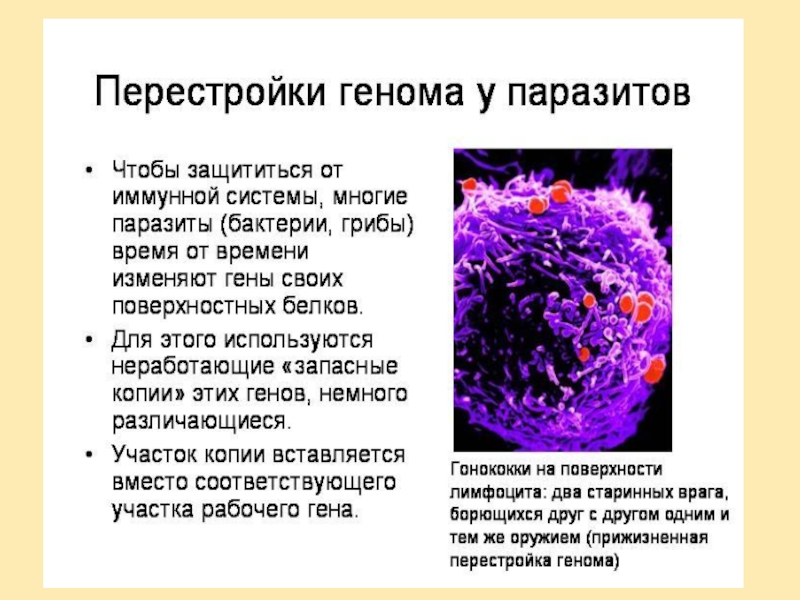
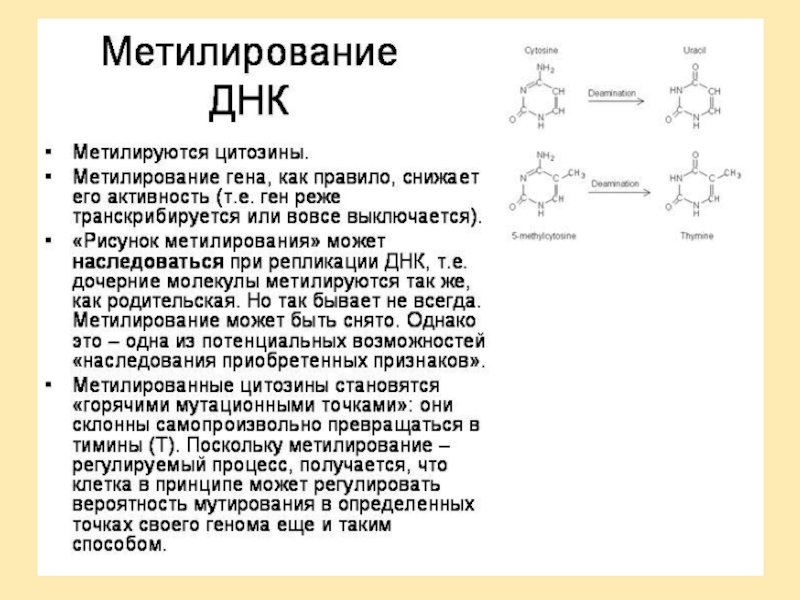
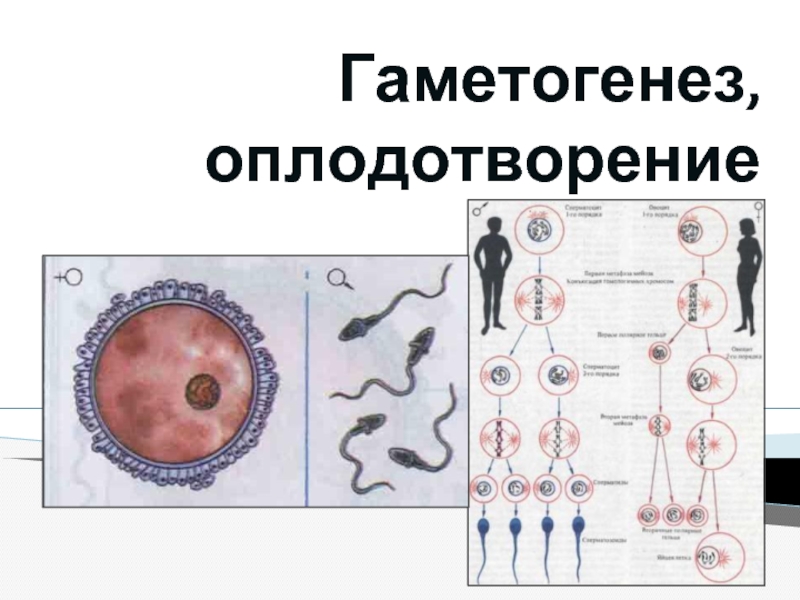
генов предрасположенности к тем или иным заболеваниям и так называемых генов "внешней среды". В частности, это гены, контролирующие детоксикацию чужеродных веществ. Все вещества, посутупающие в организм, метаболизируются в два этапа. На первом этапе образуются промежуточные генотоксические вещества. На втором этапе эти промежуточные метаболиты превращаются в растворимые безвредные соединения, которые выводятся из организма.
То, как организм реагирует на вредные воздействия среды, например, на табачный дым, также в значительной мере определяется активностью системы детоксикации. Например, у 3% женщин встречается сочетание генов, ответственных за обезвреживание канцерогенов табачного дыма ферментами печени, которое в 10 раз повышает риск развития рака молочной железы. Таким женщинам курить строго противопоказано. Влияют гены и на эффективность лечения различными препаратами. Так, лечение эндометриоза - заболевания, встречающегося почти у 10% женщин белой расы - широко используемым препаратом циклофероном у части больных безрезультатно по причинам генетического характера.
С частотой 15% встречается аллель гена аполипопротеина Е (белок, отвественный за связывание липидов), приводящий к ухудшению регенерации нервных тканей после травм или сотрясений мозга, особенно если индивид гомозиготен по данному аллелю. Значит, таким людям не стоит выбирать профессию боксера или автогонщика.










 мутацийТочечными называют мутации, которые изменяют небольшой участок ДНК, обычно длиной от одной")

 или вставке (инсерции) количества п.н.,")

, потеря пурина (такое")






")