- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Ліпіди. Загальна характеристика ліпідів презентация
Содержание
- 1. Ліпіди. Загальна характеристика ліпідів
- 2. ПЛАН 1. Загальна характеристика ліпідів 2. Обмін ліпідів
- 3. 1.ЗАГАЛЬНА ХАРАКТЕРИСТИКА ЛІПІДІВ Ліпі́ди — це
- 4. ФУНКЦІЇ ЛІПІДІВ Жири є формою запасання енергії,
- 5. КЛАСИФІКАЦІЯ ЛІПІДІВ Розрізняють дві групи ліпідів: прості
- 7. ПОДІЛІ ЇХ НА ВІСІМ ОСНОВНИХ ГРУП Жирні
- 8. 2. ОБМІН ЛІПІДІВ Обмін ліпідів складається з
- 9. В залежності від віку, фізичного навантаження, кліматичних
- 10. ПЕРЕТРАВЛЮВАННЯ ЛІПІДІВ. Більшість ліпідів корму засвоюється організмом
- 11. Корми, що містять ліпіди, механічно подрібнюються, перемішуються,
- 12. У процесі травлення в шлунково-кишковому тракті ліпіди
- 13. За хімічною структурою жовчні кислоти відносяться до
- 15. За фізико-хімічними властивостями жовчні кислоти є амфіфільними
- 16. З шлунка кормові маси дрібними порціями надходять
- 17. ВСМОКТУВАННЯ ЛІПІДІВ. Більшість ліпідів всмоктується в
- 18. Ліпідні речовини та продукти їх розщеплення проникають
- 19. У товстій кишці немає ферментів, що чинять
- 20. ДЛЯ ЗАБЕЗПЕЧЕННЯ НОРМАЛЬНОГО ТРАВЛЕННЯ І ВСМОКТУВАННЯ ПРОДУКТІВ
- 21. ПРОМІЖНИЙ ОБМІН. У ліпідів він має
- 22. РЕСИНТЕЗ ЛІПІДІВ У ТОНКІЙ КИШЦІ. В
- 23. ЛІПІДИ КРОВІ. Різні органи і тканини
- 24. ВИДИ ТРАНСПОРТУВАННЯ ЛІПІДІВ: з допомогою хіломікронів, ліпопротеїдів, вільних жирних кислот.
- 25. ОБМІН ЛІПІДІВ У ПЕЧІНЦІ Основна маса ліпідів
- 26. Обмін ліпідів у жирових депо. Основна
- 27. Кінцевий обмін. Основні кінцеві продукти ліпідного
- 28. Переварювання жирів. Основна маса жирів (95-97%) перетравлюється
- 30. β-моногліцериди можуть частково всмоктуватися стінкою кишкивника, йти
- 31. ЖИРИ При повному окисленні жиру в тканинах
- 32. 3. Біосинтез ліпідів Біосинтез жирів складається з
- 33. Утворення гліцерину. Близько 60 % гліцерину утворюється
- 34. Частина гліцерину утворюється в результаті гідролізу гліцеридів клітин і міжклітинної рідини під впливом ліпаз:
- 35. У подальшому гліцерин при наявності АТФ і
- 36. УТВОРЕННЯ ВИЩИХ ЖИРНИХ КИСЛОТ. Розрізняють два
- 37. МІТОХОНДРІАЛЬНИЙ Відбувається в мітохондріях клітин багатьох
