- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Лекция 8. Молекулярная генетика. Транскрипция РНК эукариот презентация
Содержание
- 1. Лекция 8. Молекулярная генетика. Транскрипция РНК эукариот
- 2. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ Словарик
- 3. Словарик Фосфорилирование – процесс переноса остатка фосфорной
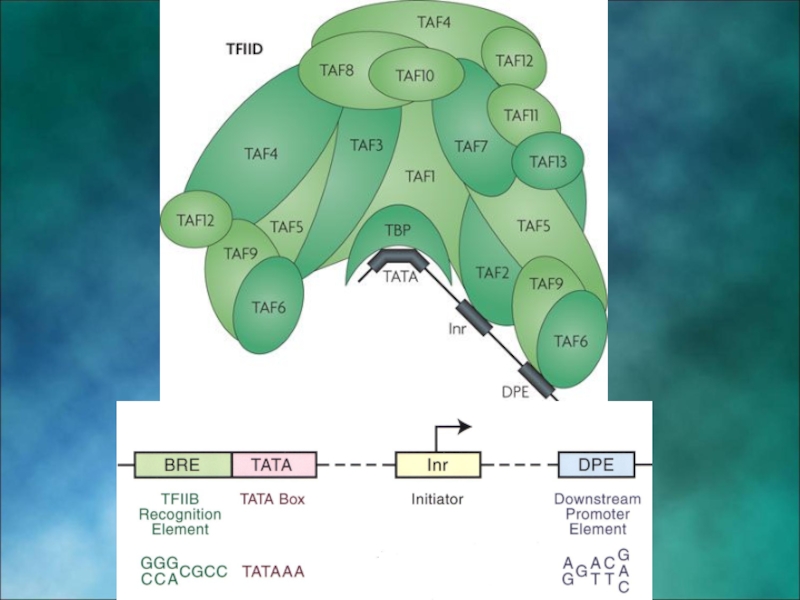
- 5. Обозначения структурных элементов транскриптона
- 6. Особенности транскриптона эукариот
- 7. Структура промотора эукариот На расстоянии -25
- 8. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ Факторы транскрипции эукариот
- 9. Основные ДНК-связывающие белки
- 10. Ядерные РНК-полимеразы
- 11. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ РНК-полимераза I
- 12. РНК-полимераза I. Функции РНК
- 13. У млекопитающих промотор гена предшественника rРНК состоит
- 14. В состав SL1 входит белок TBP (TATA-box
- 15. Факторы регулирующие транскрипцию rРНК, ассоциированы с РНК-полимеразой
- 16. Транскрипционная активность РНК полимеразы I зависит от:
- 17. РНК-полимераза I должна быть фосфорилирована для успешного
- 18. Активность основных факторов инициации (SL1 и
- 19. Концентрация активного UBF регулируется связыванием с белком,
- 20. Белок TTFI способствует передвижению нуклеосом по ДНК
- 21. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ РНК-полимераза III
- 22. РНК-полимераза III. Функции. РНК полимераза
- 23. Ген 5S rРНК содержит внутренний промотор, области
- 24. TFIIIA, 5S rРНК специфичный транскрипционный фактор, взаимодействует
- 25. TFIIIC связывает А-последовательность ДНК, без этого невозможна
- 26. В состав TFIIIB входит TBP TFIIIB фактор
- 27. Гены tРНК содержат дополнительный участок ДНК –
- 28. Регуляторные элементы промотора генов snРНК расположены
- 29. С PSE связывается белок PBP С DSE
- 30. Промотор содержит TATA-бокс. Промоторы snРНК, транскрибируемых
- 31. Терминация транскрипции для РНК полимеразы III изучена
- 32. Работа РНК полимеразы III, регулируется в
- 33. Фосфорилирование направлено на подавление транскрипции во
- 34. Транскрипция РНК полимеразой III чувствительна к структуре хроматина Регуляция транскрипции гена snРНК
- 35. Субъединица 131 kD фактора TFIIIC способна
- 36. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ РНК-полимераза II
- 37. РНК-полимераза II. Функции транскрибирует mРНК,
- 40. Основные стадии транскрипции
- 43. Инициация транскрипции «в комиксах» ДНК К цепи
- 45. Инициация транскрипции «в комиксах» ТВР связывается с
- 46. Инициация транскрипции + Pol II «в
- 47. Инициация транскрипции + Pol II «в
- 48. Инициация транскрипции + Pol II «в
- 50. Элонгация транскрипции «в комиксах» В двухцепочечной
- 51. Элонгация транскрипции «в комиксах» Элонгация mРНК
- 52. Терминация транскрипции «в комиксах» Комплекс Pol
- 54. Ключевая стадия инициации транскрипции РНК полимеразой II
- 55. Большая субъединица белка TFIIF фосфорилируется.
- 56. До начала синтеза РНК, С-концевой домен РНК
- 57. TFIIH - комплекс, важный для репарации
- 58. После фосфорилирования С-концевого домена CTD, РНК полимераза
- 59. Помимо CTD, TFIIH фосфорилирует (активируя) несколько
- 60. Кроме описанных факторов в инициации транскрипции
- 61. Основной маркер элонгационного состояния РНК полимеразы II
- 62. Кроме pTEFb, фактором элонгации РНК полимеразы II
- 63. В процессе элонгации РНК полимераза может встретить
- 64. Если работающая РНК-полимераза встретит повреждение ДНК, нуждающееся
- 65. За супрессию пауз РНК-полимеразы отвечает TFIIF, а
- 66. Кроме факторов, обеспечивающих элонгацию транскрипции, с фосфорилированной
- 67. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ Регуляция транскрипции
- 68. Некодирующие регуляторные элементы Геном животных и человека
- 69. Энхансеры. Структура. Энхансеры - это последовательности ДНК,
- 70. Энхансеры: «петлевая модель» действия Активаторы (Activators) -
- 71. САЙЛЕНСЕРЫ
- 72. Инсуляторы (англ. insulate - изолировать )
- 73. Механизм действия инсуляторов А Схема функционирования двух генов
- 74. Механизм действия инсуляторов В Если пограничный элемент, инсулятор


