- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Funcţiile biologice ale lipidelor презентация
Содержание
- 1. Funcţiile biologice ale lipidelor
- 2. OBIECTIVELE: Funcţiile biologice ale lipidelor. Clasificarea lipidelor(
- 3. LIPIDE Substanţe organice, insolubile în
- 4. ROLUL Energetic - principala formă de
- 5. CLASIFICAREA LIPIDELOR 1. DUPĂ STRUCTURA CHIMICĂ (HIDROLIZĂ)
- 6. CLASIFICAREA LIPIDELOR DUPĂ STRUCTURA CHIMICĂ II. Lipidele
- 7. CLASIFICAREA LIPIDELOR DUPĂ STRUCTURA CHIMICĂ Complexe
- 8. CLASIFICAREA LIPIDELOR DUPĂ STRUCTURA CHIMICĂ
- 9. CLASIFICAREA LIPIDELOR DUPĂ STRUCTURA CHIMICĂ III.
- 10. CLASIFICAREA LIPIDELOR 2. DUPĂ PROPRIETĂŢILE FIZICO-CHIMICE: Lipide
- 11. LIPID NEPOLAR LIPID POLAR
- 12. CLASIFICAREA LIPIDELOR 3. DUPĂ ROLUL BIOLOGIC: 1.
- 13. DISTRIBUȚIA LIPIDELOR Lipidele constituie 10-20% din masa
- 14. STRUCTURA CHMICĂ A LIPIDELOR
- 15. AG – STRUCTURA, PROPRIETĂŢI AG- derivaţii hidrocarburilor
- 16. STRUCTURA CHIMICĂ A AG SATURAȚI CH3-(CH2)2-COOH
- 17. STRUCTURA CHIMICĂ A AG MONONESATURAŢI
- 18. AG POLINESATURAŢI a.linoleic (C18:2cis Δ 9,12
- 19. PROPRIETĂŢILE AG AG saturaţi până la C10
- 20. PROPRIETĂŢILE CHIMICE AG saturaţi şi nesaturaţi pot
- 21. ROLUL AG: Sursă de energie pentru miocard
- 22. GLICERIDE ( ACILGLICEROLII, GRĂSIMI NEUTRE)
- 23. GLICERIDE - MONOGLICERIDE- MAG
- 24. DIGLICERIDE - DAG
- 25. TRIGLICERIDE - TAG
- 26. ROLUL TAG Constituie grăsimea de rezervă din
- 27. PROPRIETĂŢILE TAG Sunt determinate de natura şi
- 28. GLICEROFOSFOLIPIDELE (FOSDOGLICERIDE) Derivaţi ai acidului fosfatidic Reprezentanţii:
- 29. FOSFATIDILSERINA
- 30. FOSFATIDILETANOLAMINA (CEFALINELE)
- 31. FOSFATIDILCOLINA (LECITINA)
- 32. FOSFATIDILINOZITOLUL
- 33. FOSFATIDILINOZITOLUL
- 34. PLASMALOGENI creier muşchi nervi
- 35. CARDIOLIPINA (DIFOSFATIDILGLICEROL)
- 36. ROLUL FOSFOGLICERIDELOR Rolul: Structural Fosfatidilcolina cu 2
- 37. PROPRIETĂŢILE FL Caracter amfipatic (lecitinile şi cefalinele
- 38. PROPRIETĂŢILE FL 2. Proprietăţi tensioactive puternice 3.
- 39. SFINGOLIPIDELE Derivaţi ai sfingozinei Sunt abundente în
- 40. SFINGOLIPIDELE Proprietăţi; amfioni, au caracter amfipatic 2 categorii: sfingomielina şi glicosfingolipidele
- 41. SFINGOZINA Sfingozina +AG= ceramidă
- 42. SFINGOMIELINA: CERAMIDA+ REST DE FOSFORILCOLINĂ
- 43. SFINGOMIELINA Sfingozina +AG = ceramidă Ceramida+ rest de fosforilcolină = sfingomielina
- 44. SFINGOMIELINA Sfingomielina: constituent al fibrelor nervoase mielinice
- 45. GLICOLIPIDE (GLICOSFINGOLIPIDELE) - includ ceramida
- 46. GLICOLIPIDE (GLICOSFINGOLIPIDELE) Cerebrozide – se află
- 47. SULFATIDE:
- 48. 3. Gangliozide: sunt prezente în
- 49. GANGLIOZIDE:
- 50. STEROIZII - STEROLII Sunt derivaţi ai ciclului pentanoperhidrofenantren (steranului –C17)
- 51. COLESTEROLUL Sterol C27 Origine animală (lipseşte în
- 52. STRUCTURA CHIMICĂ A COLESTEROLULUI LIBER
- 53. STRUCTURA CHIMICĂ A COL ESTERIFICAT
- 54. ACIZII BILIARI Se sintetizează în ficat
- 55. Acidul colanic Structura chimică a AB Acid
- 56. ACIZII BILIARI La PH alcalin se găsesc
- 58. STRUCTURA CHIMICĂ A AB
- 59. ACIZII BILIARI Bacteriile intestinale: 1. pot înlătura
- 60. ACIZII BILIARI Rolul: proprietăţi de solubilitate
- 61. MEMBRANELE BIOLOGICE Structuri superorganizate constituienţii de
- 62. FUNCŢIILE BIOLOGICE ALE MEMBRANELOR: de separare
- 63. FUNCŢIILE BIOLOGICE ALE MEMBRANELOR: de transformare
- 64. STRUCTURA MB Componenţi de bază:
- 65. Fosfolipidele
- 67. MEMBRANELE CELULELOR EUCARIOTE CONŢIN COLESTEROL colesterol
- 69. ROLUL LIPIDELOR STRUCTURALE: Fosfolipidele şi glicolipidele –
- 70. PROTEINELE MEMBRANARE După localizare: - periferice
- 71. PROTEINELE MEMBRANARE Proteinele periferice sunt legate de
- 72. ROLUL PROTEINELOR MEMBRANARE asigură transportul, participă în transmiterea informaţiei, rol în transformările de energie.
- 73. GLUCIDELE MEMBRANARE Sunt sub formă de
- 74. Lanţul de glucide ataşat pe lipide (glicolipide)
- 76. Modelul mozaic- fluid: Bistratul de fosfolipide
- 77. PROPRIETĂŢILE MEMBRANELOR BIOLOGICE. Fluiditatea - starea în
- 78. PROPRIETĂŢILE MEMBRANELOR BIOLOGICE. Asimetria: 1. repartizarea neuniformă
- 79. DISTRIBUŢIA LIPIDELOR ESTE ASIMETRICĂ exemple de
- 80. PROPRIETĂŢILE MEMBRANELOR BIOLOGICE. Mobilitatea Moleculele lipidelor
- 81. Proprietăţile membranelor biologice Permeabilitatea selectivă Dublu
- 82. se realizează: Prin transport pasiv (independent de
- 83. Transportul pasiv se realizează prin: Difuzie simplă Difuzie facilitată Osmoză
- 84. Difuzia simplă
- 85. O substanţă difundează conform gradientului său de
- 86. Viteza de difuzie se modifică la: Creşterea
- 87. Se realizeaza: prin dublul strat lipidic: pentru
- 88. Difuzia facilitată se face prin intermediul unei proteine membranare.
- 89. a) transportul anionilor; b) transportul ureei; c) transportul glicerolului; d) transportul
- 90. Osmoza
- 91. Apa se va deplasa din partea hipotonică (diluată) spre cea hipertonică (concentrată)
- 92. Osmoza este apa care se deplasează conform gradientului său de concentraţie
- 93. Transport activ Are nevoie de SURSĂ DE
- 94. Transport activ – in sens contrar
- 95. pompa Na+/K+ (Na+/K+-ATP-aza) - transportă 3Na+ spre exterior si
- 96. 3. pompa de Ca2+ (Ca 2+-ATP-aza) - este caracteristică
- 97. 4. pompele protonice (H+-ATP-azele) se întâlnesc:
- 98. Se realizeaza cu ajutorul proteinelor transportor
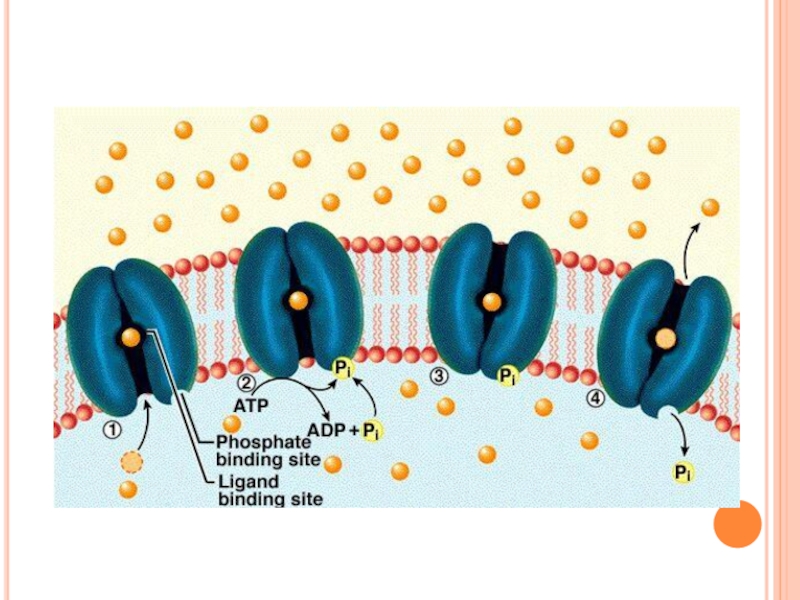
- 99. Transport activ secundar
- 100. Endocitoza - transportul intracelular al moleculeor mari,
- 102. Proteinele membranare permit pasajul pentru substanţele oprite de lipide:
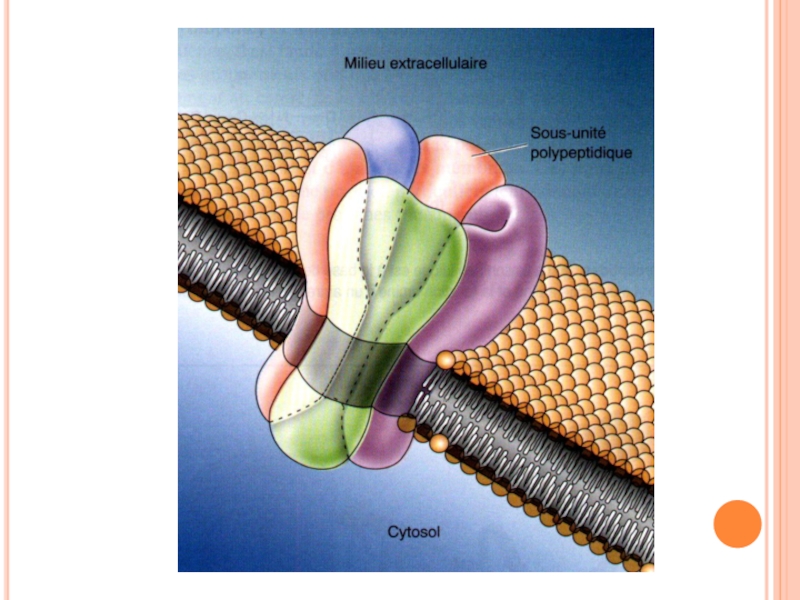
- 103. Canalele sunt foarte specifice: o singură substanţă
- 104. Canalele membranare sunt deseori formate din mai multe subunităţi:
- 106. Unele canale se pot deschide şi închide:
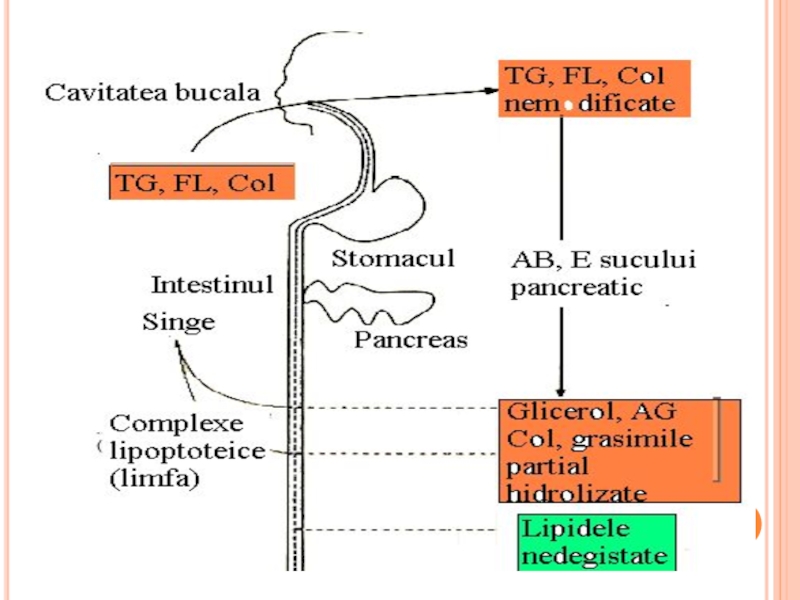
- 107. Digestia şi absorbţia lipidelor
- 108. IMPORTANŢA LIPIDELOR ÎN ALIMENTAŢIE. Aportul
- 109. DIGESTIA LIPIDELOR ALIMENTARE Digestia lipidelor începe în
- 110. DIGESTIA LIPIDELOR ALIMENTARE La nou născut, ambele
- 111. DIGESTIA LIPIDELOR ÎN INTESTIN pH alcalin -
- 112. DIGESTIA LIPIDELOR ÎN TGI În intestin -
- 113. DIGESTIA TAG lipazei pancreatice: Este o glicoproteidă,
- 114. DIGESTIA FOSFOGLICERIDELOR fosfolipazele (A1, A2, C,
- 115. DIGESTIA FOSFOGLICERIDELOR Cea mai activă este fosfolipaza
- 116. DIGESTIA COL Colesterolesteraza pancreatică (activată de AB) scindează Col esterificat în Col şi AG
- 118. PRODUSELE FINALE ALE DIGESTIEI LIPIDELOR: Acizii graşi Monogliceridele Colesterolul Lizolecitina
- 119. REGLAREA DIGESTIEI Colecistochinina (CCK) determină contracţia vezicii
- 120. REGLAREA DIGESTIEI Secretina - stimulează eliberarea de
- 121. ABSORBŢIA LIPIDELOR La nivelul porţiunii superioare a
- 122. CICLUL ENTERO-HEPATIC AG +AB formează o soluţie
- 123. MALABSORBȚIA LIPIDELOR Malabsorbţia lipidelor caracterizată prin creşterea
- 124. RESINTEZA LIPIDELOR ÎN ENTEROCITE Rolul: sinteza lipidelor
- 125. RESINTEZA LIPIDELOR ÎN ENTEROCITE MAG
- 126. RESINTEZA LIPIDELOR Moleculele lipidelor reconstituite împreună cu
- 127. METABOLISMUL ACIZILOR GRAȘI ȘI TAG
- 128. OBIECTIVELE: Biosinteza acizilor graşi: saturaţi cu număr
- 129. BIOSINTEZA AG Sinteza AG şi încorporarea
- 130. BIOSINTEZA AG CUPRINDE MAI MULTE PROCESE:
- 131. Sinteza de novo cu formarea acidului palmitic
- 132. PARTICULARITĂŢILE SINTEZEI AG Are loc
- 133. ACID GRAS SINTAZA = Palmitat sintetaza
- 134. ACID GRAS SINTAZA
- 135. ACID GRAS SINTAZA Fiecare monomer cuprinde 2
- 136. Lehn fig 21-5 modified
- 137. ACID GRAS SINTAZA (AGS)
- 138. SINTEZA DE NOVO CU FORMAREA ACIDULUI PALMITIC
- 139. I. TRANSFERUL LUI ACETIL COA DIN MITOCONDRII ÎN CITOZOL
- 140. II. SINTEZA DE MALONIL COA
- 141. Activatori -citrat, insulina Inhibitori - palmitoil CoA, glucagonul
- 142. III. SINTEZA ACIDULUI PALMITIC
- 143. III: SINTEZA ACIDULUI PALMITIC
- 144. III: SINTEZA ACIDULUI PALMITIC
- 145. III: SINTEZA ACIDULUI PALMITIC
- 146. REACȚIA SUMARĂ:
- 147. Biosinteza acizilor graşi saturaţi cu număr
- 148. ELONGAREA AG Localizată: reticulul endoplasmatic
- 149. ELONGAREA AG
- 150. BIOSINTEZA ACIZILOR GRAŞI NESATURAŢI Se produce în
- 151. BIOSINTEZA AG NESATURAŢI Sistemul de desaturare are
- 152. BIOSINTEZA AG MONOENICI
- 153. BIOSINTEZA AG POLIENICI Acidul linoleic şi linolenic
- 154. BIOSINTEZA A . ARAHIDONIC
- 155. SINTEZA TAG 2 căi: calea monoacilglicerolului:
- 156. 1. CALEA MONOACILGLICEROLULUI
- 157. II. CALEA GLICEROLFOSFATULUI
- 158. ORIGINEA GLICEROL FOSFATULUI În ficat: În ţesut adipos
- 159. Oxidarea AG Oxidarea glicerolului Sinteza corpilor cetonici
- 160. METABOLISMUL TAG Are loc preponderent în ficat,
- 161. MOBILIZAREA TAG DIN ŢESUTUL ADIPOS are loc
- 163. TRIGLICERIDLIPAZA cunoscută ca “lipaza hormonsensibilă”. Enzima este
- 164. SOARTA AG ŞI GLICEROLULUI: AG sunt transportaţi
- 165. SOARTA GLICEROLULUI: Glicerolul nu poate fi folosit
- 166. OXIDAREA GLICEROLULUI – ÎN FICAT localizat: citoplasma E1 glicerolkinaza E2 glicerolfosfatDH
- 167. OXIDAREA AG SATURAŢI CU NUMĂR PAR DE
- 168. OXIDAREA AG SATURAŢI CU NUMĂR PAR DE
- 169. ΒETA OXIDAREA AG 3 etape: Activarea AG
- 170. ACTIVAREA AG: R-COOH + HS-CoA
- 171. ACTIVAREA AG: R-COOH + ATP ?
- 173. TRANSFERUL LUI ACIL COA ÎN MITOCONDRII Acil
- 174. TRANSFERUL LUI ACIL COA ÎN MITOCONDRII
- 175. Carnitine Acyl Transferase I and II Lehn fig 17-6
- 177. REGLAREA Malonil CoA înhibă ACT I---deci palmitatul
- 178. DEFICIENŢA DE CARNITINĂ 1. congenitale –
- 179. BETA OXIDAREA PROPRIU ZISĂ
- 182. BILANŢUL ENERGETIC Stoichiometria unui ciclu de oxidare:
- 183. STOICHIOMETRIA OXIDĂRII A. PALMITIC 16/2 -1 =
- 184. BILANŢUL ENERGETIC AL OXIDĂRII COMPLETE A ACIDULUI
- 185. OXIDAREA AG NESATURAŢI β-oxidarea AG nesaturaţi se
- 187. Pentru AG polienici e
- 189. OXIDAREA AG CU NUMĂR IMPAR DE ATOMI
- 190. OXIDAREA AG CU NUMĂR IMPAR DE ATOMI
- 191. BILANȚUL ENERGETIC PENTRU AG CU NUMĂR IMPAR
- 192. OXIDAREA AG ÎN PEROXISOMI Caracteristic AG C20-C26
- 193. Numărul peroxisomilor creşte în diabet, inaniţie, la
- 194. ALTE TIPURI DE OXIDĂRI α-oxidare – la
- 195. Comparaţia sinteză/degradare AG cu număr par de atomi de carbon
- 196. REGLAREA METABOLISMULUI LIPIDIC Se efectuiază în mai
- 197. Factorii hormonali: Catecolaminele, Glucagonul stimulează
- 198. INSULINA activează biosinteza AG în ficat Activează
- 199. METABOLISMUL LIPIDIC ÎN PERIOADELE POSTPRANDIALE Mărit raportul
- 200. METABOLISMUL LIPIDIC ÎN PERIOADELE POSTPRANDIALE
- 201. METABOLISMUL LIPIDIC ÎN PERIOADELE INTERALIMENTARE Scăzut
- 202. SINTEZA CORPILOR CETONICI (CETOGENEZA) acetoacetatul, β-hidroxibutiratul
- 203. SINTEZA CORPILOR CETONICI (CETOGENEZA) Are loc când
- 205. CETOGENEZA
- 206. UTILIZAREA CORPILOR CETONICI Sunt substrate energetice preferate
- 207. ACTIVAREA ACETOACETATULUI Transportul corpilor cetonici prin sânge
- 208. ACTIVAREA ACETOACETATULUI: 1. Calea tioforazică: Reacţie de schimb cu succinil CoA
- 209. ACTIVAREA ACETOACETATULUI 2. Calea tiokinazică: -
- 210. ACTIVAREA BETA-HIDROXIBUTIRATULUI Cea mai importantă cale este
- 211. UTILIZAREA CORPILOR CETONICI
- 212. Prin oxidarea unui mol de acetoacetat se
- 214. CETONEMIE, CETONURIE Cetonemie- mărirea c% de
- 215. CAUZELE CETONEMIEI Mărirea concentrației lui Acetil -
- 216. CETOACIDOZA DIABETICĂ CC depăşesc 20 mmol/l
- 217. CETOACIDOZA DIABETICĂ - Sub influienţa
- 218. CETOGENEZA ÎN DIABET Raportul glucagon/insulină creşte; Creşte
- 219. CETOACIDOZA DIABETICĂ Eliminarea hidroxibutiratului şi acetoacetatului
- 220. CETOGENEZA ÎN INANIŢIE Glicemia este scăzută, corpii
- 221. METABOLISMUL LIPIDELOR STRUCTURALE. EICOSANOIZII
- 222. OBIECTIVELE Metabolismul colesterolului. Biosinteza colesterolului – substratele,
- 223. SINTEZA COLESTEROLULUI Se sintetizează din Acetil-CoA Necesită
- 224. SINTEZA COLESTEROLULUI Are loc în 3 etape:
- 226. СИНТЕЗ ХОЛЕСТЕРОЛА
- 227. REGLAREA HMG-COA REDUCTAZA la nivelul
- 228. REGLAREA HMG-COA REDUCTAZA la nivelul
- 229. REGLAREA Reacţia – limită a
- 230. REGLAREA HMG-COA REDUCTAZA la nivelul
- 231. Conţinutul Col depinde de regimul alimentar.
- 232. CĂILE DE TRANSFORMARE ALE COLESTEROLULUI 1. În
- 233. CATABOLISMUL ŞI ELIMINAREA COL 1. Calea biliară:
- 234. LITIAZA BILIARĂ Colesterolul (care este hidrofob) este
- 235. SINTEZA GLICEROFOSFOLIPIDELOR LOCALIZAREA: Are loc intensiv în
- 236. SINTEZA FOSFATIDILCOLINEI (PC) ȘI FOSFATIDILETANOLAMINEI(PE) Colina
- 237. 1. SINTEZA PC
- 238. IMPORTANȚA REUTILIZĂRII COLINEI COLINA în organism: Sintetizată
- 239. (etanolamină) (Fosfoetanolamină) (CDP-etanolamină) (Fosfatidiletanolamină)
- 240. SINTEZA FOSFATIDILSERINEI (PS) FOSFATIDILETANOLAMINA
- 241. SINTEZA PC ÎN FICAT din PS
- 242. Ficatul este un organ specific de
- 243. The synthesis of PE, PC, PS in eukaryotic cells.
- 244. SINTEZA PI ȘI CARDIOLIPINEI DIN A. FOSFATIDIC
- 245. 4 5
- 246. CATABOLISMUL FOSFOGLICERIDELOR ARE LOC SUB ACŢIUNEA FOSFOLIPAZELOR
- 247. FOSFOLIPAZELE A1 – este prezentă în multe
- 248. SINTEZA SFINGOLIPIDELOR Se formează din palmitoil CoA
- 250. CATABOLISMUL SFINGOMIELINELOR Sfingomielinaza (lizozomală): ceramid+fosforilcolină Ceramidaza – degradează legătura N acilică
- 251. CATABOLISMUL SFINGOMIELINELOR Ceramida poate fi implicată în răspunsul la stres Sfingozina inhibă proteinkinaza C
- 252. SINTEZA GLICOLIPIDELOR Sinteza cerebrozidelor: La sinteză
- 253. BIOSINTEZA CEREBROZIDELOR
- 254. GRUPARI SULFAT: DE LA TRASNPORTORUL DE SULFAT
- 255. SINTEZA GANGLIOZIDELOR La sinteză participă UDP-N-acetil galactozamină Globozid +CMP_NANA-----Gangliozid +CMP
- 256. NANA is synthesized from N-acetyl-mannosamine-6-P and PEP Examples of ganglioside structure
- 257. CATABOLISMUL GLICOLIPIDELOR Cerebrozidele sunt degradate de hexozidaze
- 258. LIPIDOZE TISULARE Boli ereditare, caracterizate prin acumularea
- 259. LIPIDOZE TISULARE 1. Neimann-Pick - deficienţa de
- 260. LIPIDOZE TISULARE 2. afecţiunea Tay-Sachs – lipsa
- 261. EICOSANOIZII Sunt substanţe biologic active de natură
- 262. EICOSANOIZII Deosebim: Prostaglandinele (clasice- PG şi endoperoxizii
- 263. STRUCTURA CHIMICĂ- PROSTAGLANDINE Reprezintă AG nesaturaţi
- 265. TX conţin în structura lor ciclul
- 266. STRUCTURA CHIMICĂ - LT LT (LTA-LTE) –
- 267. BIOSINTEZA EICOSANOIZILOR Are loc la nivelul
- 268. I: SINTEZA ACIDULUI ARAHIDONIC Derivat al acidului
- 270. II.SINTEZA PG, TX ŞI LT 2 căi:
- 272. CICLOOXIGENAZA este o hemoproteină, cu o activitate
- 275. COX-1 – enzimă constitutivă; mucoasa gastrică, plachete,
- 276. INHIBITORII SINTEZEI: Corticosteroizii – inhibă fosfolipaza A2
- 277. CATABOLISMUL PG Viaţă biologică scurtă: TXA2
- 278. MECANISMELE DE ACŢIUNE Au acţiune autocrină sau
- 279. ACŢIUNILE BIOLOGICE Diferă nu numai de natura
- 280. Plachetele sanguine (trombocite) conţin tromboxan sintază;
- 281. ACŢIUNILE BIOLOGICE La nivelul TGI: PGF2α
- 282. ACŢIUNILE BIOLOGICE Sistemul respirator: PGE şi
- 283. ACŢIUNILE BIOLOGICE SNC: reglarea transmisiei sinaptice.
- 284. ACŢIUNILE BIOLOGICE Sistemul de reproducere: la
- 285. ACŢIUNILE BIOLOGICE Sistem circulator: Prostaciclina şi
- 286. ACŢIUNILE BIOLOGICE LT sunt sintetizate de leucocite
- 287. ACŢIUNILE BIOLOGICE LT (LTB4) –determină acumularea de
- 288. METABOLISMUL LIPOPROTEINELOR VITAMINELE LIPOSOLUBILE
- 289. OBIECTIVELE Metabolismul lipoproteinelor Cauze, mecanismele dereglării metabolismului
- 290. Metabolismul lipoproteinelor
- 291. LIPOPROTEINELE (LP) LP- sînt complexe
- 292. LIPOPROTEINELE (LP) Rolul LP Transportul lipidelor exogene
- 293. STRUCTURA LP miez hidrofob - lipidele nepolare
- 294. METODELE DE SEPARARE Ultracentrifugare Electroforeză.
- 295. LIPOPROTEINELE
- 299. ENZIME IMPLICATE ÎN METABOLISMUL LIPOPROTEINELOR Lipoproteinlipaza endotelială
- 300. METABOLISMUL CHILOMICRONILOR Sunt sintetizaţi în celulele mucoasei
- 301. METABOLISMUL CHILOMICRONILOR După sinteză sunt secretaţi
- 303. CATABOLISMUL CHILOMICRONILOR În prima etapă a.
- 306. METABOLISMUL CHILOMICRONILOR
- 307. PATOLOGIA Deficitul înăscut de LPL- hiper-CM-emie Mărirea
- 308. PRE-β-LP (VLDL) sintetizate în ficat din lipidele
- 309. CATABOLISMUL VLDL La nivelul ţesuturilor extrahepatice sub
- 312. PATOLOGIA Steatoza (infiltraţia grasă a ficatului) –
- 313. LDL - β-LP se formează în plasmă
- 315. RECEPTORII LDL 1) Receptori LDL clasici- sunt
- 316. CATABOLISMUL LDL I: În ficat, suprarenale, limfocite,
- 317. K Colesterolul liber: inhibă HMG-CoA-reductaza, ca
- 318. II. ÎN MACROFAGE: Macrofagele încorporează LDL care
- 320. PATOLOGIA Hiperlipoproteinemia de tip II – deficienţă
- 321. α-LP - HDL conţinut lipidic de 45-55%,
- 322. METABOLISMUL HDL HDL captează Col liber atît
- 323. HDL AND REVERSE CHOLESTEROL TRANSPORT
- 324. HDL AND REVERSE CHOLESTEROL TRANSPORT
- 325. CATABOLISMUL HDL HDL sînt catabolizaţi la nivelul
- 326. LDL-R 50% of HDL
- 328. PATOLOGIA Hipolipoproteinemia familiară – deficit de HDL
- 329. EXPLORĂRI UZUALE ALE METABOLISMULUI LIPIDIC
- 330. EXPLORĂRI UZUALE Pentru efectuarea determinărilor: pacienţii
- 331. EXPLORĂRI UZUALE Aspectul plasmei Lipidele totale Colesterol
- 332. 1. ASPECTUL PLASMEI Clar Normal Hipercolesterolemie
- 333. COLESTEROLUL TOTAL normal
- 334. HIPERCOLESTEROLEMIA FAMILIALĂ
- 335. LDL-COLESTEROLUL Formula de calcul: LDL-col (mmol/L) =
- 336. TRIGLICERIDELE normal
- 337. HDL-COLESTEROLUL Valorile normale: Bărbaţi –
- 339. LIPOPROTEINA (A) LP modificată din clasa
- 340. DISLIPOPROTEIDEMIE SAU DISLIPIDEMIE modificarea c% lipidelor totale
- 341. HIPERLIPIDEMIILE Hiperlipidemiile primare sunt de natură genetică,
- 342. HLP PRIMARE după Fredrickson, în funcţie
- 345. XANTOAME TENDINOASE
- 346. HIPERLIPIDEMIILE SECUNDARE Diabetul zaharat: Lipsa de insulină: lipoliza activată, lipogeneza inhibată
- 347. DISLIPIDEMIA DIABETICĂ Mărirea concentraţiei AGL ( activarea
- 348. HIPERLIPIDEMIA ALCOOLICĂ: se observă o lipidemie alimentară
- 349. Alcoolul amplifică sinteza şi secreţia pre-β-lipoproteinelor determinate
- 350. SUBSTANŢE LIPOTROPE Ficatul este un organ
- 351. OBEZITATEA Excesul de aport caloric, raportat
- 352. OBEZITATEA Evaluarea obezităţii se efectuează prin calcularea
- 353. TIPUL OBEZITĂŢII Se stabileşte în funcţie de
- 354. RISCUL CARDIOVASCULAR ÎN FUNCŢIE DE CT ŞI SEX
- 355. DISLIPIDEMIA DIN OBEZITATE Hipertrigliceridemie; Hiperlipidemie postprandială; Scăderea
- 356. ATEROSCLEROZA l Se caracterizează prin depunerea Col
- 357. FACTORI DE RISC MAJOR AL ATEROSCLEROZEI Dislipidemiile
- 358. FACTORI DE RISC MODERAT AL ATEROSCLEROZEI obezitatea
- 359. ATEROSCLEROZA Patogenie: leziunea pereţilor endoteliali Infiltrarea LDL
- 360. ROLUL LIPOPROTEINELOR ÎN ATEROSCLEROZĂ
- 361. ROLUL LIPOPROTEINELOR ÎN ATEROSCLEROZĂ
- 363. PROFILUL LIPIDIC ATEROGEN Majorarea LDL (îndeosebi LDL
- 364. HIPOLIPOPROTEIDEMIILE PRIMARE (EREDITARE, FAMILIALE) sunt datorate genelor
- 365. HIPOLIPOPROTEIDEMIILE PRIMARE afecţiunea Tangier - cauzată
- 366. A BETA-LIPOPROTEINEMIA FAMILIALĂ Afecţiune transmisă autosomal
- 368. VITAMINELE sunt substanţe biologic active necesare pentru
- 369. AVITAMINOZE ŞI HIPOVITAMINOZE avitaminoze - lipsa vitaminelor
- 370. PROVITAMINE - predecesorii vitaminelor, care în organism
- 371. ANTIVITAMINELE – substanţe (inclusiv şi unele medicamente)
- 372. ANTIVITAMINELE Preparatele sulfanilamidice prezintă antivitamine pentru microorganisme
- 373. VITAMINELE LIPOSOLUBILE A, D, E, K: Vitamine
- 374. VITAMINA A vitamina creşterii, retinol, antixeroftalmică La
- 375. VITAMINA A
- 376. METABOLISM Digestia are loc în intestin Absorbţia
- 378. ROLUL VITAMINEI A Retinol – acţiune hormonală
- 379. METABOLISMUL All-trans-retinol retinol RBP Retinol-RBP
- 380. ROLUL VITAMINEI A
- 381. CICLUL VIZUAL Vit A este o
- 382. CICLUL VIZUAL Când rodopsina este expusă la
- 384. CICLUL VIZUAL O parte din trans-retinal este
- 385. ROLUL RETINOLULUI necesar pentru reproducere si crestere
- 386. RETINOL ŞI ACIDUL RETINOIC necesari
- 387. ALTE EFECTE: La oxidarea AG Biosinteza gliceridelor,
- 388. VITAMINA A Se conţine în produse
- 389. CARENŢA VITAMINEI A se caracterizează prin: Orbirea
- 390. VITAMINA D (ANTIRAHITICĂ) Se cunosc mai multe
- 391. SINTEZA COLECALCIFEROLULUI (DERM ŞI EPIDERM)
- 393. METABOLISM: În intestin se supune acţiunii
- 394. CALCITRIOLUL În ficat vit D3 suferă
- 396. EFECTELE VITAMINEI D La nivel de intestin:
- 397. VITAMINA D (ANTIRAHITICĂ) Carenţa -
- 399. SURSE DE VITAMINA D ficatul de
- 400. VITAMINA E (α-TOCOFEROL) Structura chimică: are la bază tocolul (5,7,8-trimetiltocol)
- 401. METABOLISMUL VITAMINEI E Absorbită la nivelul intestinului
- 402. ROLUL VITAMINEI E Este antioxidant
- 403. DEFICIENŢA VITAMINEI E are ca urmare
- 405. VITAMINA E Se conţine în uleiuri vegetale
- 406. VITAMINA K (ANTIHEMORAGICĂ) K1- filochinona (izolată
- 407. VITAMINA K – STRUCTURA CHIMICĂ
- 408. ROLUL VITAMINEI K Participă la coagularea sângelui
- 409. ROLUL VITAMINEI K
- 410. CARENŢA VITAMINEI K Carenţa acestei vitamine
- 411. CARENŢA VITAMINEI K Hemoragiile pot apărea când
- 412. Antivitaminele acestei vitamine, de ex. dicumarolul sunt
- 413. LIPIDOZE TISULARE Boli ereditare, caracterizate
- 414. LIPIDOZE TISULARE 1. Neimann-Pick - deficienţa de
- 415. LIPIDOZE TISULARE 2. afecţiunea Tay-Sachs – lipsa
- 416. LIPIDOZE TISULARE 3. Boala Krabbe – deficit
- 417. LIPIDOZE TISULARE 4. boala Gaucher – deficit
- 418. LIPIDOZE TISULARE 5. leucodistrofia metacromatică – deficit
- 419. LIPIDOZE TISULARE boala lui Fabry (lipogranulomatoza) -
Слайд 1
LIPIDELE: ROLUL;
STRUCTURA CHIMICĂ;
PROPRIETĂŢI.
MEMBRANELE BIOLOGICE.
DIGESTIA ȘI ABSORBȚIA LIPIDELOR