- 38. НЕМІТОХОНДРІАЛЬНИЙ Відбувається в гіалоплазмі різних клітин. У
- 39. На першому етапі біосинтезу відбувається проникнення CO в молекулу біотин-ферменту під впливом ферменту карбоксилази:
- 40. Потім СОа переноситься на ацетил-КоА, що приводить
- 41. Якщо при біосинтезі утворюється пальмітинова кислота, то хімізм взаємодії ацетил-КоА і малоніл-КоА такий:
- 42. Біосинтез вищих жирних кислот відбувається з допомогою
- 43. УТВОРЕННЯ ТРИГЛІЦЕРИДІВ -здійснюється в більшості тканин.
- 44. У подальшому фосфатидна кислота під впливом ферменту фосфа-тидатфосфогідролази перетворюється на α-, β-дигліцерид:
- 45. α, β-Дигліцерид під впливом того самого мультиферментного комплексу взаємодіє з ацил-КоА, утворюючи тригліцерид:
- 46. БІОСИНТЕЗ СТЕРИНІВ І СТЕРЇДІВ. Стериди тканин
- 48. Незначна частина холестерину в тканинах синтезується з
- 49. ЛІПОЛІЗ Ліполізом називається процес ферментативного розщеплення ліпідів
- 50. ПЕРЕТВОРЕННЯ ЖИРІВ. Під впливом тканинних ліпаз,
- 51. ОБМІН ГЛІЦЕРИНУ. Після гідролізу жиру гліцерин
- 52. ОБМІН ВИЩИХ ЖИРНИХ КИСЛОТ. Механізм розпаду
- 53. Основою для створення теорії β-окислення послужили такі
- 54. СУЧАСНА СХЕМА Β-ОКИСЛЕННЯ ЖИРНИХ КИСЛОТ: 1. Під
- 55. 2. Ациладенілат взаємодіє з HS-KoA під дією
- 56. 4. Залишок жирної кислоти під дією еноїл-КоА-гідратази
- 57. 5. β-Оксіацил-КоА під впливом β-оксіацил-КоА-дегідрогенази при наявності НАД+ дегідрується, перетворюючись на кетоформу ацил-КоА;
- 58. 6. Молекула кетоформи ацил-КоА під впливом ферменту
- 59. Ацил-КоА знову зазнає β-окислення, доки вся молекула
- 60. Вищі жирні кислоти — важливе джерело хімічної
- 61. 4. Регуляція ліпідного обміну. Обмін ліпідів
- 62. Окремі гормони сприяють накопиченню організмом ліпідів (наприклад,
- 63. 5. ПАТОЛОГІЯ ЛІПІДНОГО ОБМІНУ Ліпідний обмін
- 64. При зниженій продуктивності ліполітичних ферментів травними залозами
- 65. Ліпідний обмін порушується при кетозах. Вони виникають
- 67. Частина ацетооцтової кислоти в печінці лід впливом
- 68. Частина ацетооцтової кислоти ферментативним (під впливом ацетоацилдекарбоксилази)
- 69. У крові здорових тварин кетонові тіла містяться
- 70. Часто трапляються порушення холестеринового обміну. У хворих

