біологічних наук,
старший викладач кафедри медико-біологічних основ фізичної культури
Скиба Ольга Олександрівна
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Фізіологія збудливих тканин презентация
Содержание
- 1. Фізіологія збудливих тканин
- 2. Література: Покровский В. М. Физиология человека. /
- 3. Фізіологія (от греч. physis — природа и
- 4. Напрямки фізіології
- 5. Методи фізіологічних досліджень 1. Експеримент Гострий
- 6. Об'єктивна графічна реєстрація біоелектричних потенціалів стала основою важливого розділу фізіології - електрофізіології.
- 9. УКРАЇНСЬКА ШКОЛА ФІЗІОЛОГІВ Чаговець В.Ю (1873-1941) Воронцов
- 10. Клетка– основная структурная и функциональная единица организма.
- 11. Основні властивості живих клітин Подразливість – здатність
- 12. Подразнення – вплив подразника на організм. Збудливість
- 13. Протилежний стан збудженню збудливих тканин називається гальмуванням.
- 14. Протилежний стан збудженню збудливих тканин називається гальмуванням.
- 15. Будова клітинної мембрани Жидкостно-мозаичная модель (Сингера и
- 16. Трехмерная жидкостно-мозаичная модель клеточной мембраны (по Сингеру—Николсону).
- 17. Основні функції клітинної мембрани Бар'єрна; Транспортна; Регуляторна; Контактна; Рецепторна; Секреторна.
- 18. Мембранний потенціал спокою (МПС) – це різниця
- 19. Умови виникнення МПС • 1. Наличие градиента
- 20. 1. Электрохимический градиент для ионов градиент концентрации
- 21. 2. Высокая избирательная проницаемость мембраны для ионов
- 22. 3. Наличие активного транспорта (Nа+,К+ насоса) в
- 23. Потенціал дії (ПД) ПД - швидке високоамплітудне
- 24. Тривалість ПД 0,3-10 мс
- 25. Фази ПД під час висхідної фази (фаза
- 26. Тривалість піка потенціалу дії у різних нервових
- 27. ПД виникає в той момент, коли деполяризація
- 28. Відмінності між ПД і ЛВ 1.
- 29. Під час розвитку ПД, виникає рефрактерність (незбудливість)
- 30. МАКСИМАЛЬНИЙ РИТМ ІМПУЛЬСАЦІЇ М. Є. Введенський перший
- 31. Механізм проведення збудження Проведення збуження вздовж нервових
- 32. Збільшення швидкості проведення збудження відбувається за рахунок
- 33. Закони проведення збудження Закон анатомічної і фізіологічної
- 34. Фізіологія синапсів
- 35. Синапс – (від грец. synapsis - дотик,
- 36. 1. Классификация синапсов по контакту (местонахождению)
- 38. 3. Классификация синапсов по способу передачи
- 39. Будова хімічного (нервово-м’язового) синапсу Пресинаптична частина
- 40. Механізм передачі через синапс 1. ПД по
- 41. Нейромедиаторы (нейротрансмиттеры) — биологически активные
- 42. Ацетилхолин – медиатор нервно-мышечного синапса Ацетилхолин выполняет
- 43. γ-Аминомасляная кислота (ГАМК, GABA) Аминокислота, важнейший
- 44. Особливості проведення інформації через хімічний синапс 1.
- 45. СПАСИБО ЗА ВНИМАНИЕ!
Слайд 1Фізіологія як наука: основні поняття, предмет та методи дослідження.
Фізіологія збудливих тканин
кандидат