Слайд 2OBIECTIVELE:
Funcţiile biologice ale lipidelor.
Clasificarea lipidelor( structurală, funcţională, după proprietăţile fizico-chimice).
Lipidele de
Lipidele protoplasmatice – fosfogliceridele, sfingolipidele, glicolipidele, colesterolul- structura, proprietăţilefizico-chimice, rolul biologic.
Membranele biologice: funcţiile şi structura – modelul S.G.Sînger şi G.L.Nicolson;
Proprietăţile fundamentale – fluiditatea, molilitatea, permeabilitatea selectivă, asimetria, autoasamblarea şi autorapararea;
Diversitatea şi specificitatea structurilor şi funcţiilor diferitor membrane biologice.
Importanţa lipidelor în alimentaţie.
Digestia şi absorbţia lipidelor în tractul gastrointestinal.
Acizii biliari – clasificarea, structura, funcţiile lor. Metabolismul acizilor biliari ( noţiuni generale).
Resintaza lipidelor în enterocite. Soarta lipidelor resintetizate.
.Lipidele de rezervă – acilgliceridele, reprezentanţii,")
Слайд 3LIPIDE
Substanţe organice, insolubile în apă dar solubile în solvenţi organici (eter,
.")
Слайд 4ROLUL
Energetic - principala formă de depozitare şi de transport a rezervelor
Structural - constituienţi structurali ai membranelor celulare şi intracelulare
Izolator: termo; mecano-; electroizolator
Pot avea efecte biologice: sunt vitamine, hormoni, prostaglandine

Слайд 5CLASIFICAREA LIPIDELOR
1. DUPĂ STRUCTURA CHIMICĂ (HIDROLIZĂ)
Monomerii lipidici sau lipidele monocomponente, nesaponifiabile
1.
a. saturaţi – palmitic, stearic, etc.;
b. nesaturaţi:
* monoenici – palmitoleic, oleic, etc.
* polienici – linolic, linolenic,arahidonic,
2. Alcoolii, aminoalcoolii, cetonele şi aldehi-
dele superioare;
3. izoprenoizii şi derivaţii lor – carotenoizii, vit. A, vit. K
Monomerii lipidici sau lipidele monocomponente, nesaponifiabile1. Acizii graşi: a. saturaţi")
Слайд 6CLASIFICAREA LIPIDELOR
DUPĂ STRUCTURA CHIMICĂ
II. Lipidele policomponente, saponifiabile
Simple – esteri ai acizilor
a. acilglicerolii – mono-, di- şi trigliceridele
b. cerurile

Слайд 7CLASIFICAREA LIPIDELOR
DUPĂ STRUCTURA CHIMICĂ
Complexe – esteri ai acizilor graşi şi
alcoolilor, ce conţin şi alte substanţe, ca fosfat, colină, etc.:
a. Fosfolipidele:
I. fosfogliceridele: fosfatidilserinele;
fosfatidilcolinele;
fosfatidiletanolaminele; fosfatidilinozitolii,
etc.
II. sfingolipidele - sfingomielina

Слайд 8CLASIFICAREA LIPIDELOR
DUPĂ STRUCTURA CHIMICĂ
b. Glicolipidele:
gangliozidele; sulfatidele, etc.

Слайд 9CLASIFICAREA LIPIDELOR
DUPĂ STRUCTURA CHIMICĂ
III. Steroizii
Colesterolul şi colesteridele
Hormonii steroizi
Acizii biliari

Слайд 10CLASIFICAREA LIPIDELOR
2. DUPĂ PROPRIETĂŢILE FIZICO-CHIMICE:
Lipide nepolare:
cerurile,
trigliceridele,
colesteridele;
Lipidele polare:
fosfogliceridele,
glicolipidele,
sfingomielinele,
colesterolul.


Слайд 12CLASIFICAREA LIPIDELOR
3. DUPĂ ROLUL BIOLOGIC:
1. Lipide structurale sau protoplasmatice – din
2. Lipidele de rezervă – din ţesutul adipos (preponderent) – trigliceridele.
3. Formele de transport sangvin ale lipidelor – lipoproteinele plasmatice (LPP)

Слайд 13DISTRIBUȚIA LIPIDELOR
Lipidele constituie 10-20% din masa corpului.
10-12 kg lipide:
2-3 kg
98% - concentrate în ţesutul adipos


Слайд 15AG – STRUCTURA, PROPRIETĂŢI
AG- derivaţii hidrocarburilor alifatice ce conţin gruparea carboxil
Clasificarea
1. După nr atomilor de C:
- AG cu nr par
- AG cu nr impar
2. După gradul de saturare:
saturaţi
nesaturaţi
3. După rolul fiziologic:
esenţiali (linoleic şi linolenic)
neesenţiali

Слайд 16STRUCTURA CHIMICĂ A AG SATURAȚI
CH3-(CH2)2-COOH butiric –C4
CH3-(CH2)4-COOH caproic –C6
CH3-(CH2)6-COOH caprilic – C8
CH3-(CH2)8-COOH caprinic – C10
CH3-(CH2)10-COOH lauric –C12
CH3-(CH2)12-COOH miristic – C14
CH3-(CH2)14-COOH palmitic –C16
CH3-(CH2)16-COOH stearic – C18
2-COOH butiric –C4 CH3-(CH2)4-COOH caproic")
Слайд 17STRUCTURA CHIMICĂ A AG MONONESATURAŢI
AG nesaturaţi conţin una sau mai
Au configuraţia cis
a. palmitooleic (C16:1 cisΔ9)
CH3- (CH2)5- CH=CH-(CH2)7-COOH
a. oleic (C18:1 cisΔ9)
CH3-(CH2)7-CH=CH-(CH2)7-COOH

Слайд 18AG POLINESATURAŢI
a.linoleic (C18:2cis Δ 9,12 )
CH3-(CH2)4-CH=CH-CH2-CH=CH-(CH2)7- COOH
a. linolenic (C18:3 cis Δ
CH3-CH2-CH=CH-CH2-CH=CH-CH2-CH=CH-(CH2)7-COOH
a.arahidonic (C20:4,cis Δ 5,8,11,14)
CH3-(CH2)4-CH=CH-CH2-CH=CH-CH2-CH=CH-CH2-CH=CH-(CH2)3-COOH
CH3-(CH2)4-CH=CH-CH2-CH=CH-(CH2)7- COOHa. linolenic (C18:3 cis Δ 9,12,15)CH3-CH2-CH=CH-CH2-CH=CH-CH2-CH=CH-(CH2)7-COOH a.arahidonic (C20:4,cis")
Слайд 19PROPRIETĂŢILE AG
AG saturaţi până la C10 – sunt lichizi; mai sus
În cristale catenele hidrocarburilor saturate au configuraţia de zig-zag
AG nesaturaţi prezintă izomerie cis-trans însă formele naturale ale acestor – sunt izomerii cis
AG – puţin solubili în apă, solubilitatea scăzând odată cu lungimea catenei
AG nu se află în stare liberă în celule şi ţesuturi ci sunt legaţi covalent
AG nesaturaţi au t de topire mai joasă comparativ cu cei saturaţi--- lungimea mică şi prezenţa legăturilor duble amplifică fluiditatea

Слайд 20PROPRIETĂŢILE CHIMICE
AG saturaţi şi nesaturaţi pot forma:
Esteri (gliceride, fosfogliceride)
Săruri (săpunuri cu
Amide (sfingolipide)
AG nesaturaţi adiţionează la nivelul dublei legături – hidrogen, halogeni (Br2; Cl2), tiocianatul (SCN)2, gruparea hidroxil
AG nesaturaţi reacţionează cu oxigenul formând peroxizi (substanţe toxice la care se alterează gustul şi mirosul (râncezire)
Săruri (săpunuri cu proprietăţi tensioactive)Amide (sfingolipide)AG nesaturaţi")
Слайд 21ROLUL AG:
Sursă de energie pentru miocard si pentru mușchii striați
AG sunt insolubili in apa si de aceea circula in plasma legați cu albuminele, formă cunoscută sub denumirea de acizi grasi liberi (AGL)