Слайд 2ПЛАН
1. Загальна характеристика ліпідів
2. Обмін ліпідів
3. Біосинтез ліпідів.
4. Регуляція ліпідного обміну.
5. Патологія ліпідного обміну.

Слайд 31.ЗАГАЛЬНА ХАРАКТЕРИСТИКА ЛІПІДІВ
Ліпі́ди — це група органічних речовин, що входять до

Слайд 4ФУНКЦІЇ ЛІПІДІВ
Жири є формою запасання енергії, фосфоліпіди та стероїди входять до

Слайд 5КЛАСИФІКАЦІЯ ЛІПІДІВ
Розрізняють дві групи ліпідів: прості і складні.
прості (естери жирних
складні (такі, що крім залишку жирної кислоти та спирту містять ще додаткові групи: вуглеводні, фосфатні та інші)
 ;")
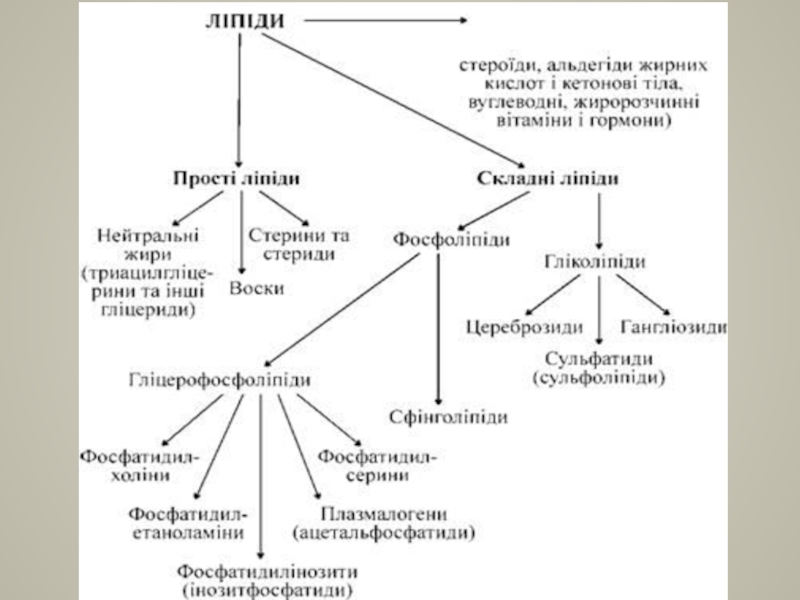
Слайд 7ПОДІЛІ ЇХ НА ВІСІМ ОСНОВНИХ ГРУП
Жирні кислоти (FA);
Гліцероліпіди (GL);
Гліцерофосфоліпіди (GP);
Сфінголіпіди (SP);
Стероїдні
Пренольні ліпіди (PR);
Сахароліпіди (SL);
Полікетиди (PK).
Холестерин
;Гліцероліпіди (GL);Гліцерофосфоліпіди (GP);Сфінголіпіди (SP);Стероїдні ліпіди (ST);Пренольні ліпіди (PR);Сахароліпіди")
Слайд 82. ОБМІН ЛІПІДІВ
Обмін ліпідів складається з чотирьох етапів:
перетравлювання,
всмоктування,
Проміжний обмін,
кінцевий

Слайд 9В залежності від віку, фізичного навантаження, кліматичних умов потреба в ліпідах
При окисленні 1 г. жиру виділяється енергії в 2 р. більше, ніж при окисленні 1 г. вуглеводів або білків. Таким чином, саме ліпіди забезпечують від 1/3 до половини загальної кількості калорій середньої дієти людини.

Слайд 10ПЕРЕТРАВЛЮВАННЯ ЛІПІДІВ.
Більшість ліпідів корму засвоюється організмом тільки після попереднього розщеплення. Під

Слайд 11Корми, що містять ліпіди, механічно подрібнюються, перемішуються, змочуються слиною і перетворюються
У шлунку кормові маси знаходяться від 4 до 12 год. Шлунковий сік містить ліпазу, здатну гідролітичне розщеплювати емульгований жир.

Слайд 12У процесі травлення в шлунково-кишковому тракті ліпіди зазнають ферментативного гідролізу до
Оскільки жири у воді нерозчинні, а ферменти-білки нерозчинні у жирі, то реакція гідролізу молекул жиру відбувається тільки на межі розподілу між ліпідною краплею і водною фазою. Тому, чим вищий ступінь емульгування жиру, тобто чим менші окремі краплі жиру, тим більша величина доступної поверхні. Основну емульгуючу дію виконують жовчні кислоти. Емульгуванню жирів сприяють також перистальтика кишечника, білки, СО2 і моноацилгліцерини.

Слайд 13За хімічною структурою жовчні кислоти відносяться до стероїдних сполук, похідних холанової
Синтезуються жовчні кислоти у печінці із холестерину.
Функціонують жовчні кислоти не у вільній формі, а у вигляді кон'югованих сполук із гліцином чи таурином.