Слайд 2Література:
Покровский В. М. Физиология человека. / В.М. Покровский, Г. Ф. Коротько.
- М.: Медицина, 1997; Т1- 448 с., Т2 - 368с.
Філімонов В.І. Фізіологія людини: Підручник. — К., 2010. — 776 с.
Кучеров І.С. Фізіологія людини і тварин. К.: Вища шк., 1991. 327 с.
Чайченко Г.М., Цибенко В.О., Сокур В.Д. Фізіологія людини і тварин. — К.: Вища шк., 2003.
Физиология человека: В 3-х т.: Пер. с англ. / Под ред. Р. Шмидта и Г. Тевса. — М.: Мир, 1996.
Солодков А. С. Физиология человека. Общая. Спортивная. Возрастная. – М. Терра-Спорт, Олимпия Пресс, 2005. – 520 с.
http://www.booksmed.com – медицинская библиотека.
http://vk.com/booksmed - группа VK
http://vk.com/mbofk - группа кафедры МБОФК
Філімонов В.І. Фізіологія людини: Підручник. — К., 2010. — 776 с.
Кучеров І.С. Фізіологія людини і тварин. К.: Вища шк., 1991. 327 с.
Чайченко Г.М., Цибенко В.О., Сокур В.Д. Фізіологія людини і тварин. — К.: Вища шк., 2003.
Физиология человека: В 3-х т.: Пер. с англ. / Под ред. Р. Шмидта и Г. Тевса. — М.: Мир, 1996.
Солодков А. С. Физиология человека. Общая. Спортивная. Возрастная. – М. Терра-Спорт, Олимпия Пресс, 2005. – 520 с.
http://www.booksmed.com – медицинская библиотека.
http://vk.com/booksmed - группа VK
http://vk.com/mbofk - группа кафедры МБОФК

Слайд 3Фізіологія (от греч. physis — природа и logos - учение) –
наука, що вивчає життєдіяльність (функціонування) організму та окремих його частин: клітин, тканин, органів, систем.
Предмет фізіології: функції живих організмів, взаємозв'язок між ними, регуляція та пристосування до зовнішнього середовища;
Фізіологічна функція (functio – деятельность) – специфічна діяльність системи чи органу, що має пристосувальне значення та спрямована на досягнення корисного (для організму) результату.
Предмет фізіології: функції живих організмів, взаємозв'язок між ними, регуляція та пристосування до зовнішнього середовища;
Фізіологічна функція (functio – деятельность) – специфічна діяльність системи чи органу, що має пристосувальне значення та спрямована на досягнення корисного (для організму) результату.
 – наука, що вивчає життєдіяльність")

Слайд 5Методи фізіологічних досліджень
1. Експеримент
Гострий (вівісекція);
В умовах ізольованого органу;
Хронічний (І.П. Павлов);
2.Графічна
реєстрація фізіологічних процесів
(графічна реєстрація АТ, дихальних актів, моторики ШКТ)
3. Метод реєстрації електричних потенціалів
(електрокардіограма, електроміограма)
(графічна реєстрація АТ, дихальних актів, моторики ШКТ)
3. Метод реєстрації електричних потенціалів
(електрокардіограма, електроміограма)
;В умовах ізольованого органу;Хронічний (І.П. Павлов);2.Графічна реєстрація фізіологічних процесів (графічна")
Слайд 6
Об'єктивна графічна реєстрація біоелектричних потенціалів стала основою важливого розділу фізіології -
електрофізіології.



Слайд 9УКРАЇНСЬКА ШКОЛА ФІЗІОЛОГІВ
Чаговець В.Ю (1873-1941)
Воронцов Д.С.
(1886-1965)
Костюк П. Г.
(1924-2010)
Щелков І. П.
(1833-1909)
Мечников
І. І.
(1845-1916)
(1845-1916)
Богомолець О.О.
(1881-1946)
Нагорний О.В.
(1887-1953)
Скок В.І.
(1932-2003)
Воронцов Д.С.(1886-1965)Костюк П. Г. (1924-2010)Щелков І. П.(1833-1909)Мечников І. І.(1845-1916)Богомолець О.О. (1881-1946)Нагорний")
Слайд 10Клетка– основная структурная и функциональная единица организма. Она способна к самообновлению,
саморегуляции и самовоспроизведению. Клетки объединяются в ткани.

Слайд 11Основні властивості живих клітин
Подразливість – здатність клітини активно реагувати на дію
факторів зовнішнього чи внутрішнього середовища (подразників) зміною обмінних процесів.
Подразники - це фактори зовнішнього або внутрішнього середовища, які впливають на живі структури і викликають активну зміну характеру їх життєдіяльності (подразнення).
Класифікація подразників:
За адекватністю (адекватні та неадекватні);
За природою подразнення (хімічні, фізичні, механічні, біологічні);
За силою подразнення (допорогові, порогові, надпорогові).
Подразники - це фактори зовнішнього або внутрішнього середовища, які впливають на живі структури і викликають активну зміну характеру їх життєдіяльності (подразнення).
Класифікація подразників:
За адекватністю (адекватні та неадекватні);
За природою подразнення (хімічні, фізичні, механічні, біологічні);
За силою подразнення (допорогові, порогові, надпорогові).

