- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Лекция 8 (А. П. Перевозчиков)ЭВОЛЮЦИЯ ДЕТЕРМИНАЦИИ ПОЛА У ЖИВОТНЫХ презентация
Содержание
- 1. Лекция 8 (А. П. Перевозчиков)ЭВОЛЮЦИЯ ДЕТЕРМИНАЦИИ ПОЛА У ЖИВОТНЫХ
- 2. Регуляция пола может осуществляться у животных различным
- 3. Зависящее от температуры (инкубации яиц) определение пола
- 4. Пути эволюции механизмов определения пола у животных
- 5. Молекулярные механизмы детерминации пола, в виде сигнальных цепочек, контролируемых определенными генами
- 6. Изначально, представления о детерминации пола было связано
- 7. Схематическое представление генетической программы детерминации пола Drosophila,
- 8. Гены нумераторы (числителя), локализованные на X-хромосоме, кодируют
- 9. В дальнейшем, когда sxl начинает транскрибироваться с
- 10. Каскады регуляций, контролирующих формирование половых структур самки
- 11. Альтернативный сплайсинг -основной путь реализации программы детерминации пола у дрозофилы (последовательность событий)
- 12. Роли DsxM и DsxF в формировании пола
- 13. Формирование мужской и женской половых гонад у дрозофилы под контролем различных форм Dsx
- 14. Эволюция регуляции пола у насекомых способом ретроградной надстройки
- 15. Эволюция детерминации пола у двукрылых, Sanchez, 2008
- 16. КОНСЕРВАТИЗМ ЦЕПОЧЕК ВЗАИМОДЕЙСТВИЙ МАКРОМОЛЕКУЛ МОЖЕТ БЫТЬ
- 17. Для механизмов эволюции детерминации пола было
- 18. Нематода Caenorhabditis elegans имеет два пола: XX
- 19. Детерминация пола у С. elegans. Определяющим
- 20. Сигнальная цепочка детерминации пола C. elegans. Перемещение
- 24. Генетическая гомология и функциональное сходство белков Dsx (дрозофила) Mab-3 (нематоды) (Zhu et al., 2000)
- 25. У нематод в 1998 году был
- 26. Ортологи генов Sox 9 активно экспрессируются
- 27. Детерминация пола у млекопитающих При анализе Y-хромосомы
- 30. Мышь с половым генотипом XX, трансгенная по
- 31. Образование гонад связывают с экспрессией в клетках-предшественницах
- 32. Проникновение (миграция) мезонефротических клеток (любого генотипа XY
- 35. Молекулярные механизмы детерминации пола у млекопитающих (Sekido, Lovell-Badge, 2008)
- 36. Хромосомная детерминация пола у млекопитающих (Graves, 2008)
- 37. Fergusson-Smith, 2007
- 38. На 9-й хромосоме человека (короткое плечо)
- 39. Возможное участие гена DMRT в формировании семенников и сперматогенезе у млекопитающих
- 40. Сколько раз в эволюции возникали механизмы детерминации пола? Гипотеза:
- 41. Скорости эволюции генов (определяются по соотношению несинонимичных
- 42. Fergusson-Smith, 2007
- 43. Различные модели детерминации пола (учитывающие лишь молекулярные механизмы)