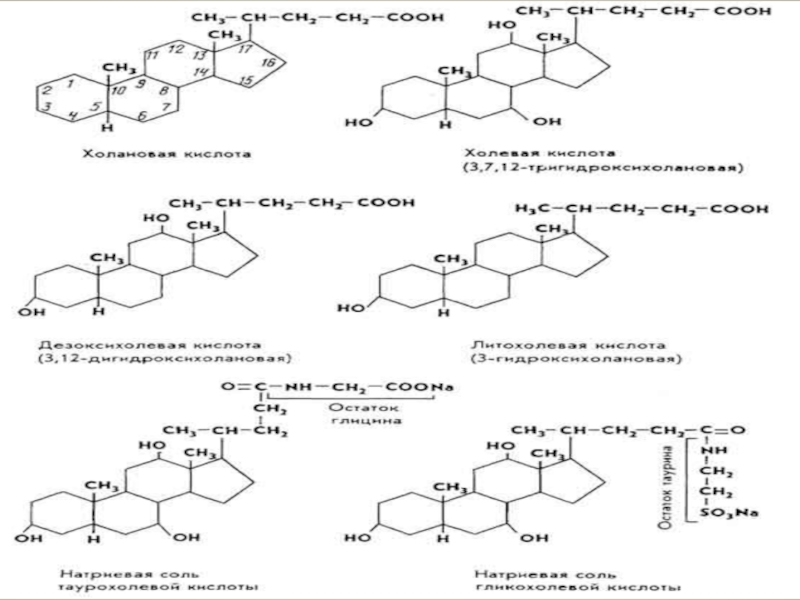
Слайд 15За фізико-хімічними властивостями жовчні кислоти є амфіфільними речовинами, у яких циклічна
Ліпаза утворюється у підшлунковій залозі у формі проферменту – проліпази. У дванадцятипалій кишці проліпаза перетворюється в активну ліпазу завдяки приєднанню білка коліпази і дії жовчних кислот.

Слайд 16З шлунка кормові маси дрібними порціями надходять у дванадцятипалу кишку, потім
завершується перетравлювання
ліпідів і
відбувається всмоктування продуктів їх розщеплення.

Слайд 17ВСМОКТУВАННЯ ЛІПІДІВ.
Більшість ліпідів всмоктується в нижній частині дванадцятипалої і у

Слайд 18Ліпідні речовини та продукти їх розщеплення проникають у клітину покривного епітелію
Кінцеві продукти травлення ліпідів розподіляються у двох фазах: ліпідній і міцелярній.
У ліпідній фазі основними компонентами є найдрібніші часточки три- і дигліцеридів, у міцелярній — вищі жирні кислоти, моногліцериди та інші продукти травлення ліпідів.

Слайд 19У товстій кишці немає ферментів, що чинять гідролітичну дію на ліпіди.
Ліпідні

Слайд 20ДЛЯ ЗАБЕЗПЕЧЕННЯ НОРМАЛЬНОГО ТРАВЛЕННЯ І ВСМОКТУВАННЯ ПРОДУКТІВ РОЗПАДУ ЛІПІДІВ МАЄ ЗНАЧЕННЯ
1) секреція підшлунковою залозою гідролітичних ферментів, які каталізують розрив складноефірних зв'язків;
2) надходження жовчних кислот, які емульгують жири і забезпечують всмоктування продуктів їх гідролізу;
3) захоплення продуктів травлення ліпідів клітинами слизової оболонки кишечника;
4) перетворення продуктів травлення у частинки для транспорту від клітин слизової у лімфатичні судини і далі – в кров.
 секреція")
Слайд 21ПРОМІЖНИЙ ОБМІН.
У ліпідів він має особливості, які полягають у тому,

Слайд 22РЕСИНТЕЗ ЛІПІДІВ У ТОНКІЙ КИШЦІ.
В епітеліальних клітинах слизової оболонки відбувається

Слайд 23ЛІПІДИ КРОВІ.
Різні органи і тканини одержують ліпіди та продукти їх

Слайд 24ВИДИ ТРАНСПОРТУВАННЯ ЛІПІДІВ:
з допомогою хіломікронів,
ліпопротеїдів,
вільних жирних кислот.