Слайд 12Подразнення – вплив подразника на організм.
Збудливість - це здатність збудливих тканин
реагувати на подразнення збудженням.
*Збудливість властива тільки нервовим, мязовим та секреторним тканинам!
Збудження – це активний фізіологічний процес, відповідь тканини на її подразнення, що проявляється у специфічній для неї діяльності (проведення збудження нервової тканиною, скорочення м'яза, секреція залози) і неспецифічних реакціях (генерація потенціалу дії, метаболічні зміни).
*Збудливість властива тільки нервовим, мязовим та секреторним тканинам!
Збудження – це активний фізіологічний процес, відповідь тканини на її подразнення, що проявляється у специфічній для неї діяльності (проведення збудження нервової тканиною, скорочення м'яза, секреція залози) і неспецифічних реакціях (генерація потенціалу дії, метаболічні зміни).
реакції збудливих тк-н у відповідь на подразнення:
нервової клітини - проведення нервового імпульсу,
м'язової клітини - скорочення,
секреторної - синтез і виділення біологічно активної речовини.

Слайд 13Протилежний стан збудженню збудливих тканин називається гальмуванням.
Гальмування - це активний
процес, що супроводжується специфічними змінами обміну речовин, що виражаються в уповільненні збудження.

Слайд 14Протилежний стан збудженню збудливих тканин називається гальмуванням.
Гальмування - це активний
процес, що супроводжується специфічними змінами обміну речовин, що виражаються в уповільненні збудження.

Слайд 15Будова клітинної мембрани
Жидкостно-мозаичная модель (Сингера и Николсона) – белки погружены в
фосфолипидный бислой.
Липидный бислой –
Фосфолипиды:
фосфатидилхолин (лецитин),
фосфатидилэтаноламин,
фосфатидилсерин,
фосфатидилинозит
Кардиолипин;
Сфингомиелин;
Холестерол;
Гликолипиды.
Белки -
Интегральные (каналы, переносчики, насосы, рецепторы)
Периферические (цитоскелет, гликокаликс)
*электрические явления, возникающие в возбудимых тканях, обусловлены электрическими свойствами клеточных мембран.
 – белки погружены в фосфолипидный бислой. Липидный бислой")
Слайд 16Трехмерная жидкостно-мозаичная модель клеточной мембраны (по Сингеру—Николсону). А — фосфолипидный бислой, в
который погружены белки; Б — различные моменты движения Na+ через клеточную мембрану.
. А — фосфолипидный бислой, в который погружены белки;")
Слайд 17Основні функції клітинної мембрани
Бар'єрна;
Транспортна;
Регуляторна;
Контактна;
Рецепторна;
Секреторна.

Слайд 18Мембранний потенціал спокою (МПС) – це різниця потенціалів між зовнішньою і
внутрішньою сторонами мембрани.
Суть теорії - мембранний потенціал спокою виникає завдяки направленого руху заряджених частинок.
В основному це дифузія іонів К+ через мембрану клітини з внутрішньоклітинного середовища в позаклітинне.
Суть теорії - мембранний потенціал спокою виникає завдяки направленого руху заряджених частинок.
В основному це дифузія іонів К+ через мембрану клітини з внутрішньоклітинного середовища в позаклітинне.
***Величина МПС становить від -50 до -90 мВ
Через фосфоліпідний бішар можуть проникати різні речовини, причому ступінь проникності (Р) – здатність клітинної мембрани пропускати ці речовини, залежить від різниці концентрацій цієї речовини по обидві сторони мембрани.
 – це різниця потенціалів між зовнішньою і внутрішньою сторонами мембрани. Суть")
Слайд 19Умови виникнення МПС
• 1. Наличие градиента концентраций для ионов K+, Na+,
Cl-;
• 2. Избирательно высокая проницаемость мембраны для К+;
• 3. Наличие активного транспорта (Nа+,К+ насоса) в мембране.
• 2. Избирательно высокая проницаемость мембраны для К+;
• 3. Наличие активного транспорта (Nа+,К+ насоса) в мембране.

Слайд 201. Электрохимический градиент для ионов
градиент концентрации ионов по обе стороны мембраны
(концентрация К+ внутри клетки больше, чем вне, а
ионов Na+ наоборот).
ионов Na+ наоборот).