Слайд 3Словарик
Фосфорилирование – процесс переноса остатка фосфорной кислоты от фосфорилирующего агента-донора к
Ацетилирование – замещение атомов водорода в органических соединениях остатком уксусной кислоты CH3CO (ацетильной группой), катализируется ацетилтрансферазами
Энхансеры – последовательности ДНК, усиливающие транскрипцию при взаимодействии со специфическими белками
Сайленсеры – последовательности ДНК репрессирующие активность генов, ослабляющие транскрипцию при взаимодействии с белками
Инсуляторы – последовательности ДНК, определяющие специфичность действия энхансеров и сайленсеров



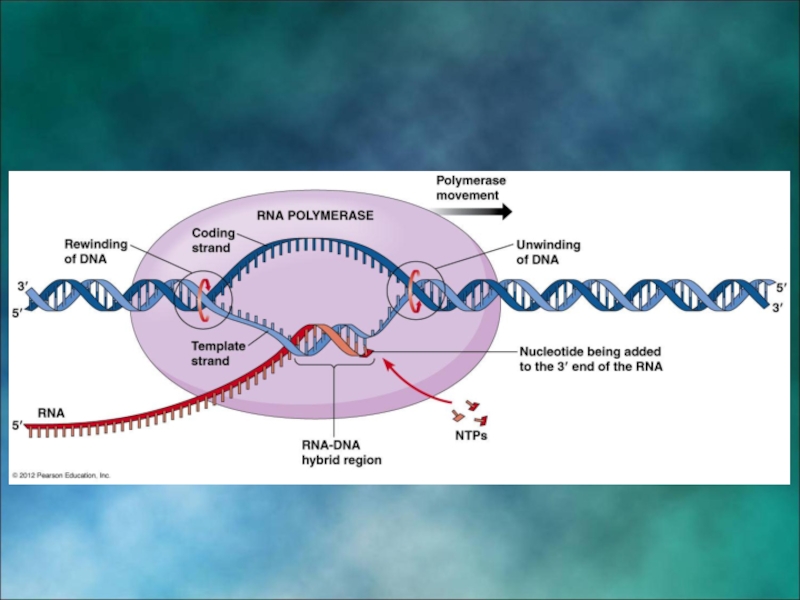
Слайд 7Структура промотора эукариот
На расстоянии -25 п.н. от +1 нукл. находится ТАТА-бокс,
На расстоянии -60-80 п.н. находится ЦААТ-бокс
Расстояние между ЦААТ и ТАТА большое и РНК-полимераза не способна накрыть всю эту область. ЦААТ опознается своим белком, а ТАТА - своим.