Слайд 22GLICERIDE
( ACILGLICEROLII, GRĂSIMI NEUTRE)
Sunt esteri ai glicerolului cu AG
După nr
monogliceride;
digliceride;
trigliceride
Sunt esteri ai glicerolului cu AGDupă nr grupărilor alcoolice esterificate din")



Слайд 26ROLUL TAG
Constituie grăsimea de rezervă din ţesuturi - rol energetic
Izolator
TAG din
Oleic-45%, palmitic-25%, linoleic – 8%, palmitooleic – 7%, stearic- 7% şi alţii -7%
Sunt:
simpli R1=R2=R3
micşti

Слайд 27PROPRIETĂŢILE TAG
Sunt determinate de natura şi numărul de AG constituienţi
Insolubile în
TAG ce conţin AG saturaţi – solide(grăsimi, unt)- de origine animală
TAG ce conţin AG nesaturaţi – consistenţă lichidă (uleiurile vegetale)
Prin hidroliza enzimatică (se produce în intestin sub acţiunea lipazei) se scindează în glicerol şi 3AG
Prin hidroliza alcalină (KOH, NaOH) –glicerol+săpunuri
Prezenţa AG nesaturaţi – proprietatea de a adiţiona halogenii la dublele legături
Sub acţiunea luminii, căldurii – se autooxidează

Слайд 28GLICEROFOSFOLIPIDELE (FOSDOGLICERIDE)
Derivaţi ai acidului fosfatidic
Reprezentanţii:
fosfatidilcolina (lecitina)
Fosfatidiletanolamina (cefalina)
Fosfatidilserina
Fosfatidilinozitol
Plasmalogeni
Derivaţi ai acidului fosfatidicReprezentanţii: fosfatidilcolina (lecitina)Fosfatidiletanolamina (cefalina)Fosfatidilserina FosfatidilinozitolPlasmalogeni")

")
")



")
Слайд 36ROLUL FOSFOGLICERIDELOR
Rolul:
Structural
Fosfatidilcolina cu 2 resturi de palmitil este componentul principal al
Fosfatidiletanolamina – sunt abundente în ţesutul nervos
Fosfatidilinozitolul- rol în procesul de transmitere a semnalelor extracelulare
Cardiolipina: component al surfactantului pulmonar şi principala componentă a membranei MC interne

Слайд 37PROPRIETĂŢILE FL
Caracter amfipatic (lecitinile şi cefalinele au structură bipolară: prezenţa resturilor

Слайд 38PROPRIETĂŢILE FL
2. Proprietăţi tensioactive puternice
3. În apă se dizolvă formând agregate,micelii,
4. Fosfatidilserinele au sarcină negativă
5. fosfatidilinozitolul- are un caracter puternic acid

Слайд 39SFINGOLIPIDELE
Derivaţi ai sfingozinei
Sunt abundente în substanţa nervoasă albă, cenuşie, nervii periferici
Rolul:
Sunt surse energetice (datorită AG)
Componente ale membranei celulare (nervoasă)
Participă la coagularea sângelui
Sunt transportori de e şi componente ale mitocondriilor, unde au loc procesele de O/R
Rol în fiziologia SNC

Слайд 40SFINGOLIPIDELE
Proprietăţi; amfioni, au caracter amfipatic
2 categorii:
sfingomielina şi
glicosfingolipidele




Слайд 44SFINGOMIELINA
Sfingomielina: constituent al fibrelor nervoase mielinice
Sfingomielinele au în
componenţă
fosfocolină
şi se găsesc
în membranele
plasmatice.

Слайд 45GLICOLIPIDE
(GLICOSFINGOLIPIDELE)
- includ ceramida legată glicozidic de monozaharide (hexoze) sau
nu cuprind fosfor
Cerebrozide
Sulfatide
Gangliozide
 - includ ceramida legată glicozidic de monozaharide (hexoze) sau oligozaharidenu cuprind fosforCerebrozide SulfatideGangliozide")
Слайд 46GLICOLIPIDE
(GLICOSFINGOLIPIDELE)
Cerebrozide – se află abundent în substanţa albă a creierului;
Structură: ceramida (AG-24C)+βGal (mai rar β Gl)
Proprietăţi: nu au sarcină electrică; au caracter amfipatic
Cerebrozide – se află abundent în substanţa albă a creierului; în nervi. Structură: ceramida")

Слайд 48
3. Gangliozide:
sunt prezente în toate ţesuturile în cantităţi mici, mai
au rol în transmiterea impulsului nervos dea lungul sinapselor (fac parte din receptorii acetilcolinei)
la PH=7 au sarcină negativă
Structură: ceramida (AG-stearic)+oligozaharid (Gal+Gl) legat de 1, 2 sau 3 resturi de a sialic NANA +Nacetilgalactoz (glucoz)amină
Ceramida-Gl-Gal–GalNHAc+Gal
NANA
N acetilneuraminic mono-; di-; tri- sialogangliozid
Componenţa glucidică se găseşte pe faţa externă a membranelor


")
Слайд 51COLESTEROLUL
Sterol C27
Origine animală (lipseşte în plante)
Solid, alb, insolubil în apă
Rol:
1.
2.Precursorul tuturor compuşilor steroidici (provitaminei D, acizilor biliari, hormonilor steroizi)
2 forme: esterificat şi neesterificat
Solid, alb, insolubil în apă Rol:1. structural2.Precursorul tuturor compuşilor steroidici")


Слайд 54ACIZII BILIARI
Se sintetizează în ficat din colesterol
Structură: steroizi C24 cu
AB primari:
colic,
chenodezoxicolic

Слайд 55Acidul colanic
Structura chimică a AB
Acid colic
(3,7,12-trihidroxicolanic)
НО
НО
ОН
3
7
12
AB sunt derivați ai acidului
Acidul dezoxicolic (3,12-dihidroxicolanic)
Acidul chenodezoxicolic (3,7-dihidroxicolanic)
Acidul litocolic (3-hidroxicolanic)
НОНООН3712AB sunt derivați ai acidului colanicAcidul dezoxicolic (3,12-dihidroxicolanic)Acidul chenodezoxicolic")
Слайд 56ACIZII BILIARI
La PH alcalin se găsesc sub formă de săruri biliare:conjugaţi
Glicocolic sau taurocolic
Glicochenodezoxicolic sau taurochenodezoxicolic
Astfel în bilă se găsesc doar sărurile biliare

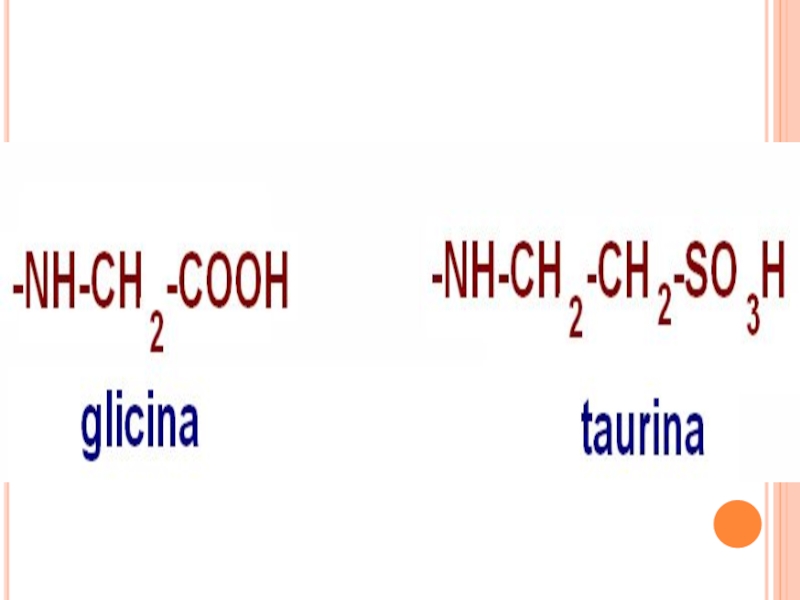

Слайд 59ACIZII BILIARI
Bacteriile intestinale:
1. pot înlătura Gly şi taurina din sărurile biliare
2. pot converti AB primari în AB secundari prin îndepărtarea unei grupări OH
A colic ---- a dezoxicolic
A chenodezoxicolic------a litocolic

Слайд 60ACIZII BILIARI
Rolul:
proprietăţi de solubilitate (micele în apă şi au capacitatea
Transport (AG)
Emulgatori
Activează lipaza pancreatică

Слайд 61MEMBRANELE BIOLOGICE
Structuri superorganizate
constituienţii de bază - proteinele şi lipidele.
CARACTERISTICI:
despart celula
reprezintă o barieră de permeabilitate selectivă
conţin pompe ionice şi canale cu permeabilităţi specifice (sisteme de transport)
sunt flexibile, labile, permanent se reînnoiesc
sunt elastice şi dure la deformare

Слайд 62FUNCŢIILE BIOLOGICE ALE MEMBRANELOR:
de separare (de barieră)
de integrare sau asociere
de transport (pasiv, activ);
osmotică – de concentrarea a substanţelor între spaţiile intra- şi extracelulare
electrică – repartizarea neuniformă a sarcinilor pe ambele părţi ce duce la apariţia diferenţei de potenţial electric;
de integrare sau asociere – asociază procesele biochimice")
Слайд 63FUNCŢIILE BIOLOGICE ALE MEMBRANELOR:
de transformare a energiei – asigură transformarea
de recepţie – prin intermediul receptorilor ele reglează schimbul de informaţie între celule şi mediul extern;
reglatoare; metabolică – participă la formarea AMPc; GMPc; enzimele membranare – în diverse transformări metabolice;
antigenică – glicoproteinele membranare determină capacitatea de formare a anticorpilor;

Слайд 64STRUCTURA MB
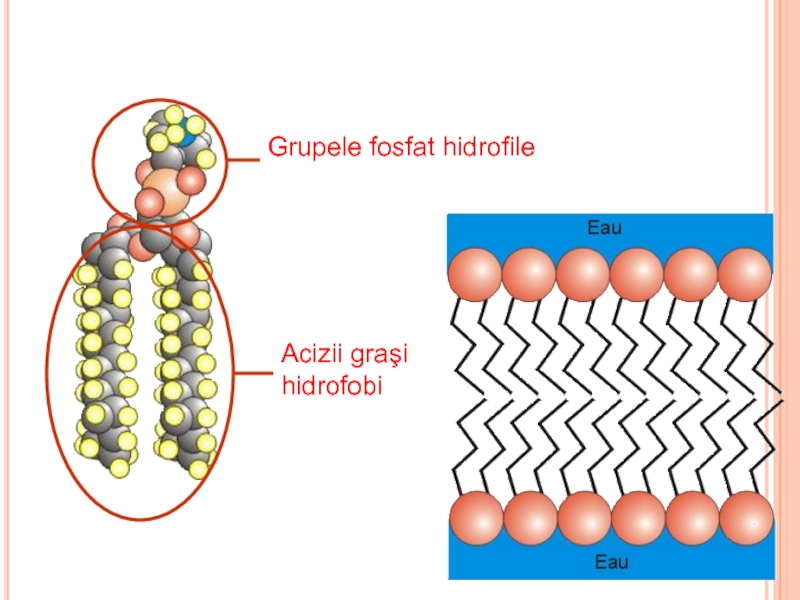
Componenţi de bază:
lipide
proteine (1:4 pînă la 4:1)
Glucide
Lipidele: conţin
GlucideLipidele: conţin grupe hidrofobe (nepolare) şi")

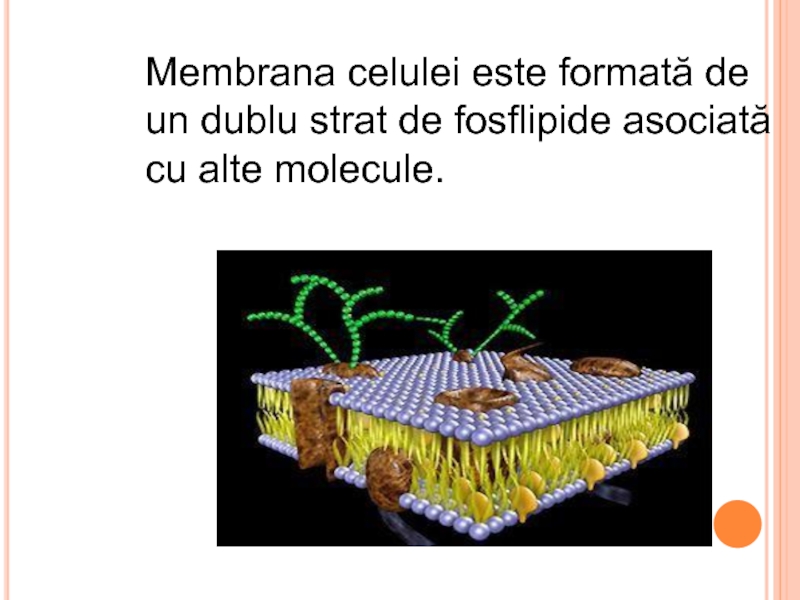
Слайд 67MEMBRANELE CELULELOR EUCARIOTE CONŢIN COLESTEROL
colesterol
celule animale
ergosterol
celule vegetale

Слайд 69ROLUL LIPIDELOR STRUCTURALE:
Fosfolipidele şi glicolipidele – constituienţii cheie ai membranelor –
1. formează spontan şi rapid bistraturile lipidice stabilizate prin legături hidrofobe între părţile nepolare ale moleculelor lor (sunt necovalente şi cooperative)
2. îndeplinesc funcţia de barieră a permeabilităţii şi asigură crearea compartimentelor unice

Слайд 70PROTEINELE MEMBRANARE
După localizare:
- periferice (extrinseci) - sunt ataşate la exteriorul
- integrale (intrinseci) - ce trec prin stratul lipidic; aceste proteine penetrează parţial sau total stratul dublu de lipide. Proteinele care străbat bistratul lipidic de la o faţă la altă se numesc proteine transmembranare.
 - sunt ataşate la exteriorul stratului dublu lipidic, unde")
Слайд 71PROTEINELE MEMBRANARE
Proteinele periferice sunt legate de membrane prin forţe electrostatice şi
Proteinele integrate - amfipatice, conţin AA nepolari – formează cu lipidele membranei legături hidrofobe

Слайд 72ROLUL PROTEINELOR MEMBRANARE
asigură transportul,
participă în transmiterea informaţiei,
rol în transformările de

Слайд 73GLUCIDELE MEMBRANARE
Sunt sub formă de glicolipide şi glicoproteine
Sunt situate pe suprafaţa
Rolul:
asigură recunoaşterea şi adeziunea intercelulară – servesc în calitate de receptori moleculari;
favorizează menţinerea asimetriei membranelor biologice
au funcţia de identificare intercelulară, de detectare a celulelor străine sistemului imun
determină transportul proteinelor membranare în locusurile necesare

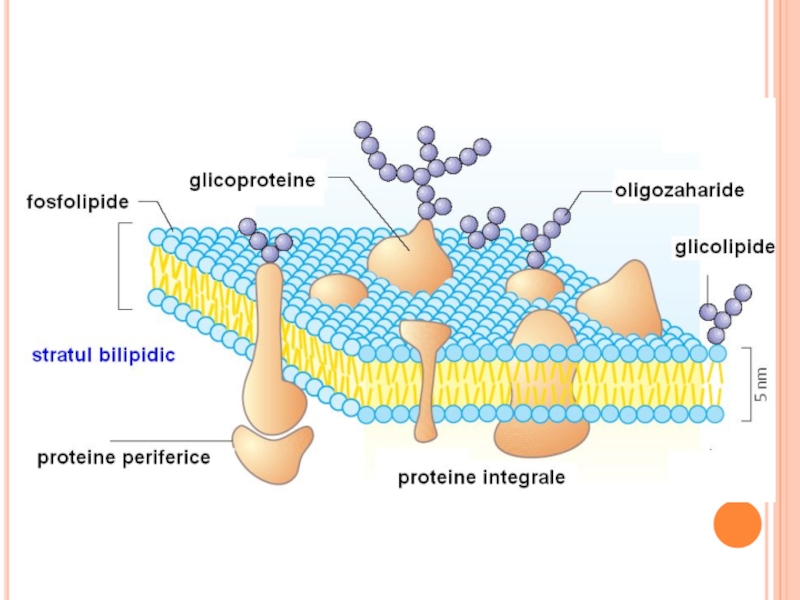
Слайд 74Lanţul de glucide ataşat pe lipide (glicolipide) sau pe proteine (glicoproteine)
Glicoproteinele
 sau pe proteine (glicoproteine)Glicoproteinele sunt foarte variate. Nu")
Слайд 76Modelul mozaic- fluid:
Bistratul de fosfolipide
Moleculele se deplasează permanent în raport
Proteinele periferice şi integrale
Polizaharidele ataşate pe lipide sau proteine
Colesterolul între fosfolipide
Propus în1972 de către Sanger şi Nicolson

Слайд 77PROPRIETĂŢILE MEMBRANELOR BIOLOGICE.
Fluiditatea - starea în care se află lipidele membranare
Reglatorul fluidităţii –
AG nesaturaţi -la procariote
şi Col -eucariote
Situat între catenele acil, Col:
Previne cristalizarea şi alipirea lor, mărind fluiditatea MC
blochează mobilitatea catenelor acil, micşorând fluiditatea MC
Col menţine fluiditatea la un nivel mediu

Слайд 78PROPRIETĂŢILE MEMBRANELOR BIOLOGICE.
Asimetria:
1. repartizarea neuniformă a lipidelor şi proteinelor pe suprafaţa
Ex.: membrana eritrocitului:
Partea internă- fosfatidiletanolamină şi fosfatidilserină
Partea externă – fosfatidilcolină şi sfingomielină
Asigură orientarea proteinelor membranare în bistrat
2. La suprafaţa externă este prezentă componenta glucidică (glicocalixul), pe când pe partea internă ele practic lipsesc.
3. Sistemele de transport din membrană funcţionează într-o singură direcţie (Na-K-ATP-aza)

Слайд 79DISTRIBUŢIA LIPIDELOR ESTE ASIMETRICĂ
exemple de la membrane plasmique
Fosfatidilserine
Fosfatidietanolamina
Fosfatidilcolina
Glicolipide
exterior
0
10
90
100
100
90
10
0

Слайд 80PROPRIETĂŢILE MEMBRANELOR BIOLOGICE.
Mobilitatea
Moleculele lipidelor membranei se află în mişcare continuă.
Proteinele membranare tot posedă mobilitate. Ele parcă plutesc în pătura lipidică, deplasându–se în procesul difuziunii laterale.

Слайд 81Proprietăţile membranelor biologice Permeabilitatea selectivă
Dublu strat de lipide este permeabil:
Pentru
Pentru molecule liposolubile (hidrofobe, nepolare)
Dublu strat de lipide este impermeabil:
Pentru moleculele mari şi pentru majoritatea moleculelor solubile în apă.
Pentru ioni (K+, Cl-, Na+ )

Слайд 82se realizează:
Prin transport pasiv (independent de energie)
Prin transport activ ( dependent
Transportul membranar
Prin transport activ ( dependent de energie) Transportul membranar")


Слайд 85O substanţă difundează conform gradientului său de concentraţie: din zona mai

Слайд 86Viteza de difuzie se modifică la:
Creşterea temperaturii mediului
La creşterea diferenţei de
La micşorarea permeabilităţii membranei (are mai puţine canale)

Слайд 87Se realizeaza:
prin dublul strat lipidic: pentru moleculele puternic liposolubile (acizi grasi,
Prin canale ionice si, respectiv, a apei prin canale de apa (acvaporine).
Difuzia simplă
 si a moleculelor")

Слайд 89a) transportul anionilor;
b) transportul ureei;
c) transportul glicerolului;
d) transportul unor neelectroliţi prin membrana eritrocitului;
e) transportul glucozei;
f) transportul aminoacizilor
Prin difuziune facilitată se realizează:
 transportul anionilor;b) transportul ureei;c) transportul glicerolului;d) transportul unor neelectroliţi prin membrana eritrocitului;e) transportul glucozei;f) transportul aminoacizilor – prin plasmalema celulelor.Prin")

 spre cea hipertonică (concentrată)")

Слайд 93Transport activ
Are nevoie de SURSĂ DE ENERGIE
Se poate face CONTRA
Necesită un TRANSPORTOR (proteină de transport)
Se aseamănă cu difuza facilitată (necesită un transportor) DAR:

Слайд 94 Transport activ – in sens contrar gradientilor; necesita consum de
➔ Transport activ primar
Pompe ionice → ioni
➔ Transport activ secundar
simport → ioni, molecule mari
antiport → ioni, molecule mari
3. Macrotransport
➔ Trafic vezicular → proteine, alte molecule mari
Transport activ

Слайд 95pompa Na+/K+ (Na+/K+-ATP-aza) - transportă 3Na+ spre exterior si 2K+ spre interior, pentru fiecare
pompa H+/K+ (H+/K+ - ATP-aza) - este prezentă în celula parietală gastrică unde are rol în formarea HCl, scoate din celula 1H+ și introduce 1K+,
Transport activ primar
 - transportă 3Na+ spre exterior si 2K+ spre interior, pentru fiecare moleculă de ATP hidrolizată pompa")
Слайд 963. pompa de Ca2+ (Ca 2+-ATP-aza) - este caracteristică fibrei musculare:
- la nivelul sarcolemei;
- la nivelul membranei reticulului sarcoplasmatic: asigură recaptarea Ca2+ în principalul depozit de Ca2+intracelular.
- Rolul pompelor de Ca2+ este cel de scădere a concentratiei Ca2+ citosolic de la 10-5 M la 10-7 M, necesara relaxării musculare.
Transport activ primar
 - este caracteristică fibrei musculare:- la nivelul sarcolemei; rol: expulzeaza Ca2+ in mediul")
Слайд 974. pompele protonice (H+-ATP-azele) se întâlnesc:
- la nivelul membranelor mitocondriale,
- la nivelul polului apical al nefrocitului distal, asigurând eliminarea renală a H+, cu rol in menținerea echilibrului acido-bazic.
.
Transport activ primar
 se întâlnesc: - la nivelul membranelor mitocondriale, fiind implicate în transportul electronilor")
Слайд 98Se realizeaza cu ajutorul proteinelor transportor
● Transportul unei specii moleculare,
● Variante:
– simport (transportul ambelor specii in acelasi sens, relativ la membrana)
– antiport (transportul celor doua specii in sens opus, relativ la membrana)
● Exemple:
– simport Na+ /glucoza din lumenul intestinal in celulele epiteliale
– Antiport 2Na+ /Ca2+ in celulele musculare cardiace (Ca2+ transportat din interiorul spre exteriorul celulei)
Transport activ secundar


Слайд 100Endocitoza - transportul intracelular al moleculeor mari, polare
membrana citoplasmatică se
- poate fi:
fagocitoză – transportul materiei solide
pinocitoză – transportul materiei în stare lichidă
endocitoză mediată de receptori membranari
Exocitoza - procesul invers endocitozei, vezicule formate în interiorul celulei sunt transportate spre exterior şi golite - implicată în producerea hormonilor sau enzimelor digestive
MACROTRANSPORT:


Слайд 103Canalele sunt foarte specifice: o singură substanţă bine precizată se poate
Deci, dacă e important ce substanţă poate traversa membrana = permeabilitate selectivă.