- 44. Возможность существования консервативных факторов детерминации пола у разных животных
- 45. Распространённость генов DM (Volff et al., 2003)
- 46. Хронология образования различных групп позвоночных, Graves, 2008
- 47. Эволюция половых хромосом Есть ли эволюционная преемственность между половыми хромосомами у позвоночных?
- 48. Детерминация пола у позвоночных, Ezaz et al., 2006
- 49. Слоистость половых хромосом, выявляемая при прекращениии рекомбинации между ними (Bergero, Charlesworth, 2008)
- 51. Различные (включая хромосомные) способы детерминации пола у позвоночных, Graves, 2008
- 52. Эволюция пар половых хромосом в пределах классов позвоночных, (Graves, 2008)
- 53. Хронология образования различных групп позвоночных, Graves, 2008
- 54. Преемственность в хромосомной детерминации пола у амниот
- 55. Хромосомная преемственность детерминации пола у позвоночных ограничена, Graves, 2008
- 56. Факторы внешней среды и гормональный контроль детерминации пола
- 57. У эхиуриды Bonellia (организма, обитающего на морском
- 58. У улитки- блюдечка Crepidula fornicata, новая особь,
- 59. Половых гормонов, по-видимому, нет у беспозвоночных
- 60. Гинандроморфы дрозофилы и мотылька, образовавшиеся в результате
- 61. Синтез стероидных половых гормонов у позвоночных
- 62. Половые гормоны, продуцируемые сигнальными цепочками
- 63. Зависящее от температуры (инкубации яиц) определение пола
- 64. Факторы внешней среды, определяющие формирование пола у
- 65. Мужские и женские стероидные гормоны. Эстрогены (включая
- 66. Виды женских половых гормонов и их функции
- 67. Районы мужской половой системы, формирующиеся под контролем тестостерона и дидротестостерона.
- 68. Заключение
 ЭВОЛЮЦИЯ ДЕТЕРМИНАЦИИ ПОЛА У ЖИВОТНЫХ")
Слайд 2Регуляция пола может осуществляться у животных различным образом благодаря генетическим факторам
(GSD) или (и) факторам внешней среды (TSD)
 или (и) факторам")
Слайд 3Зависящее от температуры (инкубации яиц) определение пола у трех представителей класса
пресмыкающихся: грифовая (Macroclemis) и красноухая (Trachemys) черепахи и аллигатор.
 определение пола у трех представителей класса пресмыкающихся: грифовая (Macroclemis) и")
Слайд 4Пути эволюции механизмов определения пола у животных
Молекулярные механизмы детерминации пола, в
виде сигнальных цепочек, контролируемых определенными генами
Формирование половых хромосом, для которых вырабатываются блоки рекомбинации
Факторы внешней среды, влияющие на тип пола
Появление специфичных половых гормонов
Формирование половых хромосом, для которых вырабатываются блоки рекомбинации
Факторы внешней среды, влияющие на тип пола
Появление специфичных половых гормонов

