м-РНК
- 22. Общая схема биосинтеза белка
- 23. Компоненты белоксинтезирующей системы мРНК 20
- 24. Ядерные стадии биосинтеза белка Биосинтез мРНК (транскрипция) Созревание мРНК (посттранскрипционный процессинг)
- 25. Функции т-РНК Акцепторная: связывает аминокислоту и транспортирует
- 26. Строение т-РНК антикодон
- 27. Таблица генетического кода
- 28. Характеристика генетического кода Триплетность (1 аминокислота кодируется
- 29. Колинеарность генетического кода Глу-тРНК
- 30. Цитоплазматические стадии биосинтеза белка Активация аминокислот, или образование аминоацил-тРНК Инициация Элонгация Терминация Посттрансляционная модификация
- 31. Синтез аминоацил-тРНК
- 32. Рибосома эукариотов
- 33. Функции активной рибосомы Связывание и удерживание белоксинтезирующей
- 34. Функционирующая рибосома
- 35. Образование инициирующего комплекса
- 36. Инициация: К малой субъединице рибосомы прикрепляется м-РНК
- 37. Инициация трансляции
- 38. Элонгация: В пептидильном центре находится Мет-т-РНК, а
- 39. ГТФ ГДФ+НР
- 40. Строение полирибосомы
- 41. Посттрансляционный процессинг Модификация N-конца полипептидной цепи Фолдинг
- 42. Регуляция биосинтеза 1: РНК полимераза, 2: Репрессор,
- 43. Действие регуляторных белков
- 44. Регуляция биосинтеза Энхансеры – участки ДНК, присоединение
- 45. Амплификация
- 46. Типы генов в геноме Структурные гены (кодируют
- 47. Клеточная дифференцировка
- 48. Классификация мутаций
- 49. Типы мутаций
- 50. Типы генных мутаций
- 51. Проявления мутаций
- 53. Механизмы увеличения числа и разнообразия генов в геноме
- 54. Механизмы увеличения числа и разнообразия генов в геноме (кроссинговер)
- 55. Механизмы увеличения числа и разнообразия генов в геноме (амплификация)
- 56. Полиморфизм белков –
- 57. Схема возникновения наследственных болезней
- 58. Биохимические проявления наследственных болезней
- 59. Клинические проявления наследственных болезней
- 60. КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ КАФЕДРА