Слайд 106Unele canale se pot deschide şi închide:
Canal închis
Membrana
Canal deschis
Proteinele ce
formează


Слайд 108IMPORTANŢA LIPIDELOR ÎN ALIMENTAŢIE.
Aportul alimentar de lipide necesar zilnic -
Principalele lipide ale raţiei alimentare sunt: TAG (90%); FL; Col liber şi esterificat.
Lipidele alimentare sunt sursa AG indespensabili – linoleic şi linolenic.
Funcţionarea normală a organismului necesită un consum minim obligatoriu de vitamine liposolubile ( A,D,E,K )

Слайд 109DIGESTIA LIPIDELOR ALIMENTARE
Digestia lipidelor începe în stomac sub acțiunea lipazei linguale
Ambele acționează asupra TAG care conţin AG cu catenă scurtă (mai puţin de 12 atomi de C, prezente în grăsimile din lapte)

Слайд 110DIGESTIA LIPIDELOR ALIMENTARE
La nou născut, ambele lipaze „acide” sunt importante în
Digestia TAG din lapte are loc în poziţia 3, rezultând 1,2 diglicerid

Слайд 111DIGESTIA LIPIDELOR ÎN INTESTIN
pH alcalin - bicarbonaţii sucului pancreatic
prezenţa AB
a. emulsionarea lipidelor alimentare,
b. activarea enzimelor lipolitice;
c. absorbţia produselor finale ale digestiei.
3. Prezenţa enzimelor lipolitice: lipaza, colipaza, fosfolipazele, colesterolesteraza
")
Слайд 112DIGESTIA LIPIDELOR ÎN TGI
În intestin - chimul din stomac este neutralizat
Grăsimile se supun emulsionării sub acţiunea sărurilor AB.
Peristaltismul intestinului ajută la fărîmiţarea picăturilor mici de grăsime, iar sărurile AB împiedică contopirea picăturilor mici de grăsime

Слайд 113DIGESTIA TAG
lipazei pancreatice:
Este o glicoproteidă, PH=8-9
Lipaza pancreatică acționează împreună cu colipaza

Слайд 114DIGESTIA FOSFOGLICERIDELOR
fosfolipazele (A1, A2, C, D) pînă la glicerol, acizi
 pînă la glicerol, acizi graşi, acid fosforic şi")
Слайд 115DIGESTIA FOSFOGLICERIDELOR
Cea mai activă este fosfolipaza A2 (se activează de tripsină,
fosfatidilcolina-------lizolecitina (proprietăţi detergente puternice; participă la solubilizarea lipidelor în intestin)
AG de la nivelul atomului C1 poate fi îndepărtat de lizofosfolipază, rezultând gliceril-fosforil-colina ce poate fi excretată în scaun, degradată sau reabsorbită.

Слайд 116DIGESTIA COL
Colesterolesteraza pancreatică (activată de AB) scindează Col esterificat în Col
 scindează Col esterificat în Col şi AG")
Слайд 118PRODUSELE FINALE ALE DIGESTIEI LIPIDELOR:
Acizii graşi
Monogliceridele
Colesterolul
Lizolecitina

Слайд 119REGLAREA DIGESTIEI
Colecistochinina (CCK) determină contracţia vezicii biliare şi eliberarea bilei (un
reduce motilitatea gastrică, astfel conţinutul gastric ajungând mai lent în intestinul subţire
 determină contracţia vezicii biliare şi eliberarea bilei (un amestec de AB, FL,")
Слайд 120REGLAREA DIGESTIEI
Secretina - stimulează eliberarea de către ficat şi pancreas a

Слайд 121ABSORBŢIA LIPIDELOR
La nivelul porţiunii superioare a intestinului subţire;
sunt absorbite prin difuzie
Prin difuzie simplă sunt transportaţi: AG cu catena scurtă și medie – sunt transportați la ficat de către albumine
AG cu catena lungă, 2 monoacilglicerolul, Col liber se absorb cu ajutorul sărurilor AB sub formă de micele

Слайд 122CICLUL ENTERO-HEPATIC
AG +AB formează o soluţie micelară şi pătrund în spaţiile
Sărurile AB rămîn în lumen, participînd la solubilizarea şi transportul altor lipide. Abia în porţiunea distală a ileonului sărurile AB se absorb printr-un mecanism activ.
Prin sistemul portal trec în ficat (se reînnoiesc)– bila --- intestin - circulaţia entero-hepatică a acizilor biliari.

Слайд 123MALABSORBȚIA LIPIDELOR
Malabsorbţia lipidelor caracterizată prin creşterea excreţiei lipidelor în scaun –
Biliară, hepatică (colestază, litiază biliară)
pancreatică (pancreatite)
intestinală (enterite)
Consecințe: malabsorbția vitaminelor liposolubile; AG esențiali; apare retard de creștere, tulburări neurologice și oftalmologice

Слайд 124RESINTEZA LIPIDELOR ÎN ENTEROCITE
Rolul: sinteza lipidelor specifice organismului uman din monomeri
Asigurată de:
#specificitatea enzimelor
#utilizarea ac. graşi endogeni
Se produc: trigliceride, colesteride şi fosfogliceride

Слайд 125RESINTEZA LIPIDELOR ÎN ENTEROCITE
MAG + 2 R-CO-S-CoA
Lizolecitină + R-CO-S-CoA lecitină + НS-CoA
Colesterol + R-CO-S-CoA colesterol esterificat + НS-CoA

Слайд 126RESINTEZA LIPIDELOR
Moleculele lipidelor reconstituite împreună cu cantităţi mici de proteină sunt
CM sunt secretaţi în vasele limfatice ce drenează intestinul şi la nivelul canalului toracic trec în plasmă.
CM")

Слайд 128OBIECTIVELE:
Biosinteza acizilor graşi:
saturaţi cu număr par de atomi de carbon;
nesaturaţi cu
Enzimele, coenzimele, reglarea.
Biosinteza TAG: substanţele iniţiale, enzimele şi coenzimele, reglarea.
Catabolismul triacilglicerolilor – reacţiile parţiale, enzimele, reglarea.
Oxidarea glicerolului – reacţiile parţiale, enzimele, coenzimale, reglarea, randamentul energetic al oxidării anaerobe şi aerobe.
Oxidarea acizilor graşi:
a) saturaţi cu număr par de atomi de carbon;
b) nesaturaţi cu număr par de atomi de carbon;
c) saturaţi cu număr impar de atomi de carbon;
d) în peroxizomi.
Reacţiile parţiale, enzimele, coenzimele, reglarea, randamentul energetic.
Metabolismul corpilor cetonici. Căile biosintezei şi utilizării lor – reacţiile parţiale, enzimele, coenzimele, reglarea. Rolul biologic al corpilor cetonici.

Слайд 129BIOSINTEZA AG
Sinteza AG şi încorporarea lor în TAG constituie mecanismul principal
Gl nu se mai transformă în glicogen dar în TAG:
Gl Piruvat Acetil CoA

Слайд 130BIOSINTEZA AG CUPRINDE MAI MULTE PROCESE:
Sinteza de novo cu formarea acidului
Elongarea acidului palmitic
Introducerea de legături duble în AG


Слайд 132PARTICULARITĂŢILE SINTEZEI AG
Are loc în citozol
Complex multienzimatic - acid gras sintaza
Ca iniţiator este acetil CoA (rezultat din glicoliză),
sursa majoră – malonil CoA
rolul reducător îi revine NADPH+H
Majoritatea ţesuturilor (ficat, rinichi, creier, plămân, glanda mamară, ţesut adipos) dispun de acest echipament enzimatic, dar ficatul deţine rolul principal în sinteza de novo a acidului palmitic;

Слайд 133ACID GRAS SINTAZA
= Palmitat sintetaza
E dimerică
Fiecare monomer e constituit din:
1 domeniu reprezentat de proteina transportatoare a grupărilor acil -ACP, care leagă covalent o moleculă de 4- fosfopanteteină


Слайд 135ACID GRAS SINTAZA
Fiecare monomer cuprinde 2 grupe SH:
–SH furnizat de
- SH - 4 fosfopantoteină din ACP: SH-Pant, (ataşată prin legătura fosfat-Ser)


Слайд 137ACID GRAS SINTAZA (AGS)
Datorită aranjării celor 2 monomeri – se
 Datorită aranjării celor 2 monomeri – se sintetizează simultan 2 molecule")
Слайд 138SINTEZA DE NOVO CU FORMAREA ACIDULUI PALMITIC
Etapele:
transferul lui Acetil CoA
Sinteza de malonil CoA
Sinteza acidului palmitic






Слайд 144III: SINTEZA ACIDULUI PALMITIC
Ciclu de reacţii este reluat: butiril-ACP se condensează
Catena AG creşte pînă la formarea palmitoil-S-ACP



Слайд 147
Biosinteza acizilor graşi saturaţi cu număr impar de atomi de carbon
Se

Слайд 148ELONGAREA AG
Localizată:
reticulul endoplasmatic
mitocondrii (acetil CoA, NADH+H)
AG este activat
La
AG este activat La acidul preexistent (palmitil CoA)")

Слайд 150BIOSINTEZA ACIZILOR GRAŞI NESATURAŢI
Se produce în reticulul endoplasmic;
Este catalizată de –
Se pot sintetiza doar acizi graşi mononesaturaţi din acizii graşi saturaţi corespunzători (acid palmitoleic din acid palmitic şi acid oleic din acid acid stearic);

Слайд 151BIOSINTEZA AG NESATURAŢI
Sistemul de desaturare are trei componente: o desaturază, citocromul
Se poate introduce o dubla legătură între C 9 şi C 10; o a doua legătură se poate introduce numai între dubla legătură preexistentă şi gruparea carboxil (de aceea acidul oleic nu se poate transforma în acid linoleic sau linolenic);
Electronii proveniţi de la NADPH sunt cedaţi cit b5 şi preluaţi în final de oxigen.
introduce gruparea hidroxil, urmată de dehidratare


Слайд 153BIOSINTEZA AG POLIENICI
Acidul linoleic şi linolenic sunt esenţiali
Organismul uman nu
posedă carbon -9,6,5,4 –desaturaze


Слайд 155SINTEZA TAG
2 căi:
calea monoacilglicerolului: are loc în peretele intestinal (enterocite) din
calea glicerolfosfatului: în toate ţesuturile (activă: ţesutul adipos şi ficat)
 din produşi absorbiţi (resinteza lipidelor).calea")
Слайд 1561. CALEA MONOACILGLICEROLULUI
TAG împreună cu FL, Col, proteine sunt incorparate




Слайд 160METABOLISMUL TAG
Are loc preponderent în ficat, ţesut adipos şi intestin
În plasmă
CM – transportă TG exogene – de la intestin la ţesuturi
VLDL – transportă TG endogene - de la ficat spre ţesuturi
Ele sunt hidrolizate de lipoproteinlipază în AG şi glicerol
AG – ţesutul adipos – sinteza TG

Слайд 161MOBILIZAREA TAG DIN ŢESUTUL ADIPOS
are loc în etape, pînă la glicerol

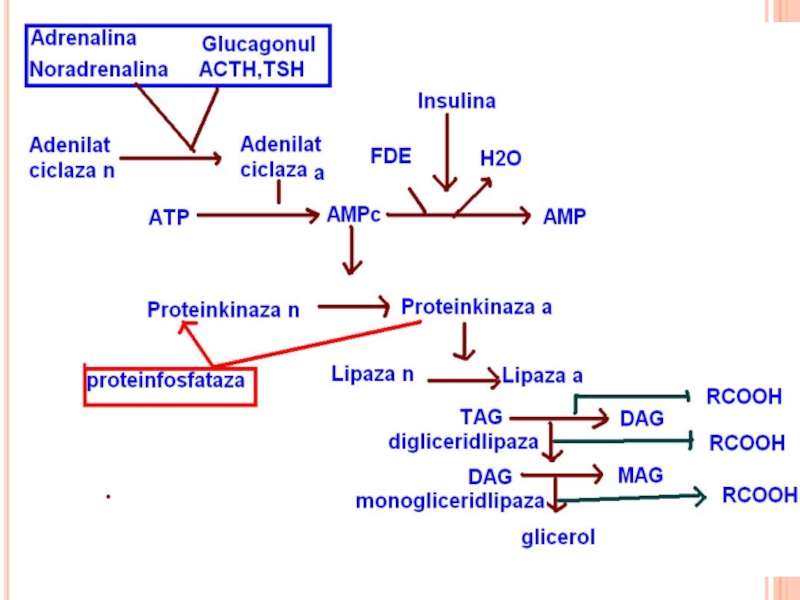
Слайд 163TRIGLICERIDLIPAZA
cunoscută ca “lipaza hormonsensibilă”.
Enzima este convertibilă prin fosforilare –defosforilare.
Forma fosforilată
A: catecolaminele (adrenalina, noradrenalina), glucagonul, ACTH, TSH.
I: insulina, prostaglandina E, acidul nicotinic, AG liberi.

Слайд 164SOARTA AG ŞI GLICEROLULUI:
AG sunt transportaţi spre ţesuturi de albumina serică,
se supun β oxidării - acetil-CoA (pentru a obţine ATP sau poate fi utilizat la sinteza Col, corpilor cetonici).
se activează şi participă la sinteza TAG, depozitate în ţesutul adipos
difuzează în plasmă şi circulă sub formă de AG liberi (sunt captaţi de ţesuturile periferice: muşchii scheletici, miocard, rinichi, ficat)
Eritrocitele şi creierul nu pot utiliza AG ca sursă de energie

Слайд 165SOARTA GLICEROLULUI:
Glicerolul nu poate fi folosit de ţesutul adipos, lipsit de
Glicerolul:
Sinteza de TAG şi FL
Sinteza glucozei - gluconeogeneză
Oxidează pînă la CO2 şi H2O


Слайд 167OXIDAREA AG SATURAŢI CU NUMĂR PAR DE ATOMI DE CARBON
β

Слайд 168OXIDAREA AG SATURAŢI CU NUMĂR PAR DE ATOMI DE CARBON
Procesul e
Excepţie:creierul şi eritrocitele (ţesuturi glucozo dependente)

Слайд 169ΒETA OXIDAREA AG
3 etape:
Activarea AG (citoplasmă)
Transferul lui Acil CoA în
β oxidarea propriu zisă (mitocondrii)
 Transferul lui Acil CoA în mitocondriiβ oxidarea propriu zisă (mitocondrii)")
Слайд 170ACTIVAREA AG:
R-COOH + HS-CoA + ATP ? R-CO~SCoA + AMP +
H2O ↓
2 Pi
E- acil Co A sintetaza
Activatori: K; Mg
Inhibitori: Na ; Li

Слайд 171ACTIVAREA AG:
R-COOH + ATP ? R-COO-AMP + PPi
PPi ? 2 Pi
R-COO-AMP + HS-CoA ? R-CO~SCoA + AMP
acil-CoA
Sumar:
R-COOH + ATP + HS-CoA ? R-CO~SCoA + AMP + PPi
H2O ↓
2 Pi

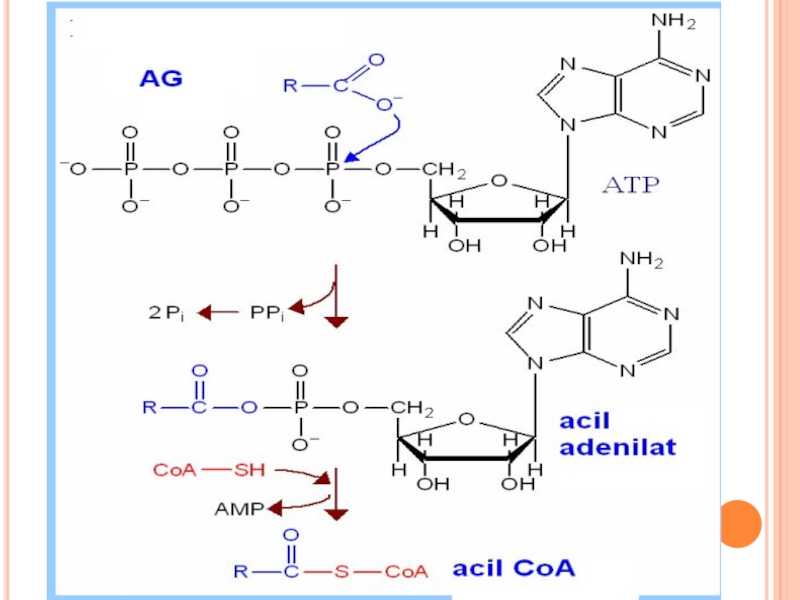
Слайд 173TRANSFERUL LUI ACIL COA ÎN MITOCONDRII
Acil CoA nu poate penetra membrana
Este transportat cu ajutorul carnitinei (β-hidroxi-γ-trimetilaminobutirat- vitamina Bt), ce se formează din Lyz şi Met activă cu participarea vitaminei C, B6, NAD



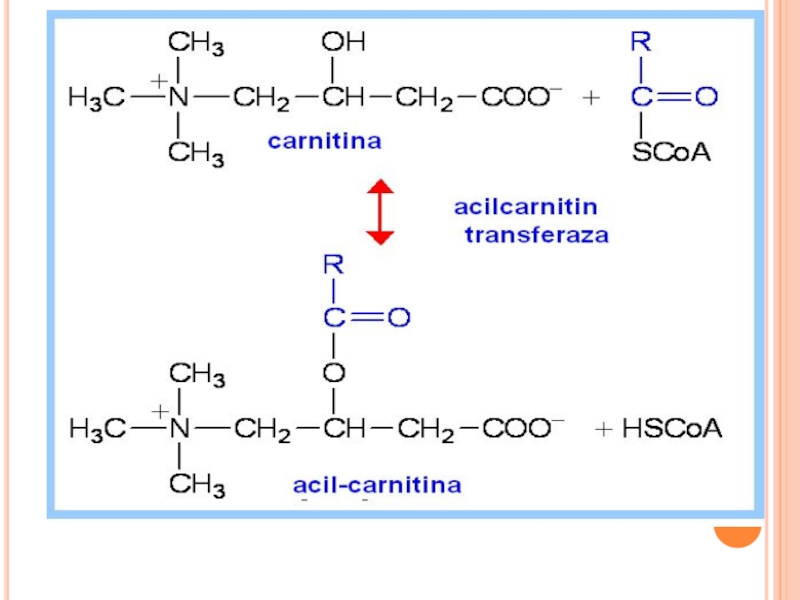
Слайд 177REGLAREA
Malonil CoA înhibă ACT I---deci palmitatul format în citozol (biosinteza lipidelor)
Creşterea raportului acetil CoA/CoA duce la inhibarea reacţiei de activare a AG
 nu poate fi transferat")
Слайд 178DEFICIENŢA DE CARNITINĂ
1. congenitale –
a.deficit de ACT I (afectează ficatul
b. deficitul de ACT II – afectează muşchiul cardiac şi scheletic – cardiomiopatie, slăbiciune musculară (efort fizic)
2. dobîndite: boli hepatice, persoanele supuse hemodializei, malnutriţie, dietă strict vegetariană.