Слайд 12РНК-полимераза I. Функции
РНК полимераза I транскрибирует гены 18S, 5.8S
Транскрипция генов rРНК и сборка рибосом происходит в ядрышке

Слайд 13У млекопитающих промотор гена предшественника rРНК состоит из двух участков:
CPE (core promoter element)
UCE (upstream control element).
С этими участками связываются белковые комплексы SL1 и UBF
Структура регуляторных элементов гена rРНК
")
Слайд 14В состав SL1 входит белок TBP (TATA-box binding protein), хотя в
В SL1 входят также три дополнительных белка, называемых TAFI110, TAFI63 и TAFI48.
РНК полимераза I и белки, регулирующие транскрипцию rРНК
, хотя в промоторе rРНК нет ТАТА")
Слайд 15Факторы регулирующие транскрипцию rРНК, ассоциированы с РНК-полимеразой I
Белок TIF-IA связывает
PAF53 - другой белок млекопитающих, ассоциированный с полимеразой, взаимодействует с UBF
РНК полимераза I и белки, регулирующие транскрипцию rРНК

Слайд 16Транскрипционная активность РНК полимеразы I зависит от: 1. фазы клеточного цикла
2. пролиферативной активности клеток (при интенсивном делении требуется много rРНК).
Регуляция транскрипции гена rРНК
; 2.")
Слайд 17РНК-полимераза I должна быть фосфорилирована для успешного взаимодействия с TIF-IA
Регуляция

Слайд 18Активность основных факторов инициации
(SL1 и UBF) регулируется в зависимости от
1. SL1 инактивируется в митозе при помощи фосфорилирования.
2. UBF активируется фосфорилированием в G1 фазе.
Регуляция транскрипции гена rРНК
 регулируется в зависимости от фазы клеточного цикла: 1.")
Слайд 19Концентрация активного UBF регулируется связыванием с белком, ассоциированным с ретинобластомой Rb
Его ингибирующая активность подавляется при фосфорилировании в S фазе.
Подобным образом p53 инактивирует TBP, входящий в состав SL1
Регуляция транскрипции гена rРНК

Слайд 20Белок TTFI способствует передвижению нуклеосом по ДНК для эффективной транскрипции РНК
Регуляция транскрипции гена rРНК




Слайд 24TFIIIA, 5S rРНК специфичный транскрипционный фактор, взаимодействует с C последовательностью внутри
Это приводит к посадке белкового комплекса – TFIIIC
Факторы транскрипции гена 5S rРНК

Слайд 25TFIIIC связывает А-последовательность ДНК, без этого невозможна эффективная транскрипции гена.
После
Факторы транскрипции гена 5S rРНК

Слайд 26В состав TFIIIB входит TBP
TFIIIB фактор способствует присоединению РНК полимеразы III
Регуляция транскрипции гена 5S rРНК

Слайд 27Гены tРНК содержат дополнительный участок ДНК – В, способствующий связыванию TFIIIC.
Белок-белковое взаимодействие TFIIIA и TFIIIC заменяется ДНК-белковым.
Далее, TFIIIC присоединяет TFIIIB и РНК полимеразу III
Факторы транскрипции гена tРНК

Слайд 28Регуляторные элементы промотора генов snРНК
расположены вне транскрибируемой области – PSE
Факторы транскрипции гена snРНК

Слайд 29С PSE связывается белок PBP
С DSE – фактор Oct-1.
Связывание Oct-1
Факторы транскрипции гена snРНК

Слайд 30Промотор содержит TATA-бокс.
Промоторы snРНК, транскрибируемых РНК полимеразой III схожи с
Для всех трех типов промоторов общий механизм - это посадка TFIIIB на промотор при помощи ДНК-связывающих белков-активаторов
Факторы транскрипции гена snРНК

Слайд 31Терминация транскрипции для РНК полимеразы III изучена плохо.
Известно, что транскрипты
способствующий процессингу и стабильности транскриптов
Факторы транскрипции гена snРНК