- 61. Биологическая роль липидов Резерв и источник энергии
- 62. Классификация липидов
- 63. Классификация простых липидов Простые липиды:
- 64. Строение простых липидов
- 65. Классификация фосфолипидов
- 66. Строение глицеролфосфолипидов
- 67. Строение сфингофосфолипидов
- 68. Классификация гликолипидов
- 69. Строение сфингогликолипидов
- 70. Жирные кислоты тканей человека
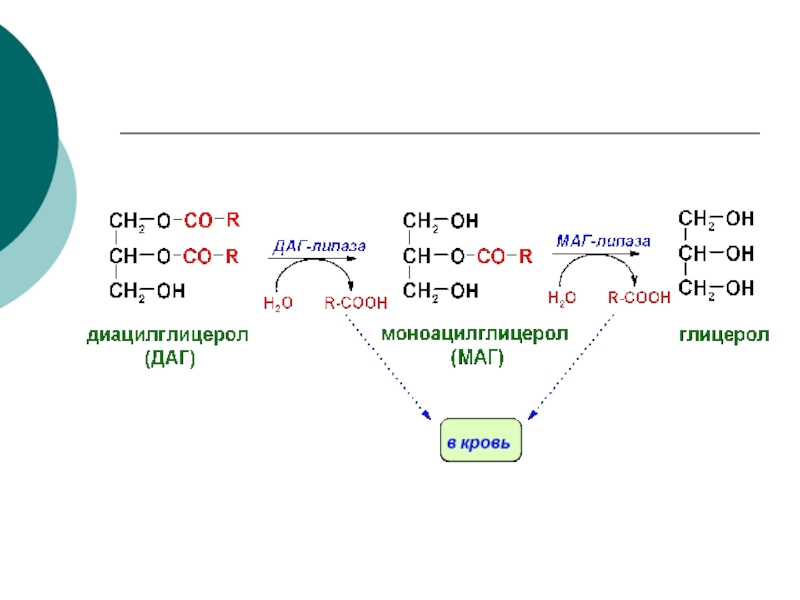
- 71. Переваривание нейтрального жира глицерол триглицерид моноглицерид диглицерид липаза липаза липаза
- 72. Строение желчных кислот
- 73. Переваривание фосфолипидов
- 74. Переваривание стеридов
- 75. Ресинтез нейтрального жира (II)
- 76. Строение хиломикрона
- 77. Роль липопротеинлипазы липопротеинлипаза триглицерид (ТАГ) глицерол
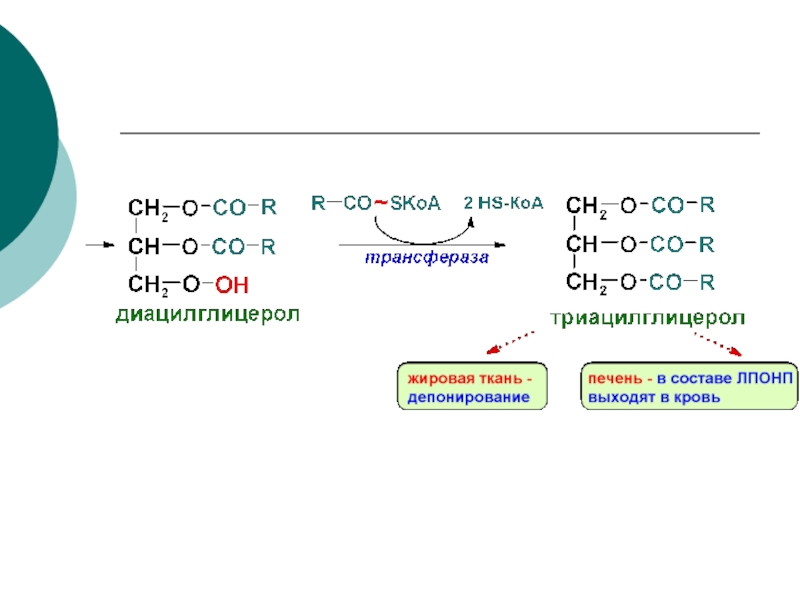
- 78. СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ В ПЕЧЕНИ И ЖИРОВОЙ ТКАНИ
- 80. МОБИЛИЗАЦИЯ ТРИАЦИЛ-ГЛИЦЕРОЛОВ
- 82. Активация жирных кислот
- 83. ТРАНСПОРТ ЖИРНЫХ КИСЛОТ В МИТОХОНДРИИ
- 84. β-ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ 2 АТФ
- 85. 3 АТФ ЦТК 12 АТФ следующий цикл β-окисления
- 86. ОБЩАЯ СХЕМА ЦИКЛА β–ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ дегидрогеназа
- 87. ЭНЕРГЕТИЧЕСКИЙ ИТОГ
- 88. Окисление ненасыщенных жирных кислот Цис-транс- β-окисление …
- 89. ИСТОЧНИКИ АЦЕТИЛ-КоА
- 90. Строение пальмитоилсинтетазы
- 91. Биосинтез пальмитиновой кислоты
- 92. Схема биосинтеза пальмитиновой кислоты
- 93. Схема биосинтеза пальмитиновой кислоты
- 94. Суммарное уравнение биосинтеза пальмитиновой кислоты
- 95. Регуляция биосинтеза и окисления жирных кислот активная неактивная
- 96. Удлинение жирных кислот
- 97. Биосинтез непредельных жирных кислот
- 98. Синтез кетоновых тел
- 100. Окисление кетоновых тел
- 101. Типы липопротеинов
- 102. Состав липопротеинов
Слайд 1
КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ
КАФЕДРА ФУНДАМЕНТАЛЬНОЙ И КЛИНИЧЕСКОЙ БИОХИМИИ
Лекция по биохимии
Тема:
«Матричные биосинтезы»
Краснодар
2016