Слайд 179
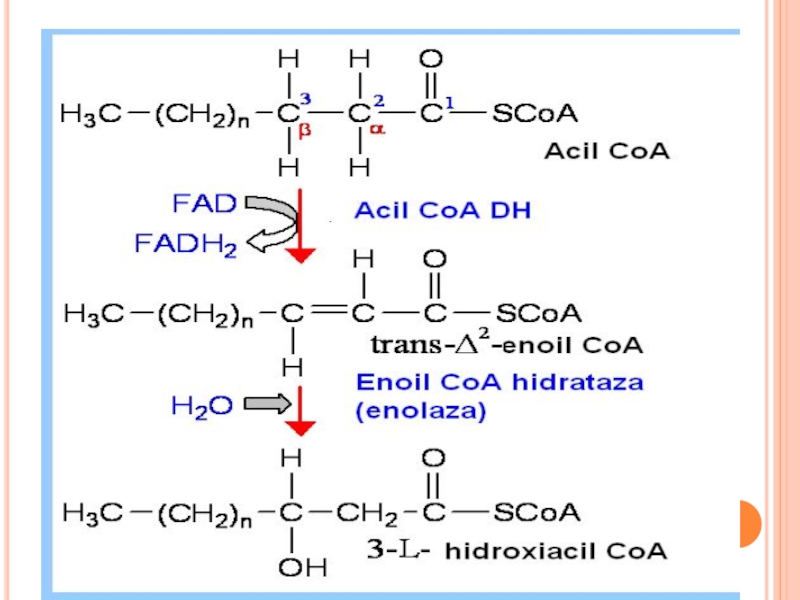
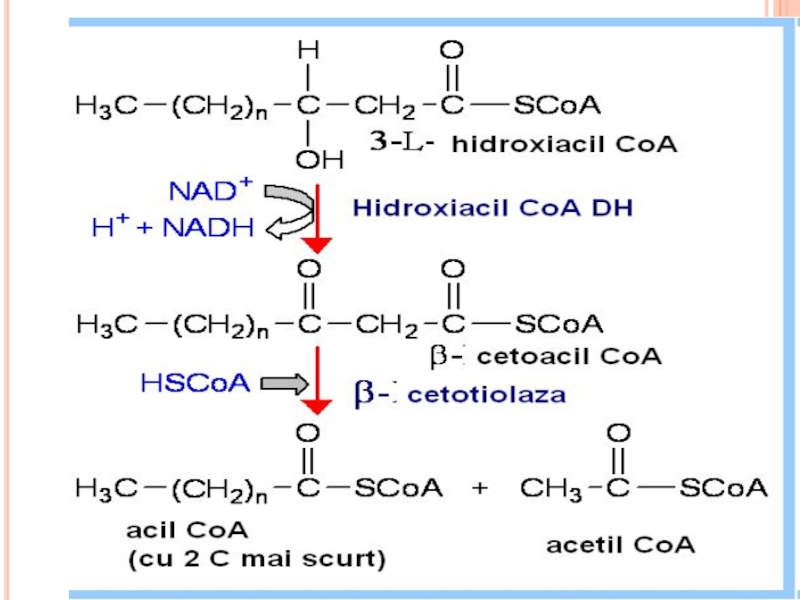
BETA OXIDAREA PROPRIU ZISĂ
Localizat în MC (matrixul MC)
repetarea a 4
Dehidrogenarea lui acil Co A (FAD)
hidratare
a doua dehidrogenare (NAD)
reacţie tiolazică (clivare)
În rezultat - se formează acetil CoA şi acil CoA cu doi atomi de carbon mai puţin
repetarea a 4 reacţii:Dehidrogenarea lui acil")
Слайд 182BILANŢUL ENERGETIC
Stoichiometria unui ciclu de oxidare:
CH3- (CH2)n-CH2 – CH2-COSCoA +FAD+
H2O+NAD+HSCoA
Acil CoA (Cn-2) +FADH2+NADH+H+ Acetil CoA
Stoichiometria oxidării a. palmitic (C16):
n/2 -1 – numărul de cicluri pînă la oxidarea completă
n– numărul atomilor de C
n-CH2 – CH2-COSCoA +FAD+H2O+NAD+HSCoA")
Слайд 183STOICHIOMETRIA OXIDĂRII A. PALMITIC
16/2 -1 = 7 cicluri
7FADH2--------– 7 X
7NADH+H ----- 7X3=21 ATP
8 CH3COSCoA--- 8X12= 96 ATP
Sumar: 131 mol de ATP
131-1=130 ATP
Deoarece 2 legături macroergice sunt irosite la activarea acidului – beneficiul net este de 129

Слайд 184BILANŢUL ENERGETIC AL OXIDĂRII COMPLETE A ACIDULUI PALMITIC
Beneficiul net al arderii

Слайд 185OXIDAREA AG NESATURAŢI
β-oxidarea AG nesaturaţi se desfăşoară normal pînă în vecinătatea
După trei cicluri normale de β-oxidare se ajunge la un cis – Δ3 – enoil – CoA.
Sub acţiunea cis Δ3 - trans ∆2 enoil Co A izomerazei legătura dublă din cis – Δ3 trece în trans-∆ 2 – se formează trans – Δ2 – enoil – CoA, intermediar normal al β-oxidării.
Exemplu: oxidarea acidului oleic (C18:1Δ9)
CH3-(CH2)7-CH=CH-(CH2)7-COOH
După")
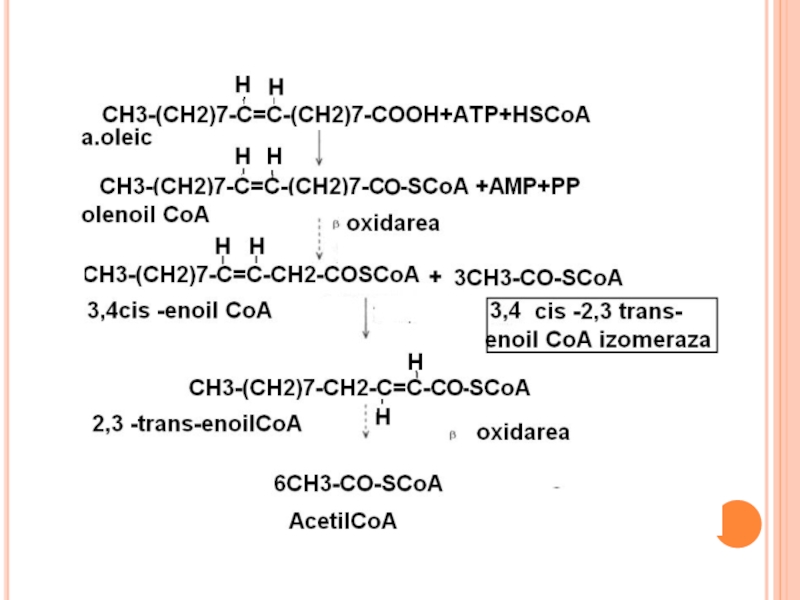
Слайд 187
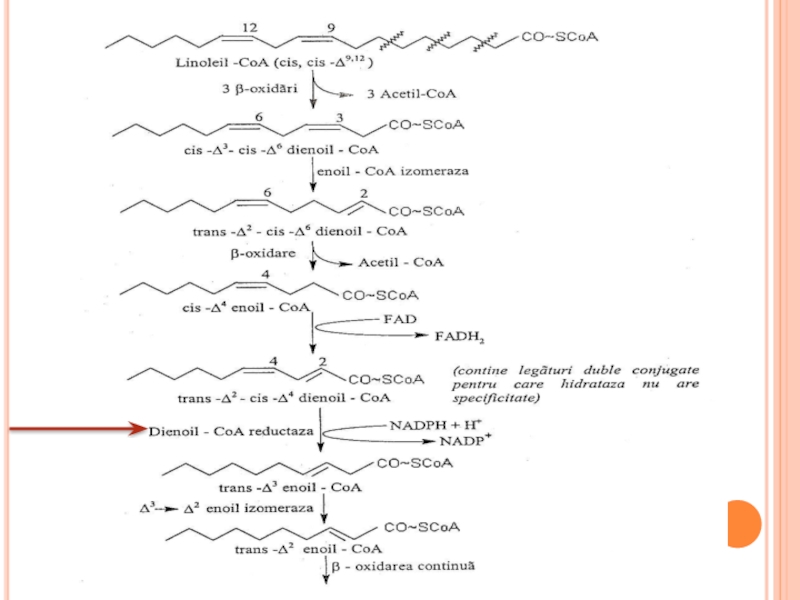
Pentru AG polienici e necesară şi o altă enzimă – dienoil
Produsul sub acţiunea izomerazei trece în trans 2 enoil CoA.
")
Слайд 189OXIDAREA AG CU NUMĂR IMPAR DE ATOMI DE C
Se oxidează în

Слайд 190OXIDAREA AG CU NUMĂR IMPAR DE ATOMI DE C
E- propionil CoA
Co- vitamina H (biotina)
E- Metilmalonil-mutaza
Co- vitamina B12
Lipsa acestei E – acidemie metilmalonică (în sînge şi urină apare acidul metilmalonic, micşorînd pH sîngelui (administrat vitamina B12)
E-")
Слайд 191BILANȚUL ENERGETIC PENTRU AG CU NUMĂR IMPAR
Succinil –CoA Succinat Fumarat
Malat
H2O
MC
CITOZOL
NADP+ NADPH+H+
Malat Piruvat
CO2
MC
Piruvat Acetil Co A
Krebs
NAD NADH+H
CO2

Слайд 192OXIDAREA AG ÎN PEROXISOMI
Caracteristic AG C20-C26 ----pînă la C8
Diferă de oxidarea

Слайд 193Numărul peroxisomilor creşte în diabet, inaniţie, la administrarea unor medicamente (aspirina,
Absenţa peroxisomilor- sindromul Zellweger: creşterea AG cu catena lungă şi deces în primele luni de viaţă
Oxidarea AG în peroxisomi
Absenţa peroxisomilor- sindromul")
Слайд 194ALTE TIPURI DE OXIDĂRI
α-oxidare – la acizii graşi ramificaţi
Reacţia este catalizată
Rezultă hidroxiacizi care pot fi din nou oxidaţi şi decarboxilaţi, formând acizi graşi cu 1 at.C mai puţin decât originalul;
ω-oxidare – oxidarea at.C cel mai depărtat de gruparea carboxil
În microzomi;
În prezenţa monoxigenazei, NADPH şi citocromul P450;
Se formează acizi dicarboxilici.


Слайд 196REGLAREA METABOLISMULUI LIPIDIC
Se efectuiază în mai multe aspecte:
Factorii Enzimatici:
a. Modificarea
b.Modificarea V sintezei şi degradării E
Factorii de mediu: stresul emoţional negativ, inaniţia, hipotermia, efortul muscular cauzează pierderea din greutate a organismului.

Слайд 197Factorii hormonali:
Catecolaminele, Glucagonul stimulează lipoliza prin AMPc, activează lipaza hormono-dependentă.
hormonul de creştere la fel, dar cu o fază mai tardivă (stimulează sinteza adenilat ciclazei de novo).
Cortizolul – acţiune catabolică asupra ţesutului adipos periferic (lipoliza)
Insulina posedă efect contrar, activează fosfodiesteraza – inhibă lipoliza şi stimulează lipogeneza.

Слайд 198INSULINA
activează biosinteza AG în ficat
Activează glicoliza – DHAP—Glicerol 3P
Activează PDH –Acetil
Intensifică formarea citratului şi transformarea sa în Acetil CoA (activarea citrat liazei)
Activarea acetil Co A carboxilazei ----malonil CoA

Слайд 199METABOLISMUL LIPIDIC ÎN PERIOADELE POSTPRANDIALE
Mărit raportul Insulină/glucagon
DIGESTIA
ABSORBŢIA ------ chilomicroni (LPL) –AG
În ficat:
A. Sinteza AG (din glucide sau alcool)
B. Sinteza AB, Col, FL, TAG
C. Sinteza LP (VLDL; HDL)
 –AG liberi, captaţi de ţesut")
Слайд 200METABOLISMUL LIPIDIC ÎN PERIOADELE POSTPRANDIALE
VLDL sub acţiunea LPL ---- AG--- sunt

Слайд 201METABOLISMUL LIPIDIC ÎN PERIOADELE INTERALIMENTARE
Scăzut raportul Insulină/glucagon
TAG din ţesutul adipos sunt
AG se supun beta oxidării------ Acetil Co A—ciclul Krebs---ATP
În ficat Acetil CoA --- sinteza corpilor cetonici (în inaniţie)

Слайд 202SINTEZA CORPILOR CETONICI
(CETOGENEZA)
acetoacetatul,
β-hidroxibutiratul
acetona
Sinteza lor are loc în ficat (în
Ca substrat – Acetil CoA (βoxidarea AG, ce provin din lipoliza adipocitară, degradarea AA cetoformatori)
Rol: sursă de energie în perioadele de inaniţie (muşchiul cardiac, stratul cortical al rinichilor)
acetoacetatul, β-hidroxibutiratul acetonaSinteza lor are loc în ficat (în mitocondriile hepatocitelor ),Ca")
Слайд 203SINTEZA CORPILOR CETONICI
(CETOGENEZA)
Are loc când concentaţia AG în sânge este mare
Principala cale de metabolizare a acetil CoA – includerea în ciclul Krebs (în condiţiile în care scindarea lipidelor şi a glucidelor este echilibrată) - “lipidele ard în flacăra glucidelor”
În lipsa glucidelor; inaniţie, diabet - OA se utilizează pentru generarea Gl.
În lipsa OA, Acetil Co A recurge la formarea corpilor cetonici: acetoacetatul, β-hidroxibutiratul şi acetona
Are loc când concentaţia AG în sânge este mare (foame, inaniţie, dietă")
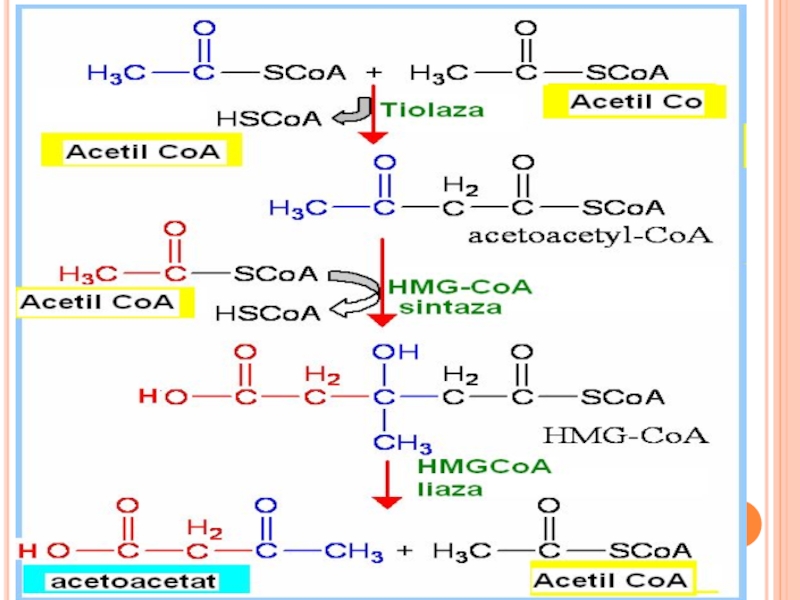

Слайд 206UTILIZAREA CORPILOR CETONICI
Sunt substrate energetice preferate pentru inimă, muşchi scheletici, cortex
Acetoacetatul şi β-hidroxibutiratul servesc ca substrate majore pentru sinteza lipidelor cerebrale neonatale;
β-hidroxibutiratul reprezintă o cale de export a echivalenţilor reducători din ficat (25% din NADH obţinut din b-oxidarea acizilor graşi este folosit pentru sinteza acestui corp cetonic).

Слайд 207ACTIVAREA ACETOACETATULUI
Transportul corpilor cetonici prin sânge se face sub formă liberă,
Acidul acetoacetic din ficat trece în sânge de unde este preluat de ţesuturile extrahepatice şi activat:
1. Calea tioforazică:
Reacţie de schimb cu succinil CoA


Слайд 209ACTIVAREA ACETOACETATULUI
2. Calea tiokinazică:
- Acetoacetatul – 2 mol de acetil

Слайд 210ACTIVAREA BETA-HIDROXIBUTIRATULUI
Cea mai importantă cale este transformarea sa în acetoacetat, urmată
Poate avea loc direct în ţesuturile extrahepatice, similar cu activarea directă a acetoacetatului, cu formarea b-hidroxi-butiril CoA, care sub acţiunea unei dehidrogenaze specifice se transformă în acetoacetil CoA.


Слайд 212Prin oxidarea unui mol de acetoacetat se obţin 23 moli ATP
BILANȚUL ENERGETIC AL OXIDĂRII CORPILOR CETONICI

Слайд 213
CORPII CETONICI
FICAT
AG
SÂNGE
CO2 +ATP acetil-CoA acetoacetat β-hidroxibutirat
acetoacetil-CoA
ciclul
Krebs
Oxidarea AG
succinil-CoA
succinat
MUŞCHI, CREIER
ŞI RINICHI
Beta-cetoacil-CoA
transferaza

Слайд 214CETONEMIE, CETONURIE
Cetonemie- mărirea c% de corpi cetonici în sînge
Cetonurie – apariţia
Se observă în:
Diete bogate în lipide, sărace în glucide;
inaniţie,
Diabet zaharat insulinodependent,
dereglări gastrointestinale la copii sau gravide
glucozurie renală

Слайд 215CAUZELE CETONEMIEI
Mărirea concentrației lui Acetil - Co A (lipoliza activată).
2. Micșorarea
.2. Micșorarea OA (se include ăn gluconeogeneză)")
Слайд 216CETOACIDOZA DIABETICĂ
CC depăşesc 20 mmol/l (normă – 0,03–0,2 mmol/l)
-deficitul de insulină
Compensator – se măreşte secreţia hormonilor de contrareglare (STH; adrenalinei, cortizolului, glucagonului)
-deficitul de insulină – blochează transportul Gl")
Слайд 217CETOACIDOZA DIABETICĂ
- Sub influienţa acestora se activează: glicogenoliza, gluconeogeneza – hiperglicemie
pentru asigurarea celulei cu energie sub influienţa H contrareglare se activează lipoliza, cu eliberarea de AGL, care la nivelul ficatului sunt transformaţi în corpi cetonici.

Слайд 218CETOGENEZA ÎN DIABET
Raportul glucagon/insulină creşte;
Creşte şi concentraţia hormonilor de stres: adrenalină,
Creşte glicemia ca urmare a GNG şi glicogenolizei;
Creşte lipoliza şi proteoliza
Reorientarea spre utilizarea corpilor cetonici

Слайд 219CETOACIDOZA DIABETICĂ
Eliminarea hidroxibutiratului şi acetoacetatului din organism (fiind anioni la excreţie)
Pierderea H2O – dehidratarea organismului
 conduce la pierderea de")
Слайд 220CETOGENEZA ÎN INANIŢIE
Glicemia este scăzută, corpii cetonici sunt utilizaţi iniţial de
După un post de 3 zile 1/3 din energia creierului provine din oxidarea corpilor cetonici;
După o săptămână de post, corpii cetonici devin sursa majoritară de energie a creierului, necesarul de glucoză fiind minim (se cruţă proteinele musculare).


Слайд 222OBIECTIVELE
Metabolismul colesterolului. Biosinteza colesterolului – substratele, etapele, reacţiile parţiale ale I
Căile de utilizare şi eliminare ale colesterolului.
Biosinteza fosfogliceridelor: substratele, reacţiile parţiale ale I şi a II căi;
Biosinteza sfingolipidelor: precursorii, reacţiile principale, enzimele, reglarea.
Oxidarea fosfo-, sfingo- şi glicolipidelor.
Metabolismul eicosanoizilor. Căile ciclooxigenazică şi lipooxigenazică ale biosintezei lor. Inactivarea.
Rolul eicosanoizilor în procesele inflamatorii, reacţiile alergice, dereglările fluidităţii sanguine

Слайд 223SINTEZA COLESTEROLULUI
Se sintetizează din Acetil-CoA
Necesită 18 moli de Acetil-CoA, 16 NADPH+H
Localizarea:
Ficat (80%), dar are loc şi în intestin, suprarenale, tegumente
Începe în citoplasmă (continuă în RE şi peroxisomi)

Слайд 224SINTEZA COLESTEROLULUI
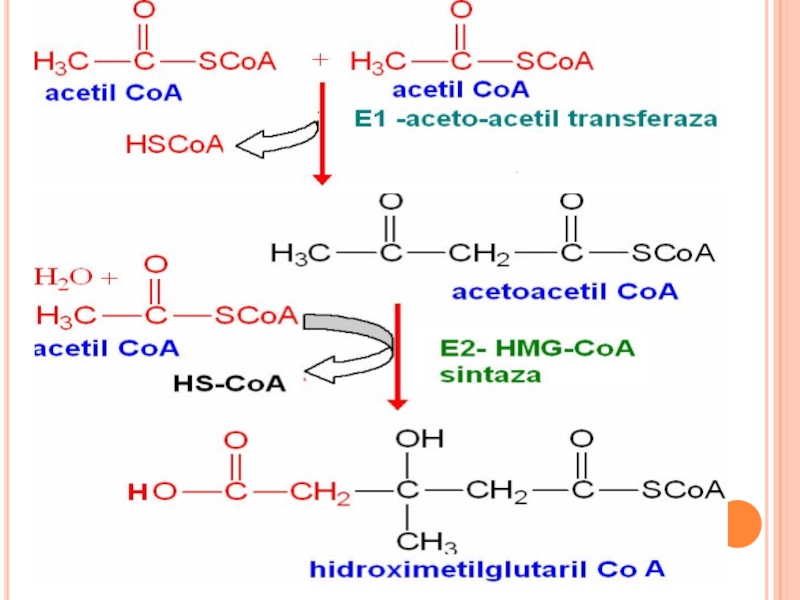
Are loc în 3 etape:
Sinteza acidului mevalonic
mevalonatul prin mai multe
Scualenul se supune ciclizării – lanosterol- Col


Слайд 227REGLAREA HMG-COA REDUCTAZA
la nivelul sintezei (v de transcriere a genelor)
expresia
Ea este asociată cu proteina care activează clivarea ei – SCAP.
Cînd nivelul sterolilor scad – complexul SREBP-SCAP părăseşte RE şi la nivelul aparatului Golgi, proteazele acţionează asupra SREBP şi formează un fragment solubil care pătrunde în nucleu şi funcţionează ca factor de transcripţie ---- se intensifică sinteza HMG-reductazei.
Cînd sterolii sunt în c% mari --- are loc reţinerea complexului SCAP-SREPB în RE şi nu are loc activarea SREBP --- inhibarea sintezei de Col.
expresia genei HMG-reductazei este controlată")
Слайд 228REGLAREA HMG-COA REDUCTAZA
la nivelul sintezei (v de transcriere a genelor)
la
3. Retroinhibiţie - enzima este inhibată de mevalonat şi de colesterol
4. Reglare covalentă- forma activă este forma defosforilată
5. Reglarea hormonală - insulina şi hormonii tiroidieni activeză HMG-CoA reductaza, iar glucagonul are efect opus
la nivelul modificării v de")
Слайд 229REGLAREA
Reacţia – limită a biosintezei e formarea acidului mevalonic, catalizată de
HMG-CoA reductaza – localizată la nivelul RE
Structura: 887 AA cu 2 domenii structurale: unul hidrofob (membranar) şi altul citoplasmatic (posedă activitate catalitică).

Слайд 230REGLAREA HMG-COA REDUCTAZA
la nivelul sintezei (v de transcriere a genelor)
la
3. Retroinhibiţie - enzima este inhibată de mevalonat şi de colesterol
4. Reglare covalentă- forma activă este forma defosforilată
5. Reglarea hormonală - insulina şi hormonii tiroidieni activeză HMG-CoA reductaza, iar glucagonul are efect opus
la nivelul modificării v de")
Слайд 231
Conţinutul Col depinde de regimul alimentar. Acest colesterol alimentar inhibă sinteza
Inhibarea prin medicamente: statinele – inhibitori reversibili, competitivi ai HMG reductazei.