Слайд 212. Высокая избирательная проницаемость
мембраны для ионов К+, Na+, Cl-
В состоянии покоя
мембрана проницательна для К+ и малопроницательна для Na+, Cl-.
(Относительная проницаемость 1:0,16:0,61)
Селективность каналов обусловлена тем, что каждый канал имеет:
• устье,
• селективный фильтр,
• воротной механизм (gate).
Проводимость одиночного открытого канала стабильна.
Суммарная проницаемость мембраны определяется соотношением открытых и закрытых каналов.
(Относительная проницаемость 1:0,16:0,61)
Селективность каналов обусловлена тем, что каждый канал имеет:
• устье,
• селективный фильтр,
• воротной механизм (gate).
Проводимость одиночного открытого канала стабильна.
Суммарная проницаемость мембраны определяется соотношением открытых и закрытых каналов.

Слайд 223. Наличие активного транспорта
(Nа+,К+ насоса) в мембране
Принцип работы. В клеточной мембране
имеется система переносчиков, каждый из которых связывается с тремя находящимися внутри клетки Na+ и выводит их наружу. С наружной стороны переносчик связывается с двумя находящимися
вне клетки К+ , которые переносятся в цитоплазму.
вне клетки К+ , которые переносятся в цитоплазму.
В следствии работы насоса: *
поддерживается высокая концентрация К+ внутри клетки, что обеспечивает постоянство величины потенциала покоя. Вследствие того что за
один цикл обмена ионов из клетки выводится на один положительный
ион больше, чем вводится, активный транспорт играет роль в создании
потенциала покоя.
* Поддерживается низкая концентрация Na+ внутри клетки, что обеспечивает работу механизма генерации потенциала.
 в мембранеПринцип работы. В клеточной мембране имеется система переносчиков,")
Слайд 23Потенціал дії (ПД)
ПД - швидке високоамплітудне коливання МПС, що виникає під
час збудження (від -90 до +30мВ);
В основі формування ПД лежать зміни іонної проникності мембрани (переважно рух Na+ в середину клітини).
Амплітуда і характер тимчасових змін потенціалу дії мало залежать від сили подразника, важливо лише, що б ця сила досягла критичної (порогової) величини, (закон «все або нічого», Х. Боудіч, 1871)
У природних умовах потенціали дії генеруються в нервових волокнах при подразненні рецепторів або збудженні нервових клітин.
В основі формування ПД лежать зміни іонної проникності мембрани (переважно рух Na+ в середину клітини).
Амплітуда і характер тимчасових змін потенціалу дії мало залежать від сили подразника, важливо лише, що б ця сила досягла критичної (порогової) величини, (закон «все або нічого», Х. Боудіч, 1871)
У природних умовах потенціали дії генеруються в нервових волокнах при подразненні рецепторів або збудженні нервових клітин.
ПД - швидке високоамплітудне коливання МПС, що виникає під час збудження (від -90")

Слайд 25Фази ПД
під час висхідної фази (фаза деполяризації) відбувається не просто зникнення
МПС, а виникає різниця потенціалів зворотного знака: внутрішній вміст клітини стає зарядженим позитивно по відношенню до зовнішнього середовища. Іони Nа+ заходять в клітину і заносять позитивний заряд. Відбувається реверсія мембранного потенціалу
(з -90 до +30 мВ)(овершут).
Під час низхідної фази (фази реполяризації) мембранний потенціал повертається до свого початкового значення. У момент досягнення вершини (піка) мембранний потенціал складає + 30 / + 40мВ. За піком ПД реєструється тривалі слідові зміни МП, після чого МП встановлюється на вихідному рівні.
(з -90 до +30 мВ)(овершут).
Під час низхідної фази (фази реполяризації) мембранний потенціал повертається до свого початкового значення. У момент досягнення вершини (піка) мембранний потенціал складає + 30 / + 40мВ. За піком ПД реєструється тривалі слідові зміни МП, після чого МП встановлюється на вихідному рівні.
 відбувається не просто зникнення МПС, а виникає різниця")
Слайд 26Тривалість піка потенціалу дії у різних нервових і скелетних м'язових волокон
варіює від 0,5 до 3 мс, причому фаза реполяризації триваліше фази деполяризації.
Зміни мембранного потенціалу, що відбуваються після піку ПД називають слідовими потенціалами. ( в клітинах відновлюється склад іонів завдяки роботі Na-K насосу.
Зміни мембранного потенціалу, що відбуваються після піку ПД називають слідовими потенціалами. ( в клітинах відновлюється склад іонів завдяки роботі Na-K насосу.