Слайд 32
Работа РНК полимеразы III, регулируется в зависимости от фазы клеточного цикла
Регуляция

Слайд 33
Фосфорилирование направлено на подавление транскрипции во время митоза.
La белок активируется
Регуляция транскрипции гена snРНК

Слайд 34
Транскрипция РНК полимеразой III чувствительна к структуре хроматина
Регуляция транскрипции гена

Слайд 35
Субъединица 131 kD фактора TFIIIC способна ацетилировать гистоны и, таким образом,
Регуляция транскрипции гена snРНК


Слайд 37РНК-полимераза II. Функции
транскрибирует mРНК, а также некоторые sРНК
наиболее подверженный
состоит из 17 субъединиц, одной из особенностей строения наибольшей субчастицы РНК полимеразы II является ее С-концевой домен CTD, построенный из повторяющихся гептамеров. Количество этих повторов варьирует для разных организмов, составляя в среднем несколько десятков.
Последовательное укорочение С-концевого домена приводит к гибели клеток.

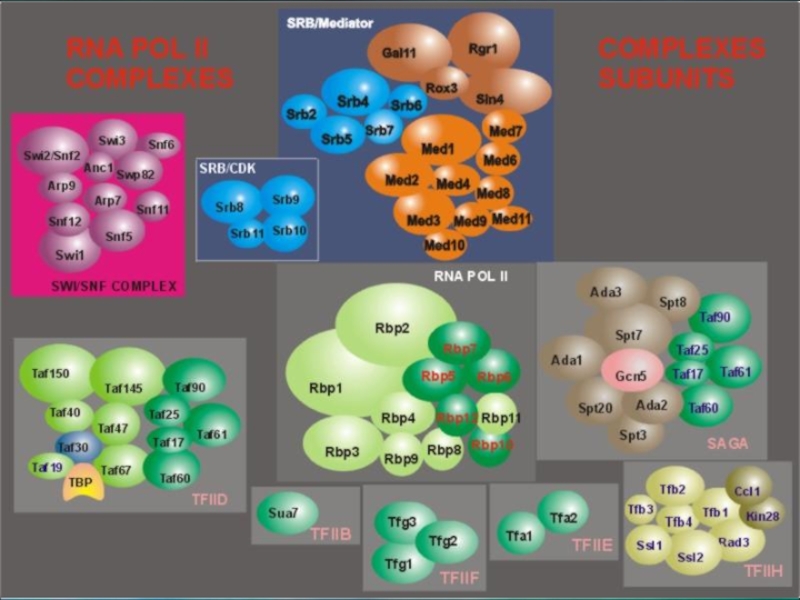
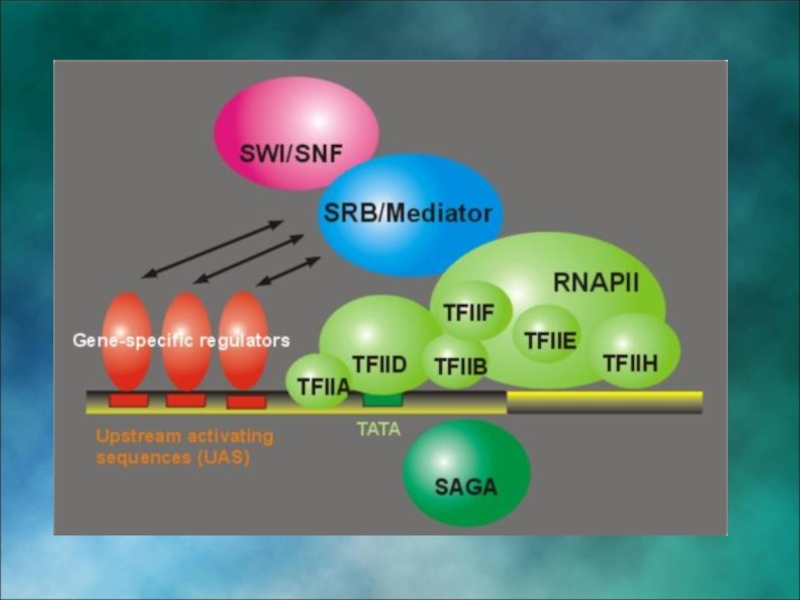