Слайд 25ОБМІН ЛІПІДІВ У ПЕЧІНЦІ
Основна маса ліпідів після проходження через печінку стає

Слайд 26Обмін ліпідів у жирових депо.
Основна маса ліпідів та їхніх похідних,

Слайд 27Кінцевий обмін.
Основні кінцеві продукти ліпідного обміну — вуглекислий газ і

Слайд 28Переварювання жирів.
Основна маса жирів (95-97%) перетравлюється в тонкому кишківнику. Переварювання складається
 перетравлюється в тонкому кишківнику. Переварювання складається з двох процесів: емульгування")
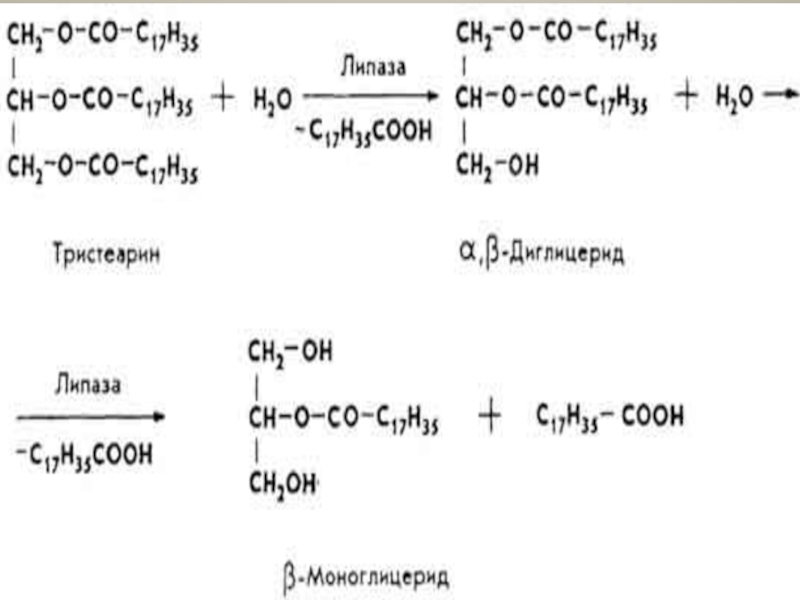
Слайд 30β-моногліцериди можуть частково всмоктуватися стінкою кишкивника, йти на ресинтез тригліцеридів організму

Слайд 31ЖИРИ
При повному окисленні жиру в тканинах утворюються вуглекислий газ і вода.
Дихальний коефіцієнт (співвідношення між кількістю вуглекислого газу, який утворився під час окислення жиру, і кількістю кисню, необхідного для окислення) тут дорівнює 0,71.

Слайд 323. Біосинтез ліпідів
Біосинтез жирів складається з трьох основних етапів: утворення гліцерину,

Слайд 33Утворення гліцерину. Близько 60 % гліцерину утворюється з продуктів проміжного обміну

Слайд 34Частина гліцерину утворюється в результаті гідролізу гліцеридів клітин і міжклітинної рідини

Слайд 35У подальшому гліцерин при наявності АТФ і під впливом фермен¬ту гліцеролкінази
Фосфорилування гліцерину відбувається у більшості тканин в організмі, крім жирової тканини і слизової оболонки тонкої кишки, в якій немає ферменту гліцеролкінази.

Слайд 36УТВОРЕННЯ ВИЩИХ ЖИРНИХ КИСЛОТ.
Розрізняють два типи біосинтезу вищих жирних кислот:

Слайд 37 МІТОХОНДРІАЛЬНИЙ
Відбувається в мітохондріях клітин багатьох органів (печінки, мозку, серця, нирок)
 і деяких тканин (жирової).")
Слайд 38НЕМІТОХОНДРІАЛЬНИЙ
Відбувається в гіалоплазмі різних клітин. У ньому беруть участь ферменти карбоксилази,

Слайд 39На першому етапі біосинтезу відбувається проникнення CO в молекулу біотин-ферменту під

Слайд 40Потім СОа переноситься на ацетил-КоА, що приводить до утворення малоніл-КоА:
Далі йде

Слайд 41Якщо при біосинтезі утворюється пальмітинова кислота, то хімізм взаємодії ацетил-КоА і

Слайд 42Біосинтез вищих жирних кислот відбувається з допомогою ацилпереносного білка (АПБ), що
, що містить простетичну групу, подібну")
Слайд 43УТВОРЕННЯ ТРИГЛІЦЕРИДІВ
-здійснюється в більшості тканин.
Джерелом для утворення є гліцерофосфат
Реакція каталізується мультиферментним комплексом — гліцерофосфат-ацилтрансферазою, локалізованою в мікросомах.