Слайд 232CĂILE DE TRANSFORMARE ALE COLESTEROLULUI
1. În tegumente ---colecalciferol
2. În suprarenale- sinteza
3.În glande sexuale- sinteza hormonilor sexuali
4. În ficat – sinteza AB primari
5. În ficat şi intestin – LP plasmatice

Слайд 233CATABOLISMUL ŞI ELIMINAREA COL
1. Calea biliară:
Col biliar---intestin --reducere la coprostanol sau
AB neabsorbiţi (0,25g/zi)----fecale
2. Celule epiteliale descuamate----fecale
3.Secreţia sebacee la nivelul tegumentelor
4.Eliminare urinară a metaboliţilor hormonilor steroizi şi a vitaminelor D
----fecale2. Celule")
Слайд 234LITIAZA BILIARĂ
Colesterolul (care este hidrofob) este dizolvat în bilă prin formarea
O modificare minoră a acestui raport determină cristalizarea colesterolului în jurul nucleului format din proteine şi bilirubină cu generare de calculi biliari - litiază biliară
 este dizolvat în bilă prin formarea de micelii cu fosfolipide")
Слайд 235SINTEZA GLICEROFOSFOLIPIDELOR
LOCALIZAREA:
Are loc intensiv în ficat; peretele intestinului, testicule, ovare, glanda
Sunt sintetizate în reticulul endoplasmatic neted, transportate în aparatul Golgi, iar ulterior către membrane
2 căî de sinteză:
sinteză PC și PE din colina sau etanolamina preexistentă
Sinteza PI și cardiolipinei ce utilizează ca intermediar comun acidul fosfatidic
Particularitatea biosintezei FL este participarea precursorilor în forme active de derivaţi ai citidin fosfatului (CDP) ca CDP-colina, CDP-etanolamina, CDP-diglicerid.

Слайд 236SINTEZA FOSFATIDILCOLINEI (PC) ȘI FOSFATIDILETANOLAMINEI(PE)
Colina si etanolamina se obtin din alimentatie
Reacțiile cuprind:
fosforilarea colinei/etanolaminei
conversia la forma activata: CDP-colina/CDP-etanolamina
colin-fosfatul sau etanol-fosfatul sunt transferate din nucleotid (parasind CMP) pe o molecula de DAG
 ȘI FOSFATIDILETANOLAMINEI(PE)Colina si etanolamina se obtin din alimentatie sau din catabolizarea FLReacțiile")

Слайд 238IMPORTANȚA REUTILIZĂRII COLINEI
COLINA în organism:
Sintetizată de novo (insuficientă)
Din alimente - necesarul
La femei – 425 mg
La bărbați – 550 mg
Deasemenea ea este utilizată la sinteza acetilcolinei
Din alimente - necesarul zilnic:La femei – 425")
(Fosfoetanolamină)(CDP-etanolamină)(Fosfatidiletanolamină)")
FOSFATIDILETANOLAMINA SERINA ETANOLAMINĂ")

Слайд 242
Ficatul este un organ specific de sinteză a fosfolipidelor plasmei.
În lipsa
În consecinţă, are loc depozitarea lipidelor în ficat - provoace distrofia grasă a ficatului
Orice substanţă donatoare de CH3 pentru sinteza cholinei este denumită lipotropă.
O serie de substanţe, medicamente poartă denumirea de substanţe lipotrofe.
SUBSTANȚELE LIPOTROPE





Слайд 247FOSFOLIPAZELE
A1 – este prezentă în multe dintre ţesuturile mamiferilor
A2- este prezentă
E prezentă în veninul de şarpe şi cel de albine
Prin acţiune asupra fosfatidilinozitolului – eliberează a. arahidonic (precursorul prostaglandinelor)
Fosfolipaza C se afşă în lizozomii hepatici (produce mesagerul secund)

Слайд 248SINTEZA SFINGOLIPIDELOR
Se formează din palmitoil CoA şi Ser
Sfingozina liberă se formează
Sinteza are loc pe suprafaţa citozolică a membranelor reticulului endoplasmatic

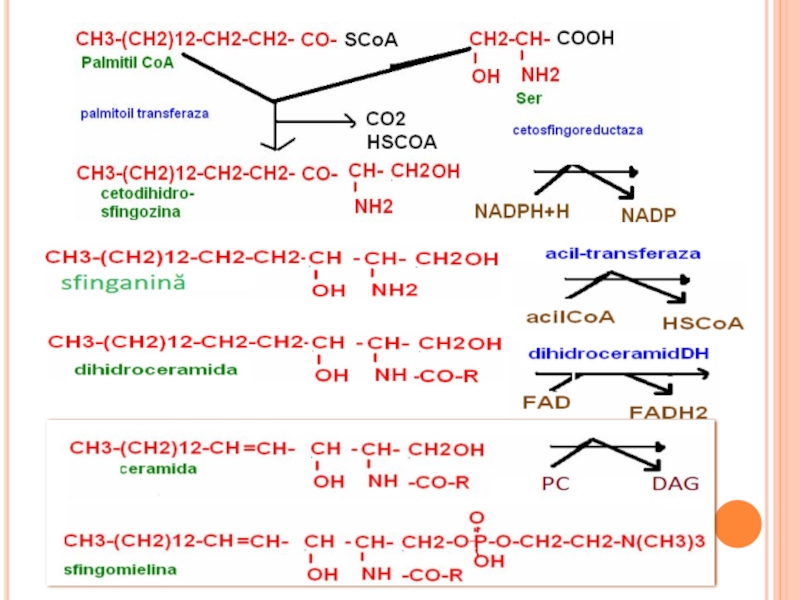
Слайд 250CATABOLISMUL SFINGOMIELINELOR
Sfingomielinaza (lizozomală): ceramid+fosforilcolină
Ceramidaza – degradează legătura N acilică
: ceramid+fosforilcolinăCeramidaza – degradează legătura N acilică")
Слайд 251CATABOLISMUL SFINGOMIELINELOR
Ceramida poate fi implicată în răspunsul la stres
Sfingozina inhibă proteinkinaza

Слайд 252SINTEZA GLICOLIPIDELOR
Sinteza cerebrozidelor:
La sinteză participă:UDP-Gl; UDP-Gal
Ceramidă +UDP-Gal (UDP-Gl)----Calactocerebrozidă (Glucocerebrozidă)
Ceramidă +(UDP-monozaharide)n-----Globozid
----Calactocerebrozidă (Glucocerebrozidă)Ceramidă +(UDP-monozaharide)n-----Globozid")

Слайд 254GRUPARI SULFAT: DE LA TRASNPORTORUL DE SULFAT 3'-FOSFOADENOZIN-5'-FOSFOSULFAT (PAPS): GALACTOCEREBROZID-3-SULFATUL=SULFATIDA CEA
BIOSINTEZA SULFATIDELOR
: GALACTOCEREBROZID-3-SULFATUL=SULFATIDA CEA MAI IMPORTANTA DIN CREIER")
Слайд 255SINTEZA GANGLIOZIDELOR
La sinteză participă UDP-N-acetil galactozamină
Globozid +CMP_NANA-----Gangliozid +CMP


Слайд 257CATABOLISMUL GLICOLIPIDELOR
Cerebrozidele sunt degradate de hexozidaze care îndepărtează resturile de glicozil
fiecare tip de legătură glicozidică este scindată de o glicozidază spesifică.
Carenţa acestor E (localizate în lizozomi) - lipidoze
 fiecare tip")
Слайд 258LIPIDOZE TISULARE
Boli ereditare, caracterizate prin acumularea de sfingolipide în creier şi

Слайд 259LIPIDOZE TISULARE
1. Neimann-Pick - deficienţa de sfingomielinază
sfingomielina SM- se acumulează în
Hepato- şi splenomegalie, tulburări digestive, întîrziere psihomotorie, convulsii
Decedează în vârsta fragedă

Слайд 260LIPIDOZE TISULARE
2. afecţiunea Tay-Sachs – lipsa hexozaminidazei (hidrolază ce scindează N
Acumularea de GM2 în creier – întîrzieri psihomotorii, creşterea în volum şi masă a creierului, atrofie musculară, paralizie, orbire.
pe retina ochiului – apar pete de culoare roşie-vişinie – gangliozid GM2
Acumularea de")
Слайд 261EICOSANOIZII
Sunt substanţe biologic active de natură lipidică, derivate din a.graşi polinesaturaţi

Слайд 262EICOSANOIZII
Deosebim:
Prostaglandinele (clasice- PG şi endoperoxizii prostaglandinici (PGG2 şi PGH2)
Prostaciclina (PGI2)
Tromboxanii (TX)
Leucotrienele
Prostaciclina (PGI2)Tromboxanii (TX)Leucotrienele (LT)")
Слайд 263STRUCTURA CHIMICĂ- PROSTAGLANDINE
Reprezintă AG nesaturaţi cu 20 C
Sunt derivaţi ai acidului
Existenţa unui ciclu pentanic format între C8-C12
PG se deosebesc între ele prin:
1. natura substituienţilor din nucleul pentanic (PGA-PGI)
2. Catenele laterale pot conţine un număr diferit de legături duble (PG1, PG2, PG3)

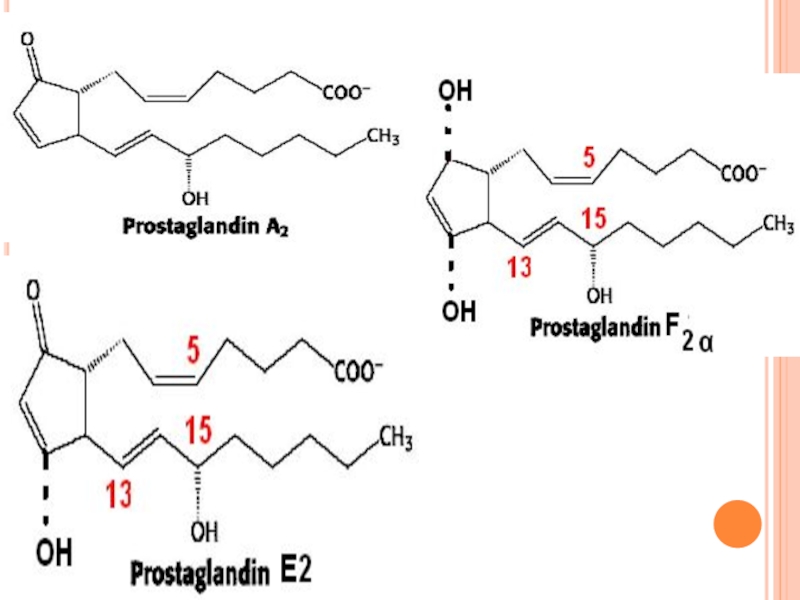
Слайд 265
TX conţin în structura lor ciclul piranic (TXA2 şi TXB2)
TXA2 este
TXA2 este sintetizat din PGH2 de")
Слайд 266STRUCTURA CHIMICĂ - LT
LT (LTA-LTE) – cuprind în structura lor mai
 – cuprind în structura lor mai multe legături duble, trei")
Слайд 267BIOSINTEZA EICOSANOIZILOR
Are loc la nivelul tuturor ţesuturilor
După sinteză nu se depozitează
Are loc în 2 etape:
Sinteza acidului arahidonic (comună)
Sinteza PG, TH, LT

Слайд 268I: SINTEZA ACIDULUI ARAHIDONIC
Derivat al acidului linoleic
Este prezent la nivelul membranelor
Se eliberează prin activarea fosfolipazei A2

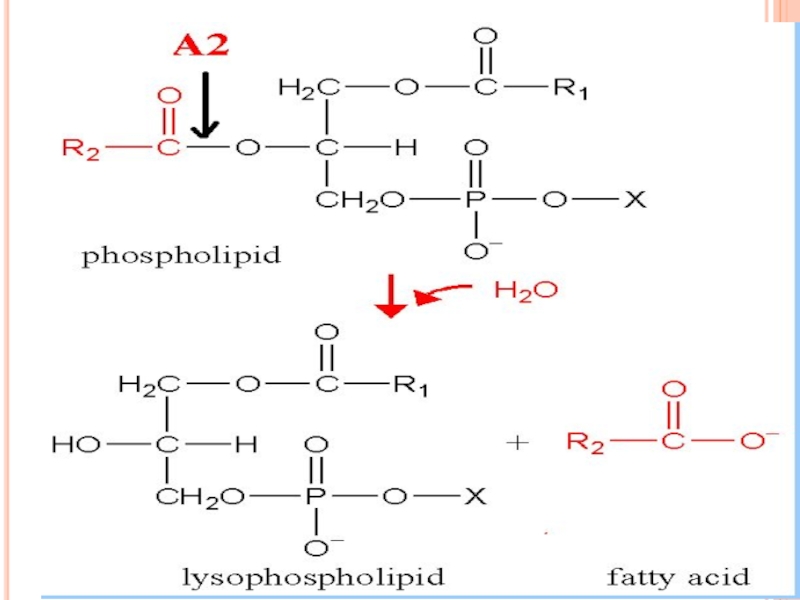
Слайд 270II.SINTEZA PG, TX ŞI LT
2 căi:
Calea ciclooxigenazei conduce la sinteza PG,
Calea lipooxigenazei – la sinteza LT


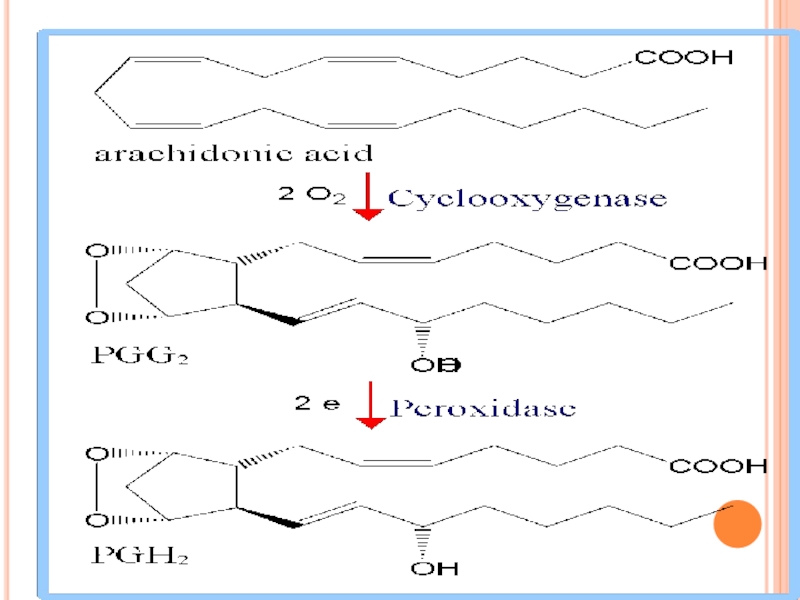
Слайд 272CICLOOXIGENAZA
este o hemoproteină, cu o activitate dublă:
a. dioxigenazică (încorporează O2 în
b. peroxidazică (descompune peroxidul).
b. peroxidazică (descompune peroxidul).")
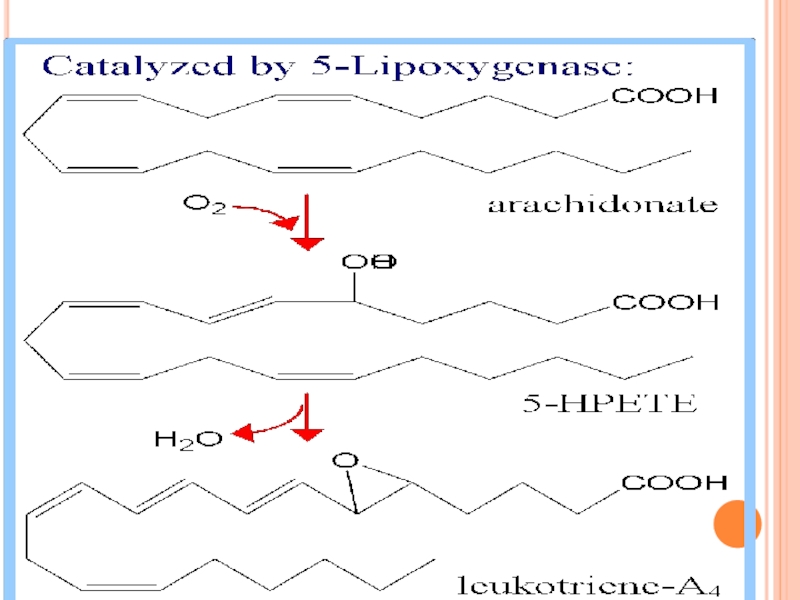
Слайд 275COX-1 – enzimă constitutivă; mucoasa gastrică, plachete, endoteliu vascular, rinichi
COX-2 –
COX-3 – în creier, ţesuturile periferice; este inhibată de paracetamol şi metamizol (analgezice şi antipiretice)
Ciclooxigenaza

Слайд 276INHIBITORII SINTEZEI:
Corticosteroizii – inhibă fosfolipaza A2
Aspirina (medicamente antiinflamatoare nesteroide) - inhibă
Derivaţii imidazolului – inhibă tromboxan sintaza.
 - inhibă ireversibil COX (prin acetilarea")
Слайд 277CATABOLISMUL PG
Viaţă biologică scurtă:
TXA2 are T1/2= 30 minute, şi suferă
Prostaciclina (PGI2) - T ½ - 3 minute la 37 °C şi pH 7,5 şi este convertită printr-o hidroliză nonenzimatică la forma inactivă.
PG clasice sunt eliminate la nivelul plămînului
Principalii derivaţi sunt 15-ceto- iar ai prostaciclinelor 6 ceto-, care se elimină cu urina.

Слайд 278MECANISMELE DE ACŢIUNE
Au acţiune autocrină sau paracrină
Determină variaţii ale nucleotidelor ciclice
PG creşte AMPc în adenohipofiză, plachetele pulmonare şi tiroidă, însă descreşte AMPc în ţesutul adipos.
Tromboxanii blochează producerea de AMPc şi mobilizează Ca2+ intracelular.

Слайд 279ACŢIUNILE BIOLOGICE
Diferă nu numai de natura chimică (PG, LT, TX) dar
Prostaglandinele:
Vasodilataţie puternică
Reducerea sintezei interleukinelor IL-1 şi IL - 2
Diminuarea agregării plachetare şi leucocitare
PGF2 alfa – brohoconstricţie, vasoconstricţie periferică şi contracţia fibrelor musculare netede
 dar şi de cea a")
Слайд 280
Plachetele sanguine (trombocite) conţin tromboxan sintază; se obţine TXA2 - vasocontrictor
Celulele endoteliale ale sistemului vascular conţin prostaciclin sintază; se obţine prostaciclina (PGI2), vasodilatator şi inhibitor al agregării plachetare
Prostaciclina şi tromboxanul alcătuiesc un sistem de control al homeostaziei tonusului vaselor sanguine şi al agregării plachetare
Acţiunile biologice
 conţin tromboxan sintază; se obţine TXA2 - vasocontrictor (contracţia arterelor) şi stimulator")
Слайд 281ACŢIUNILE BIOLOGICE
La nivelul TGI:
PGF2α – acţiune contractilă asupra musculaturii netede
PGE2 – relaxează musculatura circulară, acţiune inhibitorie asupra secreţiei de suc gastric şi absorbţiei intestinale.

Слайд 282ACŢIUNILE BIOLOGICE
Sistemul respirator:
PGE şi PGF – implicate în stări patologice
PGF2α – acţiune contractilă asupra traheei, bronhiilor
PGE2- bronhodilatatoare

Слайд 283ACŢIUNILE BIOLOGICE
SNC: reglarea transmisiei sinaptice.
PGE2- scade c% de GMPc –
PGF2α – creşte c% GMPc – efecte contrare
În inflamaţie:
Au rol tromboxanii, prostaciclina şi endoperoxizii intermediari
Implicati în apariţia şi dezvoltarea inflamaţiilor articulaţiilor (artrita reumatoidă), pielii (psoriazis), ochilor (tratament cu corticosteroizi, care inhibă sinteza prostaglandinelor)

Слайд 284ACŢIUNILE BIOLOGICE
Sistemul de reproducere:
la bărbaţi influenţează spermatogeneza; la femei controlează
PGF2α - acţiune contractilă asupra musculaturii netede a uterului.