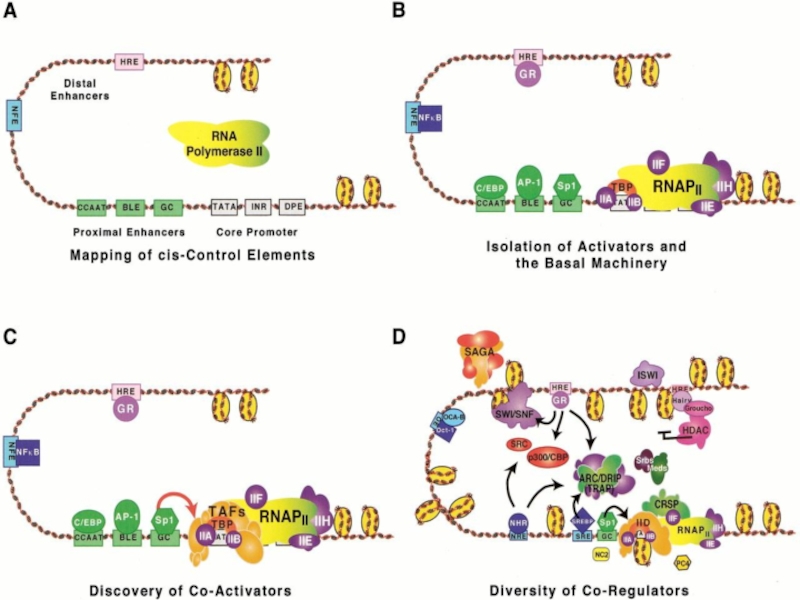
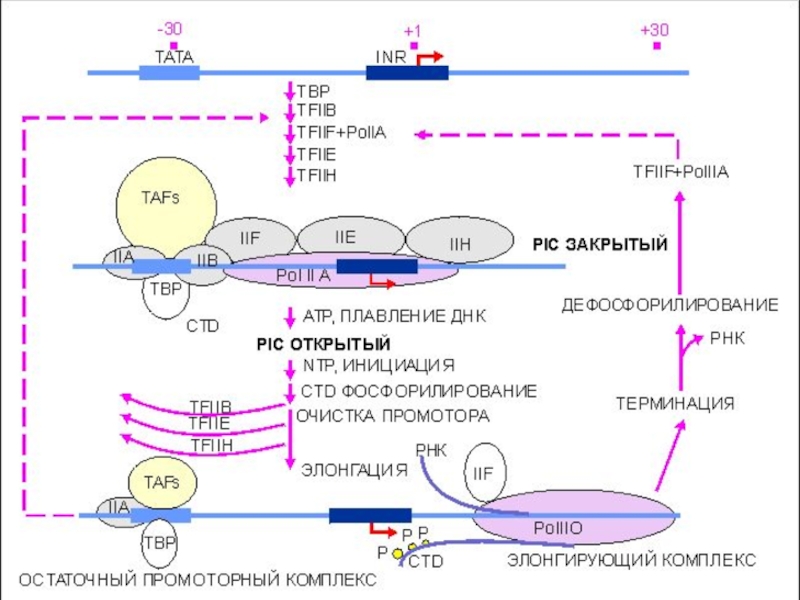
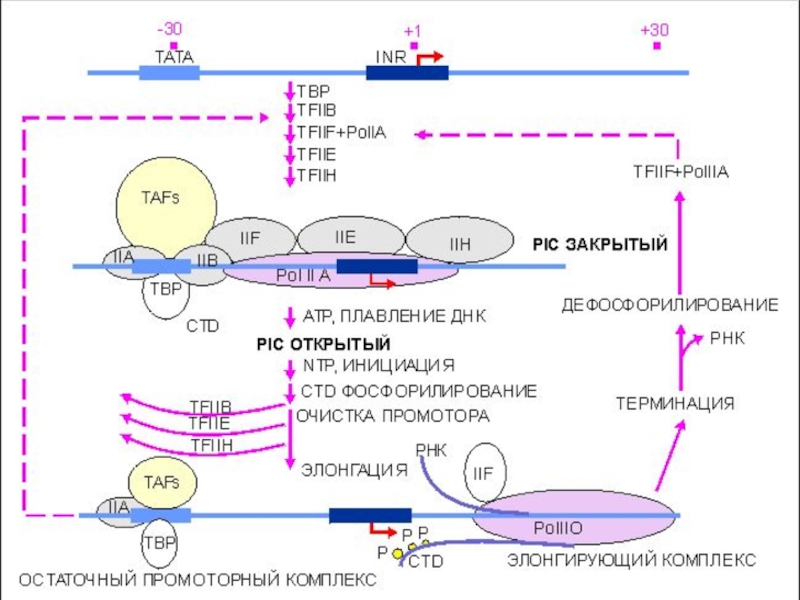
Слайд 43Инициация транскрипции «в комиксах»
ДНК
К цепи подходит фактор транскрипции TFIID
TFIID самый крупный