Слайд 44У подальшому фосфатидна кислота під впливом ферменту фосфа-тидатфосфогідролази перетворюється на α-,

Слайд 45α, β-Дигліцерид під впливом того самого мультиферментного комплексу взаємодіє з ацил-КоА,

Слайд 46БІОСИНТЕЗ СТЕРИНІВ І СТЕРЇДІВ.
Стериди тканин синтезуються з екзогенного (який надходить
 і ендогенного")
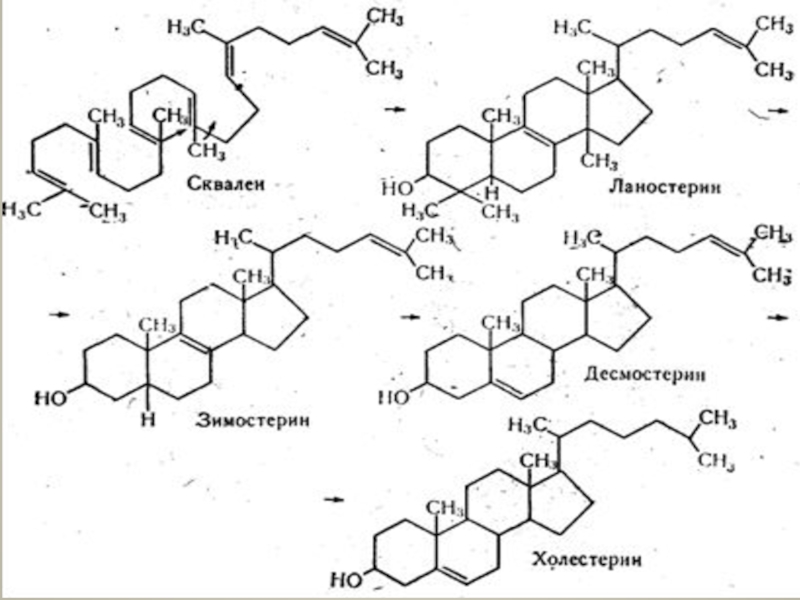
Слайд 48Незначна частина холестерину в тканинах синтезується з деяких амінокислот (лейцину).
Холестерин характеризується
.Холестерин характеризується високим ступенем обміну. Так,")
Слайд 49ЛІПОЛІЗ
Ліполізом називається процес ферментативного розщеплення ліпідів клітин до їх складових частин,

Слайд 50ПЕРЕТВОРЕННЯ ЖИРІВ.
Під впливом тканинних ліпаз, активованих солями фосфорної кислоти, нейтральні

Слайд 51ОБМІН ГЛІЦЕРИНУ.
Після гідролізу жиру гліцерин під впливом ферменту гліцеролкінази фосфорилується.

Слайд 52ОБМІН ВИЩИХ ЖИРНИХ КИСЛОТ.
Механізм розпаду вищих жирних кислот в організмі

Слайд 53Основою для створення теорії β-окислення послужили такі факти. Наприкінці XIX ст,

Слайд 54СУЧАСНА СХЕМА Β-ОКИСЛЕННЯ ЖИРНИХ КИСЛОТ:
1. Під впливом аденілаткінази жирна кислота активується

Слайд 552. Ациладенілат взаємодіє з HS-KoA під дією ферменту ацил-КоА-синтетази з утворенням
3. Під впливом флавінзалежної дегідрогенази відбувається дегідрування активованого залишку жирної кислоти;

Слайд 564. Залишок жирної кислоти під дією еноїл-КоА-гідратази гідрується в ділянці розриву

Слайд 575. β-Оксіацил-КоА під впливом β-оксіацил-КоА-дегідрогенази при наявності НАД+ дегідрується, перетворюючись на