Слайд 5
Молекулярные механизмы детерминации пола, в виде сигнальных цепочек, контролируемых определенными генами

Слайд 6Изначально, представления о детерминации пола было связано исключительно с хромосомами Хромосомная детерминация
пола у Drosophila
Особи генотипа XX -самки и генотипа XY -самцы. В случае генотипа X0 -также образуются самцы, но стерильные
У дрозофилы (и у многих других насекомых) нет гормонального контроля формообразования пола. Пол определяется соотношением числа X-хромосом к числу наборов аутосом - X:A. В случае X:A =1 -пол женский, в случае X:A = 0, 5 -пол мужской..
Балансовая гипотеза определения пола К. Бриджеса.

Слайд 7Схематическое представление генетической программы детерминации пола Drosophila, определяемое соотношением X:A. Детерминация
пола по мужскому или женскому типу определяется по цепочке генов: sex-lethal, transformer, doublesex, последовательно регулирующих друг друга путём альтернативного сплайсинга. Формирующийся в конечном счёте фактор транскрипции doublesex существует в виде двух вариантов - dsxF(самки) dsxM (самца), что и определяет будущий пол.

Слайд 8Гены нумераторы (числителя), локализованные на X-хромосоме, кодируют факторы транскрипции sisterless-a, sisterless-b=scute
(sis-a,sis-b), runt (runt), активирующие транскрипцию гена sxl с проксимального промотора. Гены деноминаторы (знаменателя), локализованные на аутосомах, кодируют факторы транскрипции deadpan (dpn), daughterless (da), extramacrohaeta (emc) и др., которые репрессируют активацию гена sex-lethal (sxl) с проксимального промотора. Активаторы (белки-нумераторы)и репрессоры (белки-деноминаторы) способны комплексировать и решающим событием в активации sxl является преодоление возможности репрессоров угнетать действие активаторов. В случае активации sxl c раннего (ближнего) промотора (при отношении X:A=1) (генотип -XX) альтернативный сплайсинг осуществляется с образованием функционально активного белкового продукта sxl, который, являясь фактором сплайсинга, взаимодействует с собственной пре-иРНКи закрепляет этот же вариант сплайсинга sxl для дальнейших целей. В случае отношения X:A=0,5 (генотип XY/X0) в транскрипционном комплексе преобладают репрессоры, sxl не транскрибируется с раннего (проксимального) промотора.
, локализованные на X-хромосоме, кодируют факторы транскрипции sisterless-a, sisterless-b=scute (sis-a,sis-b), runt (runt), активирующие")
Слайд 9В дальнейшем, когда sxl начинает транскрибироваться с позднего (дистального) промотора (при
обоих вариантах полового генотипа) альтернативный сплайсинг sxl при генотипе XY/X0 идет с сохранением третьего экзона, в котором находится стоп-кодон. В результате чего белковый продукт sxl у самца - дефектен. Он не способен функционировать как фактор сплайсинга. Напротив белковый продукт sxl самки (XX)-функционально активный фактор сплайсинга не только для своей РНК, но и для пре-иРНК гена transformer (tra), что предопрделяет образование белка tra как следующего фактора сплайсинга, модифицирующего (совместно с другим белком- tra2) сплайсинг следующего в каскаде гена – doublesex (dsx). В результате образуется фактор транскрипции -DsxF (фенотипа самки). В отсутствии активного tra-белка сплайсинг dsx проходит по другому варианту и образуется видоизмененный фактор транскрипции DsxM (фенотипа самца).
 промотора (при обоих вариантах полового генотипа)")
Слайд 10Каскады регуляций, контролирующих формирование половых структур самки и самца дрозофилы Различная
активация гена sxl (sex-lethal) будущих самок и самцов дрозофилы. Деятельность генов нумераторов и деноминаторов определяет альтернативные транскрипцию и сплайсинг sxl.
")
Слайд 11Альтернативный сплайсинг -основной путь реализации программы детерминации пола у дрозофилы
(последовательность событий)
")
Слайд 12Роли DsxM и DsxF в формировании пола у дрозофилы. Dsx- транскрипционный фактор,
действующий как репрессор или как активатор, для каждого пола в альтернативной манере




Слайд 16
КОНСЕРВАТИЗМ ЦЕПОЧЕК ВЗАИМОДЕЙСТВИЙ МАКРОМОЛЕКУЛ МОЖЕТ БЫТЬ ОТНОСИТЕЛЬНО ПОЛНЫЙ ИЛИ НЕПОЛНЫЙ. ВО
ВТОРОМ СЛУЧАЕ ВОЗМОЖНО ДОБАВЛЕНИЕ РЕГУЛЯТОРНЫХ ЭТАПОВ (СТУПЕНЕЙ) ЛИБО СВЕРХУ (UPSTREAM), ЛИБО СНИЗУ (DOWNSTREAM)

Слайд 17
Для механизмов эволюции детерминации пола было высказано две гипотезы
1. Сохранение полной
цепочки регуляции пола ( исходно для всех насекомых)
2. гипотеза ретроградной надстройки регуляторной цепочки (Wilkins, 2002).
В реальности существуют не цепочки, а трехмерные сети
2. гипотеза ретроградной надстройки регуляторной цепочки (Wilkins, 2002).
В реальности существуют не цепочки, а трехмерные сети