Слайд 27ПД виникає в той момент, коли деполяризація мембрани досягає критичного рівня
деполяризації.
Якщо сила подразника буде недостатньою, то ПД не виникне, а виникне локальна відповідь.
Якщо сила подразника буде недостатньою, то ПД не виникне, а виникне локальна відповідь.

Слайд 28Відмінності між ПД і ЛВ
1. ПД не залежить від сили
подразника, його амплітуда завжди постійна, а ЛВ – залежить;
2. ПД не здатен до сумації, а ЛВ може сумуватися (в результаті амплітуда деполяризації збільшується);
3. ПД підкоряється закону «все або нічого», а ЛВ виника на будь-який подразник.
4. ПД розповсюджується до місця призначення, а ЛВ затухає у місці виникнення.
2. ПД не здатен до сумації, а ЛВ може сумуватися (в результаті амплітуда деполяризації збільшується);
3. ПД підкоряється закону «все або нічого», а ЛВ виника на будь-який подразник.
4. ПД розповсюджується до місця призначення, а ЛВ затухає у місці виникнення.

Слайд 29Під час розвитку ПД, виникає рефрактерність (незбудливість) - зниження здатності клітини
відповідати на подразнення в результаті тимчасової інактивації натрієвих каналів.
Абсолютная рефрактерность – генерация ПД невозможна. период полной невозбудимости Обусловлена практически полной инактивацией натриевых каналов и повышением калиевой проводимости.
Относительная рефрактерность – генерация ПД возможна при увеличении интенсивности раздражителя, т.к. некоторая часть каналов для Na+ еще инактивирована
 - зниження здатності клітини відповідати на подразнення в")
Слайд 30МАКСИМАЛЬНИЙ РИТМ ІМПУЛЬСАЦІЇ
М. Є. Введенський перший звернув увагу на різну здатність
збудливих тканин відтворювати високі ритми подразнень.
Лабільність – здатність збудливої тканини реагувати на подразнення з визначеною швидкістю.
Максимальне число ПД («максимальний ритм»), яке здатна збудлива тканина генерувати за 1 с відповідно до ритму подразнення.
Лабільність визначається швидкістю процесів змін іонної провідності, що лежать в основі абсолютної і відносної рефрактерності.
Лабільність – здатність збудливої тканини реагувати на подразнення з визначеною швидкістю.
Максимальне число ПД («максимальний ритм»), яке здатна збудлива тканина генерувати за 1 с відповідно до ритму подразнення.
Лабільність визначається швидкістю процесів змін іонної провідності, що лежать в основі абсолютної і відносної рефрактерності.

Слайд 31Механізм проведення збудження
Проведення збуження вздовж нервових волокон здійснюється за допомогою місцевих
(локальних) струмів (Герман, 1899), що виникають між збудженою (деполяризованою) і нормально поляризованою ділянками волокна. Активна деполяризова ділянка (А) стає подразником для сусідньої (незбудженої) (Б) ділянки мембрани.
Завдяки такому естафетного механізму збудження поширюється вздовж усього волокна. В м'язових і немієлінових (безмякотних) нервових волокнах збудження здійснюється безперервно від однієї точки клітинної мембрани до іншої.
*У немієлінових волокнах швидкість поширення ПД залежить від: 1) діаметру волокна (чим менший діаметр, тим більший опір, тим менша швидкість ). Швидкість в тонких волокнах до 0,5 м/с.
По немієліновим волокнах збудження проводиться від нервових центрів до внутрішніх органів .
 струмів (Герман, 1899),")
Слайд 32Збільшення швидкості проведення збудження відбувається за рахунок мієлінізації волокон. Мієлінова оболонка
виконує роль ізолятора, який переривається через кожні 1-2 мм вузлами нервового волокна (перехвати Рваньє).
1. Збудження поширюється стрибкоподібно (сальтаторно), ПД виникає тільки у вузлах нервового волокна.
2. Збудження поширюється з великою швидкістю (до 120 м/с).
3. Збудження поширюється без декримента (без зниження сили збудження до кінця волокна.
*Декримент – ослаблення збудження у міру його розповсюдження вздовж нервового або м'язового волокна.
По мієлінових волокнах збудження поширюється від аналізаторів до ЦНС, до скелетних м'язів тобто там де потрібна висока швидкість реакції у відповідь.