Слайд 45Инициация транскрипции «в комиксах»
ТВР связывается с ТАТА боксом и располагается рядом
Это присоединение вызывает формирование изгиба ДНК

Слайд 46Инициация транскрипции + Pol II
«в комиксах»
Затем подходят факторы TFIIA (оранжевый)
TFIIA присоединяется к ТАТА рядом с TFIID
TFIIB подходит к ТАТА.
Рol II собирается дистанционно
 и TFIIB (красный)TFIIA присоединяется")
Слайд 47Инициация транскрипции + Pol II
«в комиксах»
TFIIB связывается с ТАТА и
Pol II собран и подходит к точке инициации транскрипции
К сформировавшемуся комплексу подходит фактор TFIIE

Слайд 48Инициация транскрипции + Pol II
«в комиксах»
TFIIE
TFIIH
TFIIE связывается, подходит TFIIH
Все готово

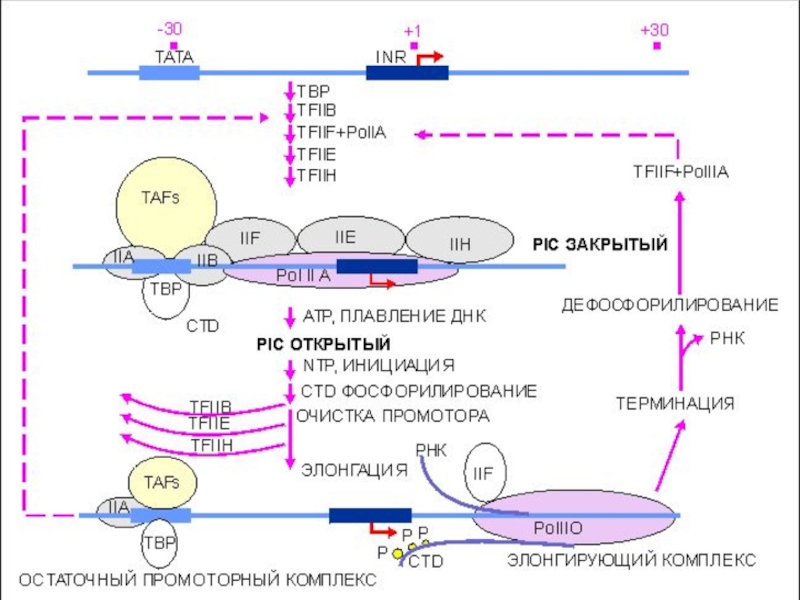
Слайд 50Элонгация транскрипции
«в комиксах»
В двухцепочечной ДНК формируется «глаз», обеспечивающий доступ Pol
Начинается элонгация цепи и факторы TFIIE, TFIIH, TFIIA, TFIIB отсоединяются
TFIIE действует дистанционно и mРНК быстро растет

Слайд 51Элонгация транскрипции
«в комиксах»
Элонгация mРНК завершается когда достигается конец транскрипта
Закрывается «глаз»
Pol II освобождается вместе с полученными от АТФ фосфатами

Слайд 52Терминация транскрипции
«в комиксах»
Комплекс Pol II отходит, mРНК освобождается. Фактор TFIID
Первичный транскрипт готов и после процессинга будет транспортирован из ядра клетки

Слайд 54Ключевая стадия инициации транскрипции РНК полимеразой II – это посадка на
а именно на TATA бокс, TBP и ассоциированных с ним факторов
Инициация транскрипции mРНК