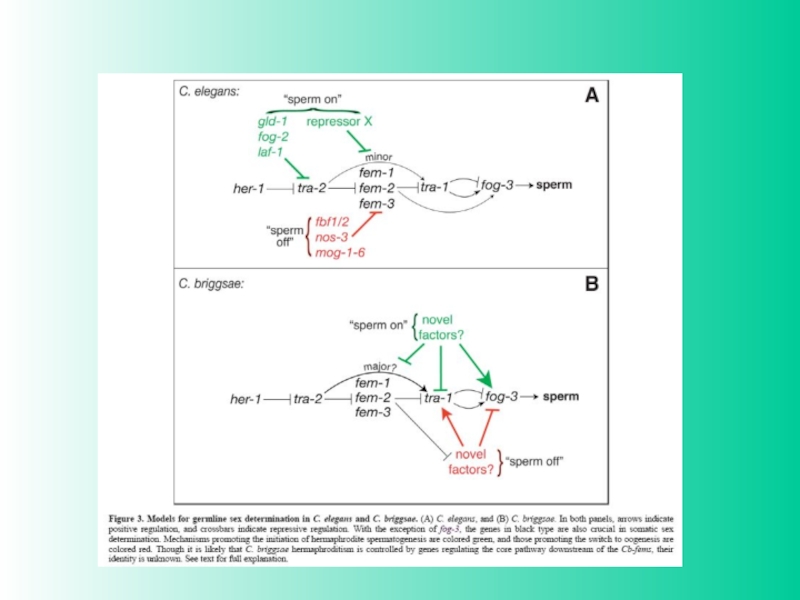
Слайд 18Нематода Caenorhabditis elegans имеет два пола: XX -гермафродиты (более 99% популяции)
и X0 -самцы. У взрослой особи (гермафродита) число соматических клеток тела- 959 (у первой личинки -558). Число половых клеток больше (более 1500). В случае самооплодотворения 0,2% новых особей -самцы, остальные -гермафродиты, в случае спаривания особей XX и X0 - половина новых особей - самцы.
 и X0 -самцы. У")
Слайд 19Детерминация пола у С. elegans. Определяющим моментом является активация гена tra-1
(не родственного гену tra дрозофилы). Соотношение X : A сканируется активностью гена xol-1. Высокие дозы белка xol-1 меняют характер регуляции в детерминации пола с активации (гермафродиты) на репрессию (самцы)

Слайд 20Сигнальная цепочка детерминации пола C. elegans. Перемещение в ядро транскрипционного фактора
TRA-1 означает реализацию гермафродитного фенотипа. Фактор Her-1 особи X0, взаимодействуя с мембрано-связанным белком TRA-2, вызывает детерминацию пола по мужскому типу. При этом промежуточный участник –белок FEM, удерживает TRA-1 в цитоплазме (диссоциирует с последним при детерминации пола по XX-гермафродитному пути.



Слайд 24Генетическая гомология и функциональное сходство белков Dsx (дрозофила) Mab-3 (нематоды) (Zhu
et al., 2000)
 Mab-3 (нематоды) (Zhu et al., 2000)")
Слайд 25
У нематод в 1998 году был описан ген Mab3, отвечающий за
формирования мужской гонады и следовательно самцов. Он выявляется в цепочке, отвечающей за детерминацию мужского пола
Его ДНК-связывающий домен гомологичен ДНК-связывающему домену белка Doublesex (мужской формы) насекомых
Его ДНК-связывающий домен гомологичен ДНК-связывающему домену белка Doublesex (мужской формы) насекомых