Слайд 285ACŢIUNILE BIOLOGICE
Sistem circulator:
Prostaciclina şi TX – controlează tonusul vaselor sanguine
TXA2 – acţiune contractilă – creşte TA. La nivelul plachetelor – stimulează agregarea şi formarea de tromb
Prostaciclina – acţiuni opuse (vasodilataţie, inhibarea agregării plachetare)

Слайд 286ACŢIUNILE BIOLOGICE
LT sunt sintetizate de leucocite şi intervin în:
Modificarea permeabilităţii vasculare
Amplificarea
Amplificarea sintezei interleukinelor
Modificarea agregării leucocitelor
Proliferarea limfocitelor T
Sunt implicate în astmul bronșic

Слайд 287ACŢIUNILE BIOLOGICE
LT (LTB4) –determină acumularea de neutrofile în focarul inflamator.
La
LTC4 - responsabilă de producerea şocului anafilactic.
 –determină acumularea de neutrofile în focarul inflamator. La nivelul plămânului – efecte")

Слайд 289OBIECTIVELE
Metabolismul lipoproteinelor
Cauze, mecanismele dereglării metabolismului lipidelor, manifestările biochimice.
Dislipidemiile:
a) hipolipoproteinemiile familiale –
b) hiperlipoproteinemiile primare şi familiale;
c) hiperlipoproteinemiile secundare (dobândite) – în diabet zaharat, alcoolism, afecţiuni ale glandelor endocrine.
Cauze, mecanismele dereglării metabolismului lipidelor, manifestările biochimice.
Metabolismul vitaminelor liposolubile: sursele alimentare, necesităţile diurne, transformările
A-, hipo- şi hipervitaminozele A, D, E, K – cauze, manifestări metabolice.
 hipolipoproteinemiile familiale – afecţiunea Tangier, α- şi")

Слайд 291LIPOPROTEINELE (LP)
LP- sînt complexe lipoproteice, alcătuite din componente lipidice (TAG, FL,
Componentele proteice sînt denumite apolipoproteine (Apo).
LP- sînt complexe lipoproteice, alcătuite din componente lipidice (TAG, FL, Col, colesteride) şi proteice.")
Слайд 292LIPOPROTEINELE (LP)
Rolul LP
Transportul lipidelor exogene şi endogene
Participă la păstrarea compoziţiei lipidice
Reglează procese metabolice celulare
Rolul Apo:
componente amfipatice a LP
oferă situsuri de recunoaştere pentru R de pe suprafaţa celulelor
sunt activatori sau inhibitori ai E ce participă la metabolismul lor
Rolul LPTransportul lipidelor exogene şi endogeneParticipă la păstrarea compoziţiei lipidice a membranelorReglează procese metabolice")
Слайд 293STRUCTURA LP
miez hidrofob - lipidele nepolare (TAG şi esterii Col)
învelişul
LP cuprind şi cantităţi mici de glucide (sub formă de glicoproteine).
 învelişul hidrofil - lipidele amfipatice")


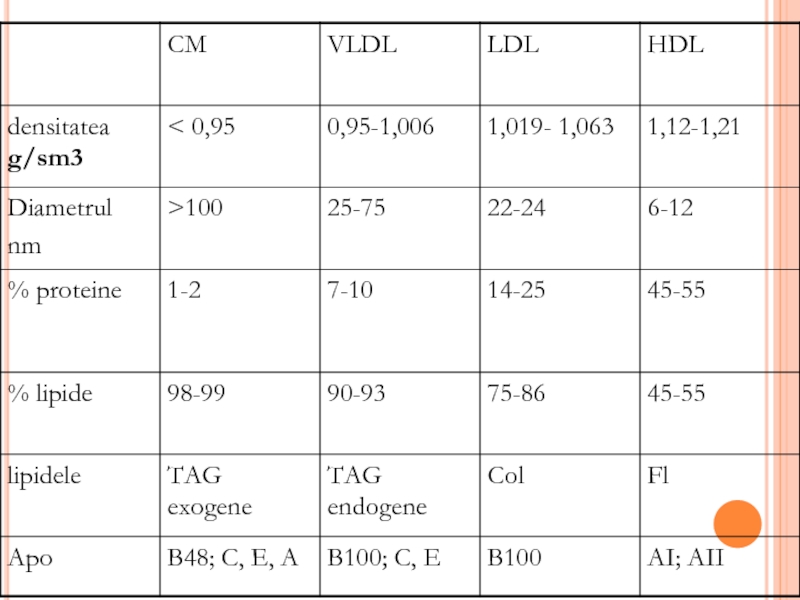
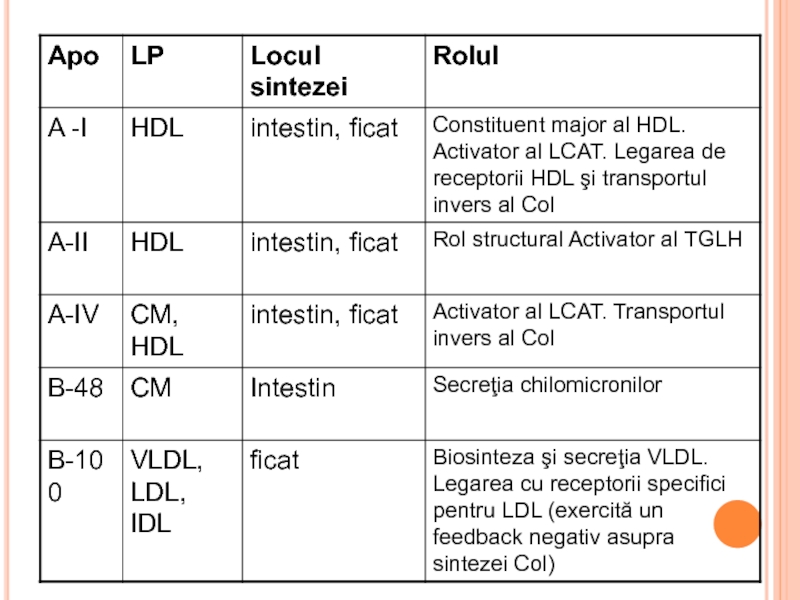
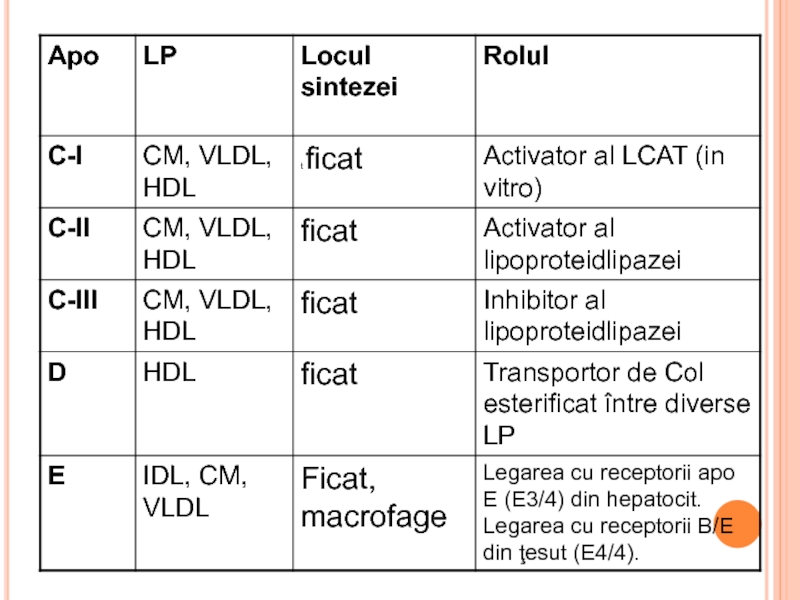
Слайд 299ENZIME IMPLICATE ÎN METABOLISMUL LIPOPROTEINELOR
Lipoproteinlipaza endotelială (LPL-aza):
- localizată în ţesutul
Rol: - hidroliza TAG din CM și VLDL
Activată de apo CII; inhibată de apo CIII
Lecitin-colesterol-acil-transferaza (LCAT) plasmatică- sintetizată în ficat şi înglobată în HDL;
Rol - în îmbogăţirea miezului LP cu CE
lecitină + Col 2-lizolecitină + CE
Lizolecitina hidrofilă părăseşte LP, iar Col liber din periferia LP, esterificându-se devine mai hidrofob şi pătrunde în miezul particulei LP.
Acil-CoA-colesterol acil transferaza (ACAT) intracelulară- catalizează esterificarea Col; rol în depozitare intracelulară a CE
: - localizată în ţesutul adipos, miocard, musculatura scheletică,")
Слайд 300METABOLISMUL CHILOMICRONILOR
Sunt sintetizaţi în celulele mucoasei intestinale şi încorporează lipidele alimentare
Particulile primare cuprind apo-B48 şi apoA.
ROLUL: transportă lipidele exogene din intestin spre țesuturi și ficat
CM sunt prezenţi în plasmă după îngerarea grăsimilor. După 6-7 ore de la îngestia de grăsimi CM dispar din sînge.
.")
Слайд 301METABOLISMUL CHILOMICRONILOR
După sinteză sunt secretaţi în vasele limfatice care drenează intestinul
În plasmă CM primesc Apo C şi E de la HDL

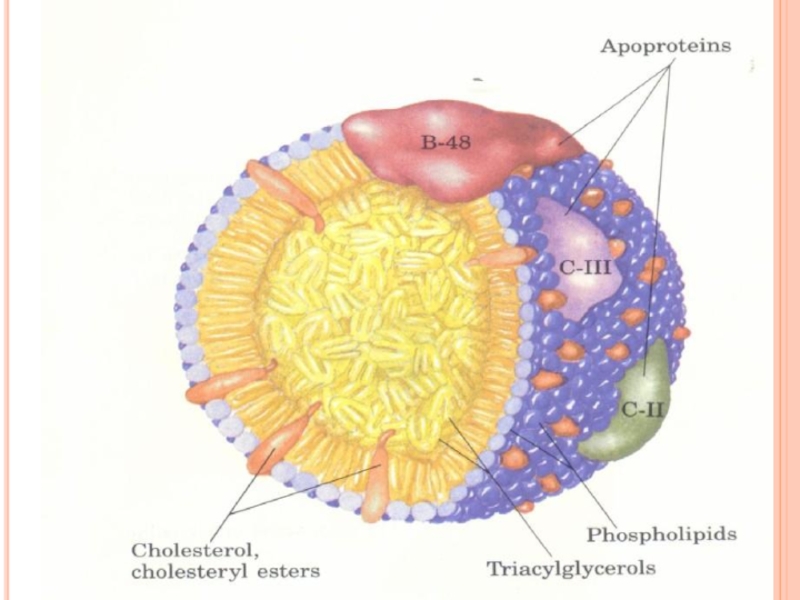
Слайд 303CATABOLISMUL CHILOMICRONILOR
În prima etapă
a. sub acţiunea lipoproteinlipazei (LPL, activată de
Ţesuturile utilizează AG pentru oxidare (muşchi scheletici, miocard), depozitare (ţesut adipos), secreţie de grăsimi (glanda mamară).
b. pe când Col, FL, Apo C sunt transferate pe HDL.
CM devin resturi CM
A două etapă
constă în captarea resturilor de CM de către ficat, facilitată de apo E (interacţionează cu E-receptorii din hepatocit) unde sunt degradate. Pe această cale ajunge la ficat o parte din Col exogen şi cel intestinal
 TAG sunt")
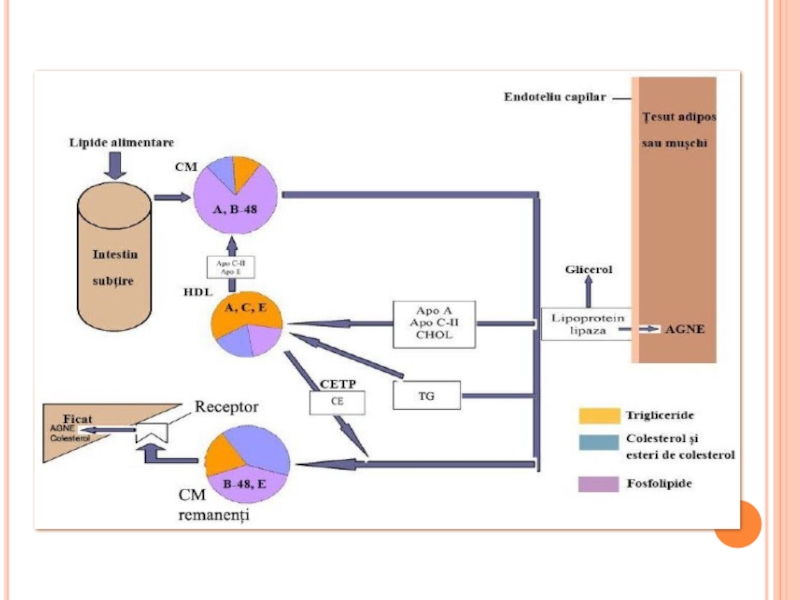
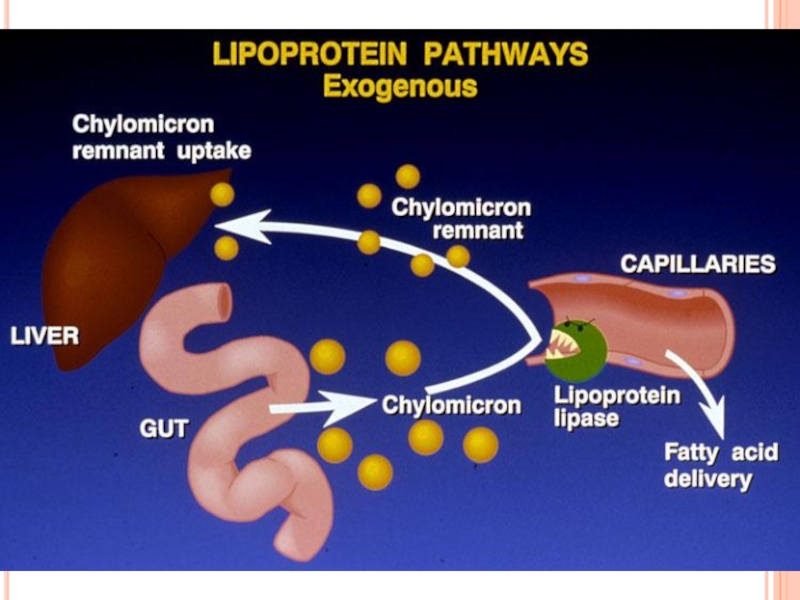

Слайд 307PATOLOGIA
Deficitul înăscut de LPL- hiper-CM-emie
Mărirea c% CM, TAG, depunerea TAG în
Anomalia sintezei de Apo B-48 – imposibilitatea formării CM şi transportului lipidelor exogene
TAG se acumulează în celulele intestinale; este perturbată absorbţia lor; ele fiind eliminate prin masele fecale – steatoree
În sînge: micşorarea lipidelor totale; TAG; Col
Anomalia sintezei de")
Слайд 308PRE-β-LP (VLDL)
sintetizate în ficat din lipidele endogene
au un conţinut ridicat
cuprind apo B-100 ,
Funcţia principală- transportul TAG sintetizate în ficat spre ţesuturile extrahepatice.
în plasmă primesc apo C (apo CII) şi apo E de la HDL.
VLDL sunt prezente în plasmă după îngerare de raţii bogate în glucide
sintetizate în ficat din lipidele endogene au un conţinut ridicat de lipide (90-93%): 55-65%")
Слайд 309CATABOLISMUL VLDL
La nivelul ţesuturilor extrahepatice sub acţiunea LPL-azei TAG din VLDL
Odată cu scăderea TAG are loc:
pierderea de apo C-II care trece pe HDL
Îmbogăţirea cu esteri de Col, furnizat de HDL -
astefl VLDL sunt transformate în IDL.
IDL – o parte sunt catabolizate în ficat prin interacţiunea cu E-R,
iar alta - sunt transformate în LDL.

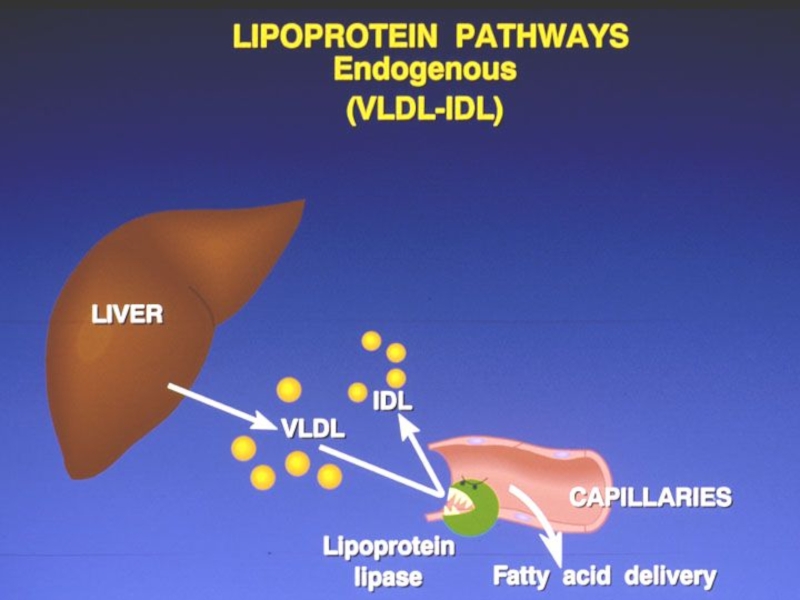
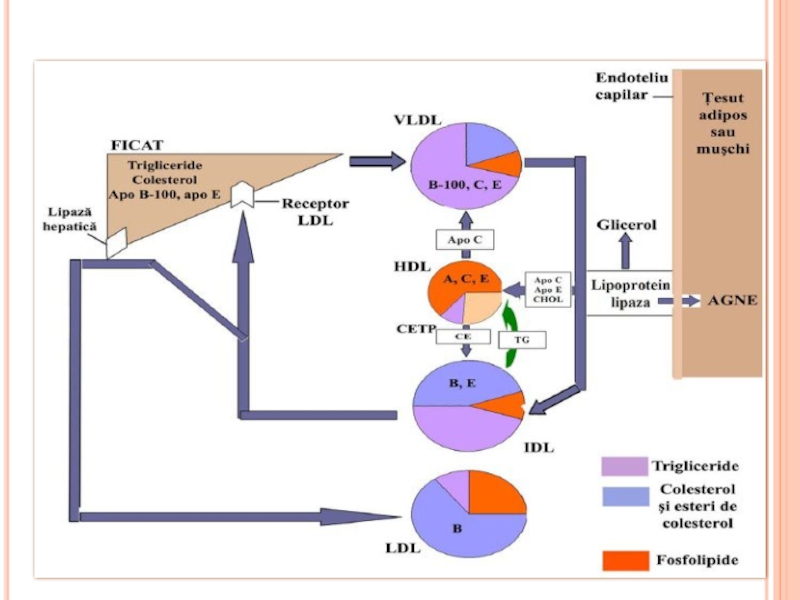
Слайд 312PATOLOGIA
Steatoza (infiltraţia grasă a ficatului) – acumularea grăsimilor în ficat
Cauza: amplificarea
Acţiune protectoare o au factorii lipotropi: Met; proteine bogate în Met; grăsimile nesaturate; vitamina E – ele favorizează exportul TAG hepatice
 – acumularea grăsimilor în ficatCauza: amplificarea sintezei TAG în ficat")
Слайд 313LDL - β-LP
se formează în plasmă din VLDL după îndepărtarea TAG
au un conţinut lipidic de 75-86%, componenta majoră fiind Col: 35-40% - Col esterificat, 3-10% -Col liber; 20-25% - FL şi 8-12% - TAG.
Apo majoritară este B-100.
sunt prezente în sîngele recoltat dimineaţa după un post de 8-10 ore şi cuprinde 70% din Col total plasmatic.
ROLUL: transportă Col de la ficat diverselor ţesuturi periferice.
")
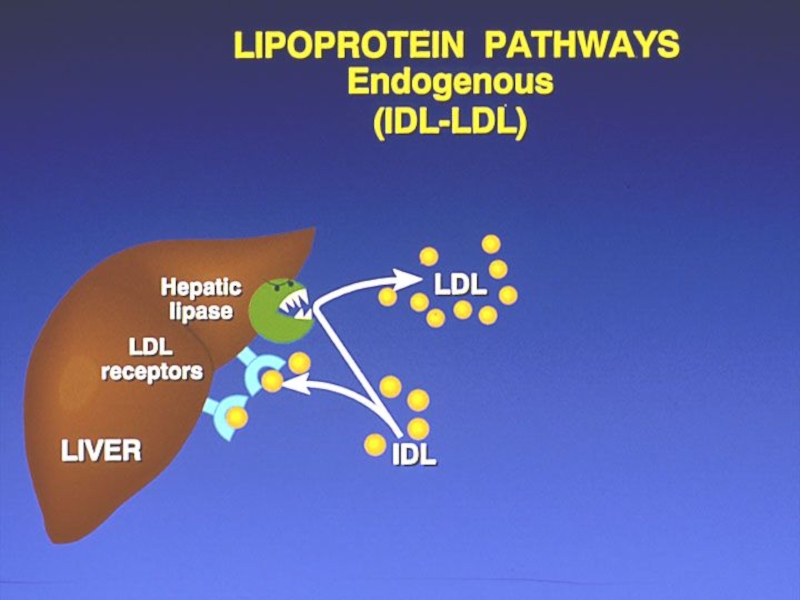
Слайд 315RECEPTORII LDL
1) Receptori LDL clasici- sunt subiectul down-reglării - în funcţie
2) Receptori de tip scavenger- captează LDL alterate (ex. oxidate); nu sunt subiectul down-reglării, ca receptorii LDL clasici; rol în iniţierea aterogenezei şi formarea celulelor spumoase (macrofage care au "înghiţit" prea mult colesterol)
Catabolismul LDL
 Receptori LDL clasici- sunt subiectul down-reglării - în funcţie de cantitatea de colesterol")
Слайд 316CATABOLISMUL LDL
I: În ficat, suprarenale, limfocite, celule musculare netede:
LDL prin intermediul
fixate pe R, LDL sunt translocate în interiorul celulei şi fuzionează cu lizozomii
În lizozomi LDL sub acţiunea E lizozomale sunt hidrolizate în: proteine, FL, Col esterificat, Tg.
Col liber:
este utilizat la construcţia membranelor, sinteza de hormoni steroidici, acizi biliari
surplusul este esterificat şi depozitat în celulă.
Esterificarea Col e catalizată de E - acil-CoA- Col-aciltransferaza (ACAT), rezultînd esteri ai Col cu acizii palmitic, palmitooleic, oleic.

Слайд 317K
Colesterolul liber:
inhibă HMG-CoA-reductaza, ca rezultat sinteza Col în ţesuturile extrahepatice
inhibă sinteza R- B,E, astef determină micşorarea numărului lor pe membrană şi diminuarea captării LDL
activează E microzomială ACAT

Слайд 318II. ÎN MACROFAGE:
Macrofagele încorporează LDL care prezintă alterări ale componentelor proteice
Receptorii care recunosc LDL modificate nu sînt reglaţi prin feed-back negativ (cum se reglează receptorul clasic LDL) şi, ca urmare, prin încărcarea macrofagelor în mod excesiv cu Col apar “celulele spumoase”.
Modificările biochimice ale LDL constau în acetilări, glicozilări ale apo (apo B-100, apo E) şi /sau alterări oxidative ale acizilor graşi nesatiuraţi.