Слайд 33Закони проведення збудження
Закон анатомічної і фізіологічної цілісності волокна - збудження може
поширюватися по нервовому волокну тільки в разі його морфологічної та функціональної цілісності.
• Закон двостороннього проведення збудження – збудження, що виникає в одній ділянці нерва, поширюється в обидві сторони від місця свого виникнення.
В ЦНС збудження завжди поширюється по аксону через синаптичну щілину до наступної клітини (ортодромнно).
• Закон ізольованого проведення - збудження, що поширюється по волокну, що входить до складу нерва, не передається на сусідні нервові волокна.
• Закон двостороннього проведення збудження – збудження, що виникає в одній ділянці нерва, поширюється в обидві сторони від місця свого виникнення.
В ЦНС збудження завжди поширюється по аксону через синаптичну щілину до наступної клітини (ортодромнно).
• Закон ізольованого проведення - збудження, що поширюється по волокну, що входить до складу нерва, не передається на сусідні нервові волокна.


Слайд 35Синапс – (від грец. synapsis - дотик, з'єднання) місце спеціалізованого контакту
між нервовими клітинами або між нервовими і ефекторними клітинами, через який відбувається передача нервових імпульсів.
Уперше термін «синапс» запропонував Шерінгтон Чарльз Скотт (1857-1952)
Нейрон може мати від 1 тис. до 10 тис. синапсів
 місце спеціалізованого контакту між нервовими клітинами або")
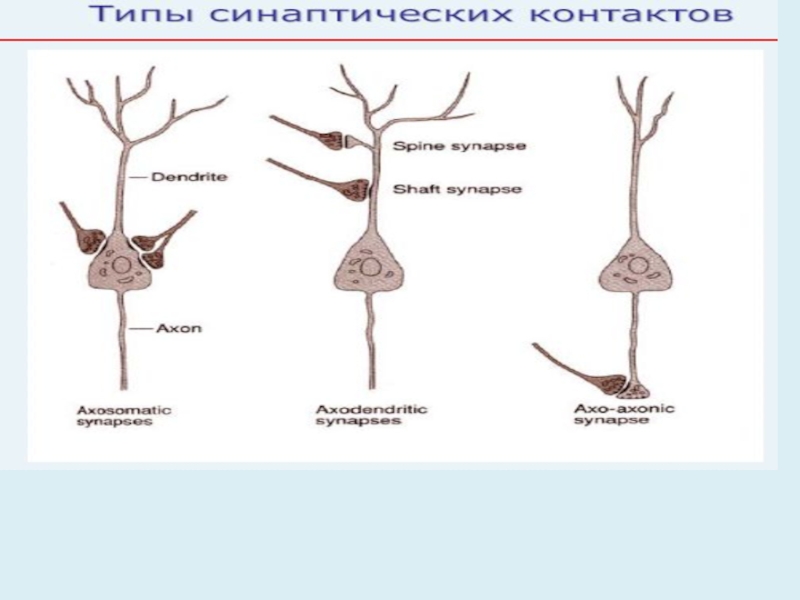
Слайд 361. Классификация синапсов по контакту (местонахождению)
Нейрон-нейрон
нейрон-эффектор (руховий нерв закінчується на волокні скелетного м'яза)
-аксон-дендрит -нейрон-мышца (нервно-мышечный синапс)
-аксон-сома -нейрон-железа
-аксон-аксон
-дендрит-дендрит
-дендрит-сома
(І. нейро-нейрональні синапси) (ІІ. нервово-м̛язові синапси)
*Збудження і гальмування передаються з однієї нервової клітини на іншу.
2. Функциональная классификация синапсов
Возбуждающие Тормозные
-аксон-дендрит -нейрон-мышца (нервно-мышечный синапс)
-аксон-сома -нейрон-железа
-аксон-аксон
-дендрит-дендрит
-дендрит-сома
(І. нейро-нейрональні синапси) (ІІ. нервово-м̛язові синапси)
*Збудження і гальмування передаються з однієї нервової клітини на іншу.
2. Функциональная классификация синапсов
Возбуждающие Тормозные
 Нейрон-нейрон нейрон-эффектор (руховий нерв закінчується на")
Слайд 383. Классификация синапсов по способу передачи
химические
электрические смешанные
Их большинство простой переход передача и
Передача возбуждения заряда с одной через химич.
происходит за счет мембраны на вещество и
выделения медиатора др. мембрану через электрич.
сигнал передается
электрическим током
Их большинство простой переход передача и
Передача возбуждения заряда с одной через химич.
происходит за счет мембраны на вещество и
выделения медиатора др. мембрану через электрич.
сигнал передается
электрическим током
Химические синапсы классифицируются по характеру медиатора:
Если используется ацетилхолил- хомоэргический синапс
Адреналин или норадренали- адренэргические
Дофамин-дофминэргические
Серотонин-серотонинэргические
Белки- пептидоэргические и др.