Слайд 26
Ортологи генов Sox 9 активно экспрессируются в клетках Сертоли у птиц
и пресмыкающихся (аллигаторы, ящерицы).
Экспрессирующийся ортолог гена Dmrt 1 был найден в семенниках птиц, пресмыкающихся (аллигаторы, черепахи) и некоторых рыб (Medaka, puffer fish), но не найден у других (zebrafish, telyapia).
Экспрессирующийся ортолог гена Dmrt 1 был найден в семенниках птиц, пресмыкающихся (аллигаторы, черепахи) и некоторых рыб (Medaka, puffer fish), но не найден у других (zebrafish, telyapia).
.Экспрессирующийся")
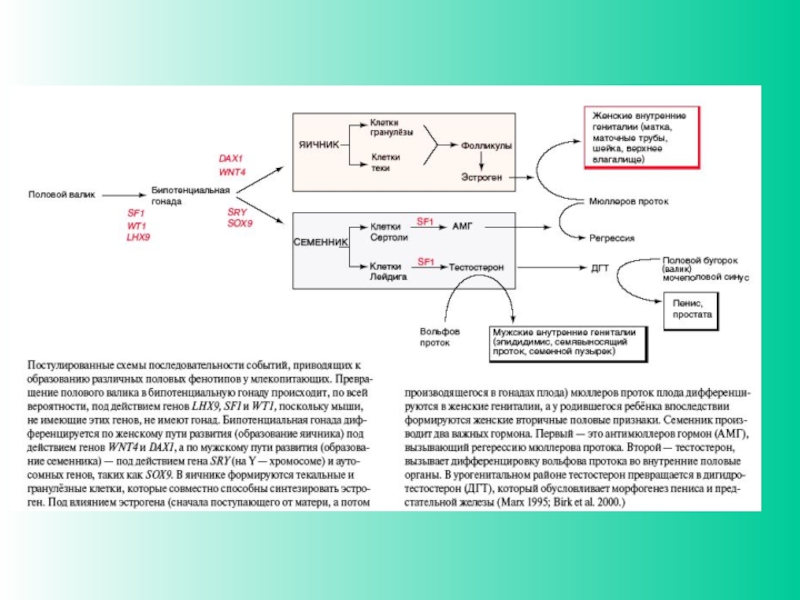
Слайд 27Детерминация пола у млекопитающих
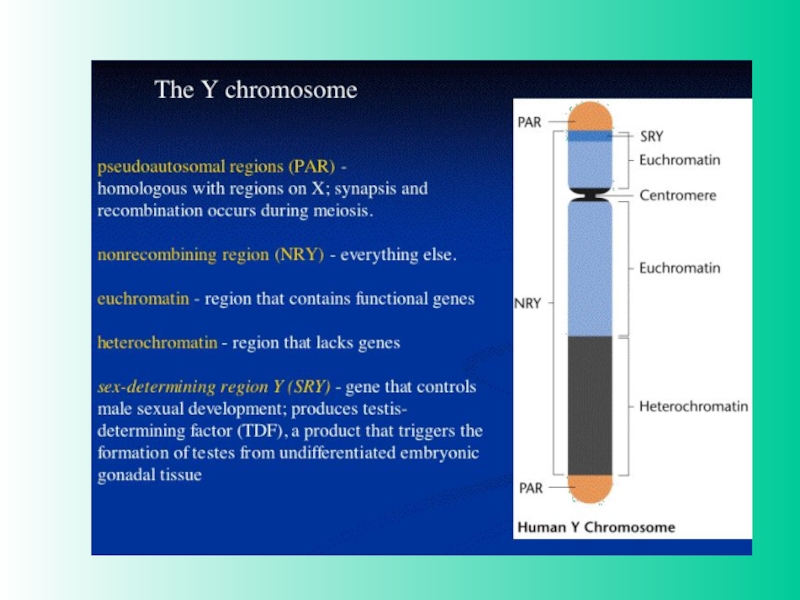
При анализе Y-хромосомы выяснилось, что гипотетический фактор, отвечающий
за формирование пола, существует в действительности- это ген SRY/Sry (sex region of Y-chromosome), локализован на коротком плече Y-хромосомы. Это транскрипционный фактор, он содержит HMG-домен и относится к семейству SOX-белков. На X-хромосоме гена SRY нет. Ген был найден также в редких случаях у лиц мужского пола с генотипом XX, которые содержали транслоцированный ген SRY. Для детерминации мужского фенотипа нужна также активность другого гена Sox-семейства - SOX9, локализованного на аутосоме (17 хромосома у человека). Мутация по нему вызывает превращение особей XY в самок или в гермафродитов. Гены SRY и SOX9 экспрессируются вместе в половых (гонадных) валиках самцов

Слайд 30Мышь с половым генотипом XX, трансгенная по гену SRY(справа), в норме
локализованным на Y-хромосоме обладает фенотипом самца
, в норме локализованным на Y-хромосоме обладает")
Слайд 31Образование гонад связывают с экспрессией в клетках-предшественницах соответствующих генов (Sry, Sox
9 или Wnt4, Dax1 и т.д.) (Sekido, Lovell-Badge, 2008)