Слайд 2Направление роста цепи
5'→3'
Поступающий нуклеотид
он х
х
х
Растущая цепь НК
Матрица – цепь ДНК
5'
5'
3'
3'
Принцип биосинтеза
НК

Слайд 3Биосинтез ДНК
матричным (матрица – обе нити ДНК)
комплиментарным
фрагментарным (нити ДНК синтезируются в виде фрагментов, которые затем соединяются между собой)
полуконсервативным (в каждой из образовавшихся молекул ДНК одна нить исходная – материнская, а одна – вновь синтезированная – дочерняя)
 является:матричным (матрица – обе нити ДНК)комплиментарнымфрагментарным (нити")



Слайд 7Этапы репликации
1. Инициация:
Топоизомераза находит точку начала репликации, гидролизует одну фосфодиэфирную связь
Хеликаза разрывает водородные связи между нитями ДНК

Слайд 8Этапы репликации
ДНК-связывающие белки (SSB-белки) стабилизируют репликативную вилку, не давая восстанавливаться водородным
ДНК-полимераза α (праймаза) строит праймер («затравку») из 8-10 рибонуклео-тидов и 40-50 дезоксирибонуклеотидов, а ДНК-полимераза δ достраивает нить из дезоксирибонуклеотидов на лидирующей нити, а ДНК-полимераза ε – на отстающей нити ДНК
 стабилизируют репликативную вилку, не давая восстанавливаться водородным связям между комплиментарными нуклеотидами")


Слайд 11 2. Элонгация
ДНК-полимераза δ продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей нити,
Этапы репликации


Слайд 13 3. Терминация
ДНК-полимераза β (фермент репарации) удаляет праймеры и достраивает фрагменты ДНК
ДНК-лигаза соединяет фрагменты между собой
Этапы репликации
 удаляет праймеры и достраивает фрагменты ДНК ДНК-лигаза соединяет фрагменты между")


Слайд 16Для биосинтеза РНК (транскрипции) необходимы:
МАТРИЦА – участок одной из нитей ДНК
– (транскриптон)
СТРОИТЕЛЬНЫЙ МАТЕРИАЛ:
– АТФ, ГТФ, ЦТФ, УТФ
ФЕРМЕНТАТИВНЫЕ БЕЛКИ
ДНК-зависимые РНК-полимеразы
I — для синтеза р-РНК
II — для синтеза м-РНК
III — для синтеза т-РНК
РЕГУЛЯТОРНЫЕ БЕЛКИ: факторы инициации, элонгации, терминации
 необходимы:МАТРИЦА – участок одной из нитей ДНК – (транскриптон)СТРОИТЕЛЬНЫЙ МАТЕРИАЛ: – АТФ,")

Слайд 18Транскрибируемая нить
Нетранскрибируемая нить
РНК-полимераза
ДНК
пре-РНК (первичный транскрипт)
Биосинтез РНК
мононуклеотиды
Сайт терминации
Биосинтез РНКмононуклеотидыСайт терминации")
")
")


Слайд 23 Компоненты белоксинтезирующей системы
мРНК
20 Аминокислот
20 Аминоацил-тРНК синтетаз (АРС-аз)
Изоакцепторные тРНК
Рибосомы в
Источники энергии (АТФ, ГТФ) и Мg2+
Белковые факторы регуляции: факторы инициации, элонгации, терминации
Специальные ферменты посттрансляционного процессинга полипептидной цепи
Изоакцепторные тРНКРибосомы в виде полисомИсточники энергии (АТФ,")
Слайд 24Ядерные стадии биосинтеза белка
Биосинтез мРНК (транскрипция)
Созревание мРНК (посттранскрипционный процессинг)
Созревание мРНК (посттранскрипционный процессинг)")
Слайд 25Функции т-РНК
Акцепторная: связывает аминокислоту и транспортирует её к месту синтеза белка
Адаптерная:



Слайд 28Характеристика генетического кода
Триплетность (1 аминокислота кодируется 3 нуклеотидами)
Специфичность (каждому кодону соответствует
Вырожденность (1 аминокислота может кодироваться более чем 1 триплетом)
Линейная запись (прочтение кода «без знаков препинания»)
Универсальность (одинаков для всех живых существ)
Наряду со значимыми есть и «бессмысленные» кодоны (терминирующие – УАА, УАГ, УГА)
Колинеарность (соответствие линейной последовательности кодонов гена и последовательности АК в кодируемом белке)
Специфичность (каждому кодону соответствует только 1 аминокислота)Вырожденность (1")

Слайд 30Цитоплазматические стадии биосинтеза белка
Активация аминокислот, или образование аминоацил-тРНК
Инициация
Элонгация
Терминация
Посттрансляционная модификация



Слайд 33Функции активной рибосомы
Связывание и удерживание белоксинтезирующей системы
Транслокация – перемещение м-РНК через
Замыкание пептидной связи между аминокислотами (каталитическая функция)



Слайд 36Инициация:
К малой субъединице рибосомы прикрепляется м-РНК
К инициирующему кодону прикрепляется Мет-т-РНК
К образовавшемуся


Слайд 38Элонгация:
В пептидильном центре находится Мет-т-РНК, а в аминоацильном – аа-тРНК, соответствующая
Мет отрывается от т-РНК и перебрасывается в аминоацильный центр
С помощью пептидилтрансферазы замыкается пептидная связь между карбоксильной группой Мет и аминогруппой второй аминокислоты (требуется энергия ГТФ и регуляторный фактор Т)
Происходит передвижение рибосомы по м-РНК на один триплет (транслокация)
В освободившийся аминоацильный центр входит аа-т-РНК, соответствующая третьему кодону и т.д.