Слайд 39Будова хімічного (нервово-м’язового) синапсу
Пресинаптична частина
Синаптична щілина
Постсинаптична частина
 синапсуПресинаптична частинаСинаптична щілинаПостсинаптична частина")
Слайд 40Механізм передачі через синапс
1. ПД по аксону надходить до пресинаптичної мембрани
і викликає її деполяризацію (зменшення МПС).
2. Деполяризація спричиняє відкриття в пресинаптичній мембрані Са+ каналів, через які в середину нервового закінчення входять іони Са+ і запускають виділення медіатора.
3. Під впливом іонів Са+, лопаються пухирці з медіатором (ацетилхолін) , який надходить до синаптичної щілини.
4. Медіатор з'єднується з білком-рецептором на постсинаптичній частині.
5. Відкриваються хемозбудливі канали, через які рухаються іони (Na+ у середину, K+ - назовні). Виникає потенціал кінцевої пластинки (збудження постсинаптичної мембрани).
2. Деполяризація спричиняє відкриття в пресинаптичній мембрані Са+ каналів, через які в середину нервового закінчення входять іони Са+ і запускають виділення медіатора.
3. Під впливом іонів Са+, лопаються пухирці з медіатором (ацетилхолін) , який надходить до синаптичної щілини.
4. Медіатор з'єднується з білком-рецептором на постсинаптичній частині.
5. Відкриваються хемозбудливі канали, через які рухаються іони (Na+ у середину, K+ - назовні). Виникає потенціал кінцевої пластинки (збудження постсинаптичної мембрани).

Слайд 41Нейромедиаторы (нейротрансмиттеры)
— биологически активные химические вещества, посредством которых осуществляется передача
электрического импульса с нервной клетки через синаптическое пространство. Нейромедиаторы характеризуются способностью реагировать со специфическими белковыми рецепторами клеточной мембраны , инициируя цепь биохимических реакций, вызывающих изменение трансмембранного тока ионов, что приводит к деполяризации мембраны и возникновению потенциала действия.
 — биологически активные химические вещества, посредством которых осуществляется передача электрического импульса с нервной")
Слайд 42Ацетилхолин – медиатор нервно-мышечного синапса
Ацетилхолин выполняет функцию медиатора в синапсах периферической
и ЦНС:
Ах выделяется из аксонов двигательных нейронов соматической н.с. в нервно-мышечных синапсах, где действует на никотиновые холинорецепторы.
Ах выделяется из аксонов преганглионарных нейронов вегетативной нервной системы в синапсах, находящихся в ганглиях за пределами ЦНС, где действуют на никотиновые холинорецепторы.
Ах выделяется из аксонов двигательных нейронов соматической н.с. в нервно-мышечных синапсах, где действует на никотиновые холинорецепторы.
Ах выделяется из аксонов преганглионарных нейронов вегетативной нервной системы в синапсах, находящихся в ганглиях за пределами ЦНС, где действуют на никотиновые холинорецепторы.

Слайд 43γ-Аминомасляная кислота (ГАМК, GABA)
Аминокислота, важнейший тормозной нейромедиатор центральной нервной системы
человека и млекопитающих.
γ-Аминомасляная кислота является биогенным веществом. Содержится в ЦНС и принимает участие в нейромедиаторных и метаболических процессах в мозге.
γ-Аминомасляная кислота является биогенным веществом. Содержится в ЦНС и принимает участие в нейромедиаторных и метаболических процессах в мозге.
 Аминокислота, важнейший тормозной нейромедиатор центральной нервной системы человека и млекопитающих. γ-Аминомасляная")
Слайд 44Особливості проведення інформації через хімічний синапс
1. Однобічність проведення.
2.Затримка проведення (Час від
надходження нервового імпульсу до розвитку постсинаптичної відповіді в хімічному синапсі становить близько 0,2 - 0,5 мс, причому основна частина цього часу витрачається на виділення медіатора).
3. Сумація.
3. Сумація.