Слайд 32Проникновение (миграция) мезонефротических клеток (любого генотипа XY или XX) в Sry+
гонадные валики (генотипа XY) in vitro, доказывает важность экспрессии гена Sry в клетках производных гонадных валиков для индуцирования ими образования связи Вольфова протока (через мезонефротические клетки) с половыми тяжами (будущими семенными канальцами), формирующимися из гонадных валиков
 мезонефротических клеток (любого генотипа XY или XX) в Sry+ гонадные валики (генотипа XY)")


")
")

Слайд 38
На 9-й хромосоме человека (короткое плечо) было идентифицировано 3 гена Dmrt
(1-3), родственные генам Dsx (дрозофилы) и Mab3 (С. elegans). Эти гены (особенно Dmrt1) важны для формирования семенников. Делеция участка 9-й хромосомы, содержащей гены Dmrt ведет к реверсии пола, как и делеция Sox9 (на 17-й хромосоме).
 было идентифицировано 3 гена Dmrt (1-3), родственные генам Dsx")


Слайд 41Скорости эволюции генов (определяются по соотношению несинонимичных и синонимичных замен аминокислот
в кодирующих последовательностях ортологичных генов близкородственных видов) (обычно средняя скорость в пределах 0,12-0,20)


")

")

Слайд 47Эволюция половых хромосом
Есть ли эволюционная преемственность между половыми хромосомами у позвоночных?


Слайд 49Слоистость половых хромосом, выявляемая при прекращениии рекомбинации между ними (Bergero, Charlesworth,
2008)
")
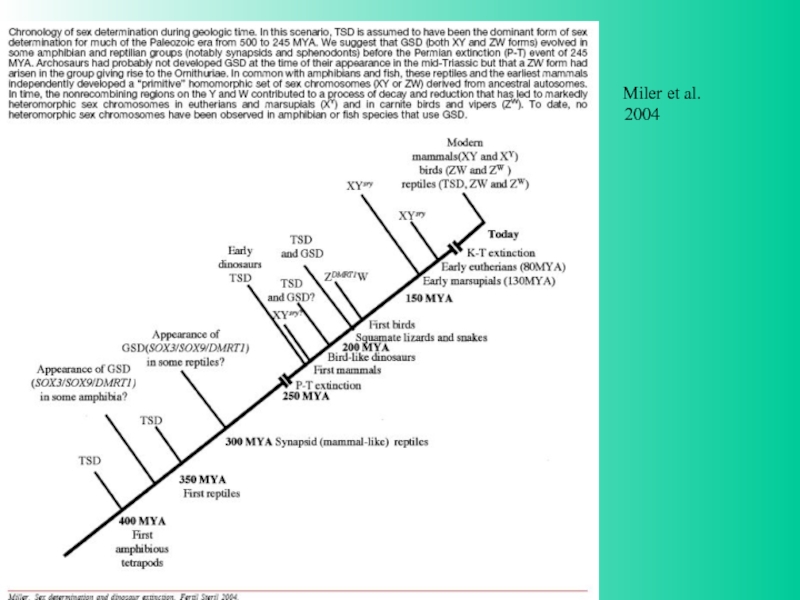
 способы детерминации пола у позвоночных, Graves, 2008")
")




Слайд 57У эхиуриды Bonellia (организма, обитающего на морском каменистом грунте) формирование пола
происходит у личинки в зависимости от места её прикрепления к поддерживающей среде. При осаждении на скалистый грунт формируется самка (тело - 10 см, ротовой хоботок -proboscis -около метра), при осаждении на proboscis самки - формируется самец (1-3 мм длиной), который мигрирует по кишечнику и обитает в матке (или нефридии), оплодотворяя яйца)
 формирование пола происходит у личинки в")
Слайд 58У улитки- блюдечка Crepidula fornicata, новая особь, располагающаяся в колонии сверху
- всегда самец. Затем мужская половая система дегенерирует и самец превращается в самку


Слайд 60Гинандроморфы дрозофилы и мотылька, образовавшиеся в результате утраты в раннем развитии
из ряда клеток одной их X-хромосом. В результате организм содержит частично женские, частично мужские структуры .