Слайд 55
Большая субъединица белка TFIIF фосфорилируется.
Эта субъединица является ATP-зависимой ДНК хеликазой
Инициация

Слайд 56До начала синтеза РНК, С-концевой домен РНК CTD полимеразы II дефосфорилирован.
После связывания РНК полимеразы II, с инициаторным комплексом связывается TFIIE, который, в свою очередь, присоединяет TFIIH
Инициация транскрипции mРНК

Слайд 57
TFIIH - комплекс, важный для репарации и транскрипции.
В его состав
Инициация транскрипции mРНК

Слайд 58После фосфорилирования С-концевого домена CTD, РНК полимераза теряет сродство в инициаторному
Инициация транскрипции mРНК

Слайд 59
Помимо CTD, TFIIH фосфорилирует (активируя) несколько циклин-зависимых киназ: cdk2, 4 и
Инициация транскрипции mРНК
 несколько циклин-зависимых киназ: cdk2, 4 и 6, а также TFIIE")
Слайд 60
Кроме описанных факторов в инициации транскрипции РНК полимеразой II участвует множество
Они обеспечивают регуляцию транскрипции различных генов.

Слайд 61Основной маркер элонгационного состояния РНК полимеразы II -фосфорилированный CTD.
За поддержание
Инактивация этого фактора влияет только на транскрипцию определенного набор генов.
Элонгация транскрипции mРНК

Слайд 62Кроме pTEFb, фактором элонгации РНК полимеразы II является гетеротример элонгин А.
Негативный регулятор этого фактора – VHL. Он связывает комплекс элонгинов B и C и предотвращает их ассоциацию с элонгином А
Элонгация транскрипции mРНК

Слайд 63В процессе элонгации РНК полимераза может встретить те же проблемы, что
Элонгация транскрипции mРНК

Слайд 64Если работающая РНК-полимераза встретит повреждение ДНК, нуждающееся в репарации, то с
Эти белки опосредуют посадку факторов репарации на повреждение, а также играют важную роль для восстановления транскрипции после репарации.
Элонгация транскрипции mРНК

Слайд 65За супрессию пауз РНК-полимеразы отвечает TFIIF, а за выход из арестованного
Элонгация транскрипции mРНК

Слайд 66Кроме факторов, обеспечивающих элонгацию транскрипции, с фосфорилированной формой РНК полимеразы II
Элонгация транскрипции mРНК


Слайд 68Некодирующие регуляторные элементы
Геном животных и человека – сложнейшая система с многоуровневой системой
Важную роль в процессах регуляции играют некодирующие последовательности: энхансеры, сайленсеры, инсуляторы. Они определяют уровень транскрипции генов.

Слайд 69Энхансеры. Структура.
Энхансеры - это последовательности ДНК, формирующие модули.
Модули - это
Например, М1+М2+М3+М4 - один энхансер, но он состоит из 4-х модулей.
Все 4 модуля узнаются своими белками, а они, сидя на ДНК, взаимодействуют друг с другом.
Если в клетке присутствуют все соответствующие белки, то участку ДНК придается определенная конформация и начинается синтез РНК.

Слайд 70Энхансеры: «петлевая модель» действия
Активаторы (Activators) - белки, связывающиеся с энхансерами, которые
 - белки, связывающиеся с энхансерами, которые помогают РНК-полимеразе правильно начать")

Слайд 72Инсуляторы (англ. insulate - изолировать )
Последовательности нуклеотидов, которые могут подавлять позитивное
Такие участки ДНК как бы изолируют ген, находящийся между ними, способствуя сохранению его обычной пространственной структуры.
Последовательности нуклеотидов, которые могут подавлять позитивное и негативное влияние эухроматина")
Слайд 73Механизм действия инсуляторов
А Схема функционирования двух генов (желтый и голубой), локализованных в
, локализованных в домене, контролируемом двумя инсуляторами")
Слайд 74Механизм действия инсуляторов
В Если пограничный элемент, инсулятор (красный), расположен между двумя энхансерами,
Бирюзовый транскрипционный фактор не может действовать на промотор темно-желтого гена в соответствии с локализацией второго инсулятора.
Энхансер может активировать транскрипцию с промотора голубого гена, который остался в том же домене.
, расположен между двумя энхансерами, формируется новый хромосомный домен,")