Слайд 586. Молекула кетоформи ацил-КоА під впливом ферменту ацето-ацетил-КоА-тіолази і при наявності

Слайд 59Ацил-КоА знову зазнає β-окислення, доки вся молекула жирної кислоти не розпадається

Слайд 60Вищі жирні кислоти — важливе джерело хімічної енергії в організмі. При

Слайд 614. Регуляція ліпідного обміну.
Обмін ліпідів регулюється нейрогуморальним шляхом. Центральна нервова

Слайд 62Окремі гормони сприяють накопиченню організмом ліпідів (наприклад, інсулін—жирів, ліпокаїн — фосфатидів),
Високий вміст у кормах вуглеводів і жирів сприяє надлишковому накопиченню жиру. При нестачі в кормах метіоніну — джерела метальних груп для холіну,-порушується ліпідний обмін і настає патологічне ожиріння печінки.
, інші стимулюють їх розщеплення.")
Слайд 63 5. ПАТОЛОГІЯ ЛІПІДНОГО ОБМІНУ
Ліпідний обмін порушується при багатьох інфекційних, інвазійних

Слайд 64При зниженій продуктивності ліполітичних ферментів травними залозами і зменшенні секреції жовчі
Відсутність у кормах ліпотропних речовин (холіну і метіоніну) викликає жирове переродження печінки, нестача жиророзчинних вітамінів зменшує всмоктування, знижує секрецію жовчі, веде до появи дерматитів, проносів.
 порушу¬ються процеси")
Слайд 65Ліпідний обмін порушується при кетозах. Вони виникають при цукровому діабеті, гепатитах,

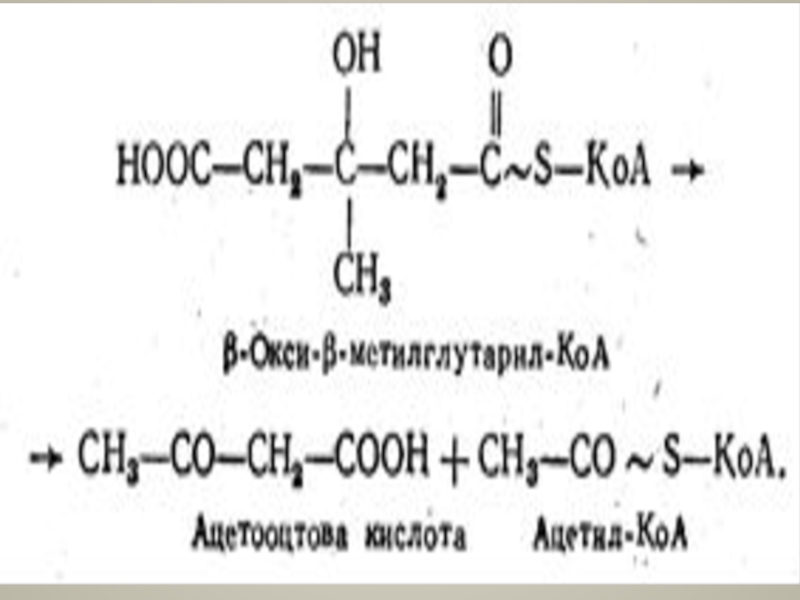
Слайд 67Частина ацетооцтової кислоти в печінці лід впливом НАД-залежної р-оксибутиратдегідрогенази відновлюється до

Слайд 68Частина ацетооцтової кислоти ферментативним (під впливом ацетоацилдекарбоксилази) і неферментативним шляхами декарбоксилується,
 і неферментативним шляхами декарбоксилується, що приводить до утворення ацетону:")
Слайд 69У крові здорових тварин кетонові тіла містяться в невеликих кількостях, поглинаються
У жуйних більша частина кетонових тіл утворюється в епітелії передшлунків, менша — в тканинах печінки. У корів під час лактації збільшується вміст кетонових тіл у крові.
 і")
Слайд 70Часто трапляються порушення холестеринового обміну. У хворих виникає надлишок холестерину в