Слайд 62
Половые гормоны, продуцируемые сигнальными цепочками детерминации пола, способны сами влиять на
работу сигнальных цепочек и контролировать образование пола у животных с TSD

Слайд 63Зависящее от температуры (инкубации яиц) определение пола у трех представителей класса
пресмыкающихся: грифовая (Macroclemis) и красноухая (Trachemys) черепахи и аллигатор.
 определение пола у трех представителей класса пресмыкающихся: грифовая (Macroclemis) и")
Слайд 64Факторы внешней среды, определяющие формирование пола у позвоночных (пресмыкающихся), могут действовать,
через изменение баланса половых гормонов.
Мужские половые гормоны - андрогены (тестостерон и ряд других)
Женские половые гормоны - эстрогены .
Показано что фермент P450 ароматаза, катализирующий превращение тестостерона в эстроген, у пресмыкающихся способен менять свою активность в зависимости от температуры окружающей среды. Температура инкубации яиц в последнюю треть развития пресмыкающихся (черепах, крокодилов, аллигаторов) является определяющей для формирования пола выводящихся из яиц животных.
, могут действовать, через изменение баланса половых")
Слайд 65Мужские и женские стероидные гормоны. Эстрогены (включая диэтилстилбестрол) важны не только
для формирования женских половых органов -матки, шейки матки, влагалища и клитора -из структур мюллерова протока, но также для развития молочных желез. Диэтилстилбестрол важен для нормального протекания беременности.
У самцов эстрогены продуцируется в ряде тканей (включая мозг) и важны для фертильности спермы. Если концентрация эстрогенов в крови выше у самок, то концентрация их в канальцах семенников - rete testis ниже, чем в крови у самок. У самок также имеются андрогены
 важны не только для формирования женских половых")

Слайд 67Районы мужской половой системы, формирующиеся под контролем тестостерона и дидротестостерона.

Слайд 68Заключение
Сигнальные цепочки, принимающие участие в
формирования пола, могут сохраняться, по меньшей мере в виде фрагментов, у Bilateria. Гены, контролирующие их работу: DMRT1 и им подобные. Новые гены могут вовлекаться в эти цепочки, по-видимому, за счёт коопции.
Далеко не все (и не всегда) из этих важных генов локализованы на половых хромосомах. Пример, гены DMRT1 и Sox9.
Половые хромосомы, вероятно, возникают, не потому, что они сцеплены с генами, детерминирующими пол, а по другим причинам, важным для реализации репродукции.
Половые хромосомы возникают из аутосом и по меньшей мере для разных классов позвоночных неоднократно. Эволюционно развивается хромосомный диморфизм (например, в ряду: питоны, ужи , гадюки), способствующий видоспецифичному оплодотворению и ассимиляции мутаций в организме. Блок рекомбинации половых хромосом угрожает сохранению меньшей хромосомы (крайний результат у C. elegans, слепушонки и т.п.) и, возможно, свидетельствует о блоке в эволюции этих ветвей.
Половые гормоны влияют на детерминацию пола и могут, шунтируя работу половых хромосом, взять на себя ответственность за детерминацию пола (черепахи, крокодилы, аллигаторы, некоторые ящерицы), независимо от полового генотипа
Далеко не все (и не всегда) из этих важных генов локализованы на половых хромосомах. Пример, гены DMRT1 и Sox9.
Половые хромосомы, вероятно, возникают, не потому, что они сцеплены с генами, детерминирующими пол, а по другим причинам, важным для реализации репродукции.
Половые хромосомы возникают из аутосом и по меньшей мере для разных классов позвоночных неоднократно. Эволюционно развивается хромосомный диморфизм (например, в ряду: питоны, ужи , гадюки), способствующий видоспецифичному оплодотворению и ассимиляции мутаций в организме. Блок рекомбинации половых хромосом угрожает сохранению меньшей хромосомы (крайний результат у C. elegans, слепушонки и т.п.) и, возможно, свидетельствует о блоке в эволюции этих ветвей.
Половые гормоны влияют на детерминацию пола и могут, шунтируя работу половых хромосом, взять на себя ответственность за детерминацию пола (черепахи, крокодилы, аллигаторы, некоторые ящерицы), независимо от полового генотипа