Слайд 41Посттрансляционный процессинг
Модификация N-конца полипептидной цепи
Фолдинг (формирование пространственной структуры)
Химическая модификация (гидроксилирование, гликозилирование
Присоединение простетических групп (у гетеропротеинов)
Объединение протомеров при образовании олигомерных белков
Присоединение сигнальных пептидов для выхода белка из клетки
Химическая модификация (гидроксилирование, гликозилирование и др.)Присоединение простетических групп")
Слайд 42Регуляция биосинтеза
1: РНК полимераза, 2: Репрессор, 3: Промотор,


Слайд 44Регуляция биосинтеза
Энхансеры – участки ДНК, присоединение к которым регуляторных белков усиливает
Сайленсеры – участки ДНК, присоединение к которым регуляторных белков уменьшает транскрипцию


Слайд 46Типы генов в геноме
Структурные гены (кодируют белки)
Регуляторные гены:
Гены-регуляторы (регулируют работу
Процессинг-гены (регулируют посттранскрипционные и посттрансляционный процессинг)
Темпоральные гены (включают в работу структурные гены в ходе клеточной дифференцировки)
Регуляторные гены: Гены-регуляторы (регулируют работу структурных генов)Процессинг-гены (регулируют посттранскрипционные")







")
")
Слайд 56
Полиморфизм белков –
изменение первичной структуры белка в пределах




Слайд 60
КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ
КАФЕДРА ФУНДАМЕНТАЛЬНОЙ И КЛИНИЧЕСКОЙ БИОХИМИИ
Лекция по БИОХИМИИ
тема:
«Обмен
Краснодар
2016



Слайд 63
Классификация простых липидов
Простые липиды: сложные эфиры жирных кислот с различными спиртами
Ацилглицеролы
Воска - сложные эфиры одноатомных или двухатомных длиноцепочечных спиртов и высших жирных кислот
Стериды - сложные эфиры циклического спирта холестерола и высших жирных кислот
")











")

глицерол")






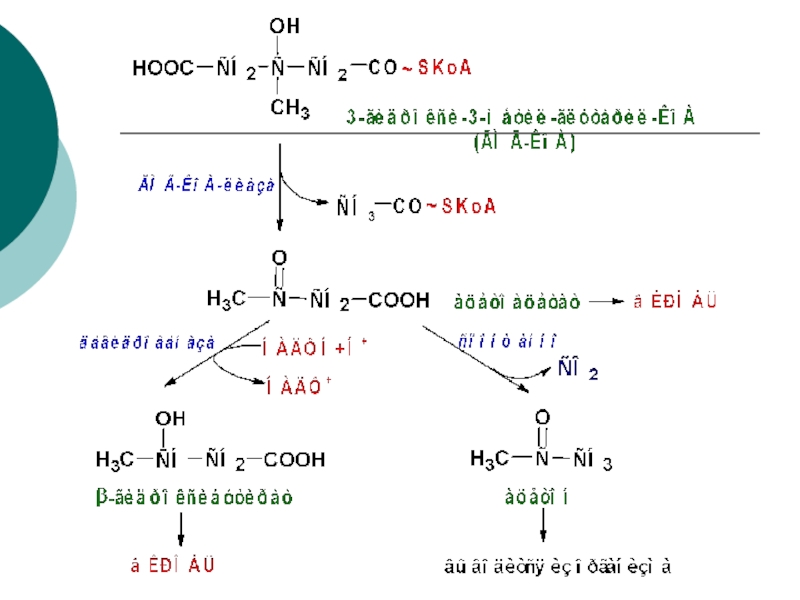
Слайд 86ОБЩАЯ СХЕМА ЦИКЛА β–ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ
дегидрогеназа
гидратаза
дегидрогеназа
тиолаза
2 АТФ
3 АТФ
12 АТФ

Слайд 87ЭНЕРГЕТИЧЕСКИЙ ИТОГ
n – количество С-атомов в жирной кислоте;
n/2 – количество молекул ацетил-КоА, образованных в процессе β-окисления;
12 – количество АТФ, синтезирующихся при окислении ацетил-КоА в ЦТК;
(n/2 – 1) – количество циклов β-окисления;
5 – количество молекул АТФ, образованных в каждом цикле за счёт двух реакций дегидрирования;
1 – затрата 1 молекулы АТФ на активацию жирной кислоты