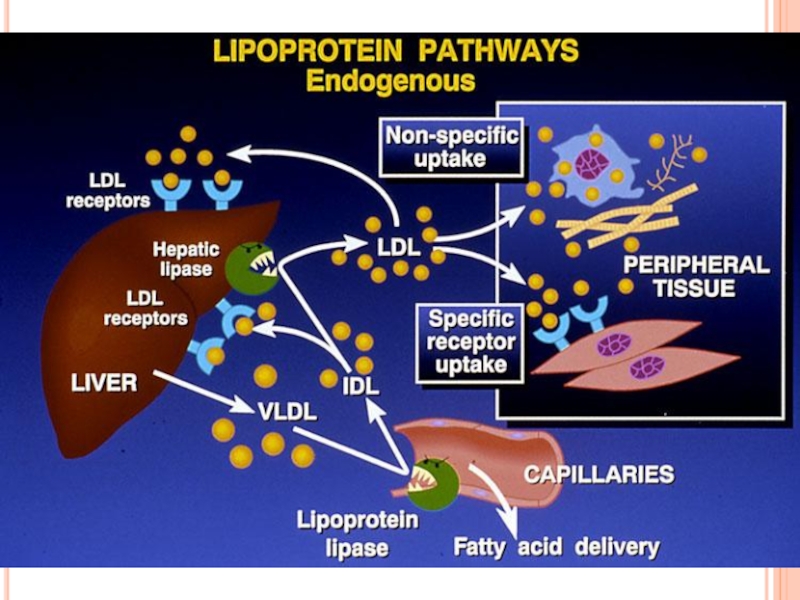
Слайд 320PATOLOGIA
Hiperlipoproteinemia de tip II – deficienţă calitativă şi cantitativă a R
IIa – mărirea Col LDL – xantomatoză
IIb – mărirea Col LDL+VLDL (Tg)- obezitate (lipsesc xantomele)
Risc de aterogeneză înaltă

Слайд 321α-LP - HDL
conţinut lipidic de 45-55%, predominînd FL – 20-30%, Col
Componenţa proteică - apo A (AI, AII) dar conţin cantităţi mici de apo C, D, E. Nu cuprind apo B.
HDL sînt secretate şi sintetizate de hepatocite şi enterocite sub forma unor particule nascente de formă discoidală, alcătuite dintr-un strat dublu lipidic (FL; Col) şi apo A, apo E.
Particulele nascente din intestin nu conţin apo E.

Слайд 322METABOLISMUL HDL
HDL captează Col liber atît din ţesuturile periferice cît și
Sub acţiunea LCAT (activată de apo AI)- Col este imediat esterificat şi migrează în interiorul particulei ---- HDL sferic matur (HDL3, sărace în CE, iar in final HDL2 bogate in CE ce sunt transportati la ficat)



Слайд 325CATABOLISMUL HDL
HDL sînt catabolizaţi la nivelul ficatului:
Prin intermediul apo E
Rolul principal al HDL este transportul Col din celulele extrahepatice în ficat, sediul catabolismului Col (transformare în AB şi excreţie prin bilă).

Слайд 326
LDL-R
50% of HDL C may
Return to the liver
On LDL via CETP
HDL

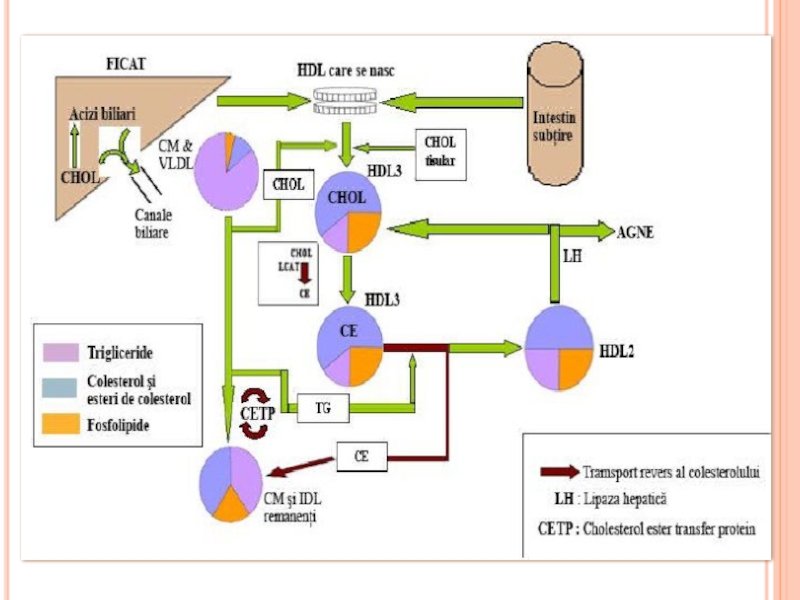
Слайд 328PATOLOGIA
Hipolipoproteinemia familiară – deficit de HDL
Cauza: deficit al sintezei de
Caracteristic: micşorarea HDL; Col HDL; FL
Splenomegalie, hipotrofie amigdaliană; anomalii neurologice


Слайд 330EXPLORĂRI UZUALE
Pentru efectuarea determinărilor:
pacienţii urmează
o dietă normală timp de 2
este eliminată medicaţia care ar putea afecta metabolismul lipidic
valorile obţinute – corelate cu vârsta şi sexul pacienţilor

Слайд 331EXPLORĂRI UZUALE
Aspectul plasmei
Lipidele totale
Colesterol total
Trigliceridele
HDL-colesterolul
LDL- colesterolul
Colesterol total/HDL-colesterol
Lipoproteina (a)
")
Слайд 3321. ASPECTUL PLASMEI
Clar
Normal
Hipercolesterolemie
Opalescent sau lactescent
Hipertrigliceridemie
Supernatant cu inel cremos
Creşterea chilomicronilor

Слайд 333COLESTEROLUL TOTAL
normal
de graniţă 5,2-6,2 mmol/L
înalt >6,2 mmol/L
HIPERCOLESTEROLEMIA
se întîlneşte în boli erediatre (hipercolesterolemia eriditară) sau dobândite (obezitate, sindrom nefrotic, ATS, hipotiroidie, icter obstructiv)


Слайд 335LDL-COLESTEROLUL
Formula de calcul:
LDL-col (mmol/L) = Col total – HDL-col – 0,45*TAG
LDL-col
normal <3,37 mmol/L
risc moderat al BIC 3,37 – 4,12 mmol/L
risc înalt al BIC >4,14
Creşterea LDL-Col apare în HLP ereditare (tip II a, II b) ; sau dobândite (hipotiroidie, sindrom nefrotic, DZ).
 = Col total – HDL-col – 0,45*TAGLDL-col (g/L) = Col total")
Слайд 336TRIGLICERIDELE
normal
acceptabil 1,6-2,2 mmol/L
nivel moderat 2,2-2,8 mmol/L
nivel înalt >2,8 mol/L
Hipertrigliceridemia
se întîlnesc în HLP esenţiale (tip II b, IV şi V) sau secundar în DZ, consum crescut de grăsimi, alcool sau glucide, în insuficienţa renală, utilizarea de corticosteroizi sau estrogeni.

Слайд 337HDL-COLESTEROLUL
Valorile normale:
Bărbaţi – 0,35 – 0,75 g/l (0,9-1,4 mmol/L);
Femei –
Nivele joase de HDL-col sunt prezente în afecţiuni ereditare (an-alfa-lipoproteidemia) şi dobîndite (obezitate, DZ, terapie cu progesteron, fumat, sedentarism).
Nivele crescute de HDL-col pot avea o cauză ereditară (hiper-alfa-lipoproteinemie) şi sunt corelate cu o creştere a longevităţii
;Femei – 0,45 – 0,85")

Слайд 339 LIPOPROTEINA (A)
LP modificată din clasa LDL (LDL + proteină adiţională)
structură
în concentraţii crescute se depozitează în pereţii arteriali inducând o
aterogeneză accelerată
LP modificată din clasa LDL (LDL + proteină adiţională)structură similară cu a plasminogenuluiîn")
Слайд 340DISLIPOPROTEIDEMIE SAU DISLIPIDEMIE
modificarea c% lipidelor totale plasmatice, a uneia dintre fracţiuni
Se clasifică:
Hiperlipiproteidemii (primare şi secundare)
Hipolipoproteidemii primare (ereditare, familiale)

Слайд 341HIPERLIPIDEMIILE
Hiperlipidemiile primare sunt de natură genetică,
Hiperlipidemiile secundare sunt determinate de

Слайд 342HLP PRIMARE
după Fredrickson, în funcţie de aspectul fenotipic , HLP primare

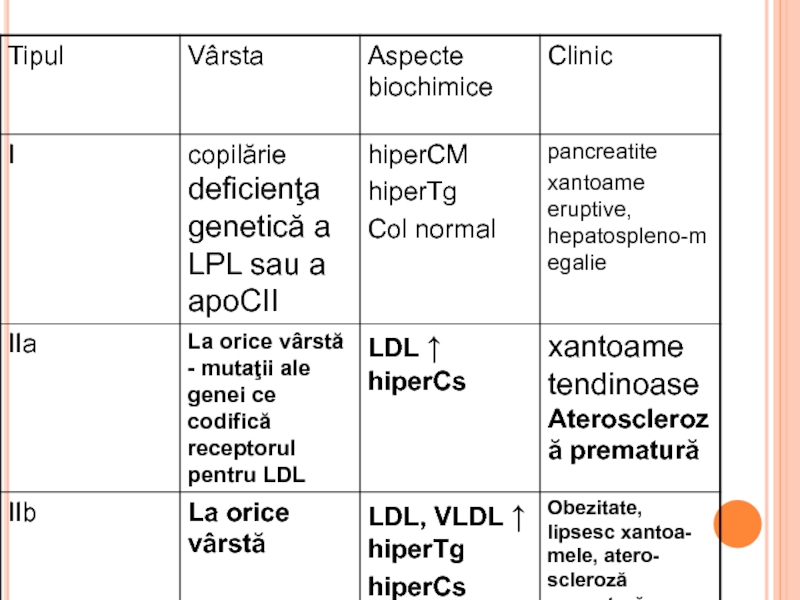
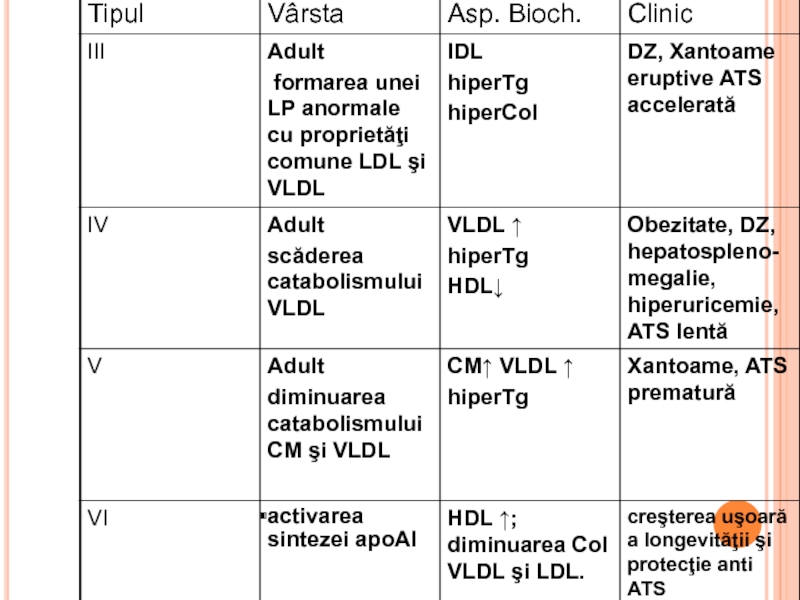

Слайд 346HIPERLIPIDEMIILE SECUNDARE
Diabetul zaharat:
Lipsa de insulină:
lipoliza activată,
lipogeneza inhibată

Слайд 347DISLIPIDEMIA DIABETICĂ
Mărirea concentraţiei AGL ( activarea trigliceridlipazei hormon-sensibile);
Sporirea concentraţiei TAG-VLDL (activarea
Dimimuarea concentraţiei HDL;
Prezenţa LDL mici şi dense;
Nivel înalt de LP modificate (oxidări, glicări etc.);
;Sporirea concentraţiei TAG-VLDL (activarea sintezei AG în ficat→sinteză")
Слайд 348HIPERLIPIDEMIA ALCOOLICĂ:
se observă o lipidemie alimentară o dată cu consumul de
e însoţită şi de o secreţie intensă a pre-β-LP, de o hiperchilomicronemie - ↑TAG
un clirens lent al ambelor particule
Steatoză hepatică

Слайд 349Alcoolul amplifică sinteza şi secreţia pre-β-lipoproteinelor determinate de valorile majorate ale
NADH +H inhibă oxidarea AG şi măreşte sinteza TAG şi a pre-β-lipoproteinelor (VLDL).
Alcoolul favorizeză inducţia enzimelor microzomale în ficat.

Слайд 350SUBSTANŢE LIPOTROPE
Ficatul este un organ specific de sinteză a fosfolipidelor plasmei.
În
În consecinţă, are loc depozitarea lipidelor în ficat.
Orice substanţă donatoare de CH3 pentru sinteza colinei este denumită lipotropă.

Слайд 351OBEZITATEA
Excesul de aport caloric, raportat la cheltuielile energetice ale organismului

Слайд 352OBEZITATEA
Evaluarea obezităţii se efectuează prin calcularea indicelui de masă corporală (IMC)
IMC
IMC = greutatea(kg)/înălţimea(m) la pătrat")
Слайд 353TIPUL OBEZITĂŢII
Se stabileşte în funcţie de distribuţia ţesutului adipos
Se evaluează prin
")

Слайд 355DISLIPIDEMIA DIN OBEZITATE
Hipertrigliceridemie;
Hiperlipidemie postprandială;
Scăderea HDL-colesterolului, în special fracţiunea HDL2;
Creşterea raportului Col
Creşterea concentraţiei apoB;
Majorarea proporţiei de LDL mici şi dense (fenotipul B).

Слайд 356ATEROSCLEROZA
l
Se caracterizează prin depunerea Col în intima vaselor sub formă de
Se reduce elasticitatea pereţilor arteriali
favorizează BIC şi a accidentelor vasculare cerebrale.l

Слайд 357FACTORI DE RISC MAJOR AL ATEROSCLEROZEI
Dislipidemiile ereditare (IIa, IIb şi III)
Hipertensiunea arterială;
Fumatul;
Diabetul zaharat (în special de tipul 2);
Sexul masculin (până la 75 ani).
 şi dobândite (diabetul zaharat,")
Слайд 358FACTORI DE RISC MODERAT AL ATEROSCLEROZEI
obezitatea (în special cea abdominală);
sedentarismul;
stresul cronic;
hiperuricemia;
majorarea
diminuarea concentraţiei acidului folic;
hipervitaminoza D;
utilizarea contracepţionalelor orale.
;sedentarismul;stresul cronic;hiperuricemia;majorarea concentraţiei homocisteinei;diminuarea concentraţiei acidului")
Слайд 359ATEROSCLEROZA
Patogenie:
leziunea pereţilor endoteliali
Infiltrarea LDL oxidate
Proliferarea “celulelor spumoase”
Formarea unor depozite lipidice la
Alte modificări: proliferarea fibrelor musculare netede, infiltrarea macrofagelor şi a limfocitelor, eliberarea locală de citokine; agregarea trombocitelor şi extinderea leziunilor endoteliale iniţiale.




Слайд 363PROFILUL LIPIDIC ATEROGEN
Majorarea LDL (îndeosebi LDL mici şi dense, LDL modificate
Mărirea apo B100 (reflectă mai exact numărul de particule LDL, comparativ cu calcularea LDL-colesterolului);
Diminuarea concentraţiei HDL:
Sporirea concentraţiei LP(a);
Creşterea concentraţiei trigliceridelor (în special când este însoţită de mărirea concentraţiei AGL):
- sporirea TAG contribuie la formarea LDL mici şi dense.

Слайд 364HIPOLIPOPROTEIDEMIILE PRIMARE (EREDITARE, FAMILIALE)
sunt datorate genelor care controlează sinteza, transportul sau
afecţiunea Tangier,
A beta-lipoproteinemia familială;
sunt datorate genelor care controlează sinteza, transportul sau utilizarea LP. afecţiunea Tangier,")
Слайд 365HIPOLIPOPROTEIDEMIILE PRIMARE
afecţiunea Tangier - cauzată de insuficienţa sau lipsa totală de
acumularea în ţesuturi a esterilor colesterolului.
Splenomegalie, anomalii neurologice, hipertrofie amigdaliană
Afecţiunea este incurabilă.
, datorate deficitului")
Слайд 366A BETA-LIPOPROTEINEMIA FAMILIALĂ
Afecţiune transmisă autosomal recesiv
Cauzată de deficienţa ereditară
Diminuarea sintezei şi secreţiei VLDL şi/sau accelerarea catabolismului VLDL.
Reducerea Tg, Col, FL; sunt absente LDL, VLDL, CM
Malabsorbţia lipidelor şi a vitaminelor liposolubile;
Neuropatie, ataxie.

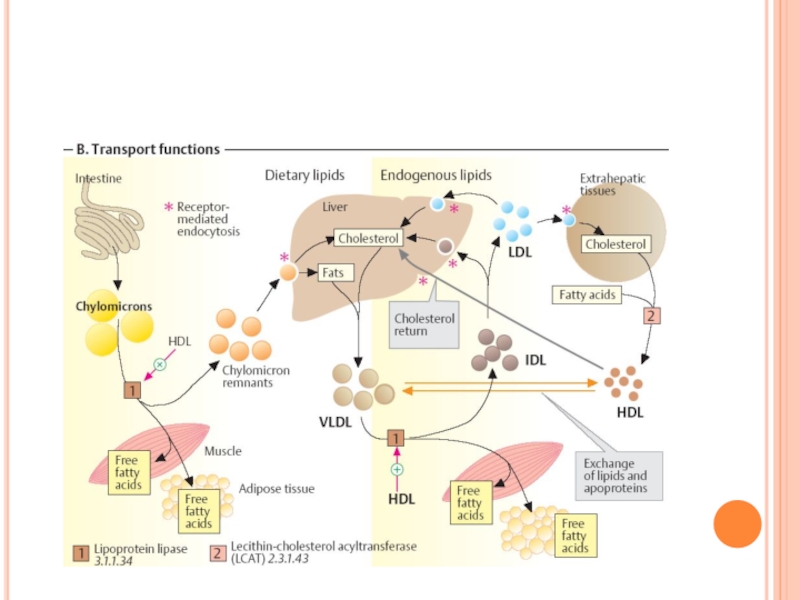
Слайд 368VITAMINELE
sunt substanţe biologic active necesare pentru creşterea, dezvoltarea şi supravieţuirea organismului.
nu se sintetizează în organismul uman
(prezenţa lor în alimente este obligatorie).
nu servesc în calitate de sursă energetică, dar îndeplinesc funcţii coenzimatice.

Слайд 369AVITAMINOZE ŞI HIPOVITAMINOZE
avitaminoze - lipsa vitaminelor în alimente
hipovitaminoze - insuficienţa
Cauzele apariţiei lor:
pot fi exogene (atunci când vitamina respectivă lipseşte sau se conţine în produsele alimentare insuficient)
endogene (dereglări în absorbţie, transport, antivitamine etc.)

Слайд 370PROVITAMINE
- predecesorii vitaminelor, care în organism se transformă în vitaminele respective

Слайд 371ANTIVITAMINELE
– substanţe (inclusiv şi unele medicamente) ce se aseamănă mult ca
La majorarea c% lor - în mod competitiv ele se vor uni cu E în locul vitaminelor --- se formează complexul apoenzimă-antivitamină lipsit de activitate --- se dereglează reacţiile metabolice catalizate de aceste enzime.
 ce se aseamănă mult ca structură cu unele vitamine.")
Слайд 372ANTIVITAMINELE
Preparatele sulfanilamidice prezintă antivitamine pentru microorganisme deoarece ele mult se aseamănă
La majorarea concentraţiei în organism a acestor medicamente, ele se vor include în sistemele enzimatice ale bacteriilor şi bacteriile vor pierde capacitatea de a se înmulţi (efect bacteriostatic).

Слайд 373VITAMINELE LIPOSOLUBILE
A, D, E, K:
Vitamine cu molecule apolare, hidrofobe, solubile în
Sunt derivaţi izoprenici
Transportate prin CM la ficat şi depozitate (vit A; D; K – în ficat; vit E – în ţesutul adipos)
Deoarece sunt depozitate în organism, supradozarea lor conduce la declanşarea unor stări de toxicitate

Слайд 374VITAMINA A
vitamina creşterii, retinol, antixeroftalmică
La baza structurii acestei vitamine se află


Слайд 376METABOLISM
Digestia are loc în intestin
Absorbţia – în regiunea superioară a intestinului
Retinolul

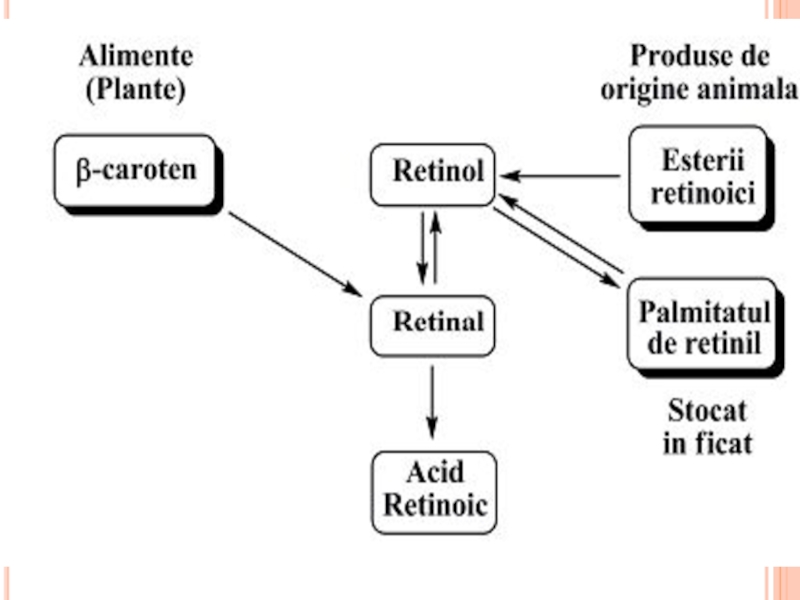
Слайд 378ROLUL VITAMINEI A
Retinol – acţiune hormonală decesivă pentru funcţia de reproducere
Retinal
A. retinoic –participă la sinteza glicoproteinelor, datorată acţiunii de factor de creştere şi de diferenţiere tisulară a lui
Retinolul şi acidul retinoic au acţiune de hormoni steroidici

Слайд 379METABOLISMUL
All-trans-retinol
retinol
RBP
Retinol-RBP
retinol
RBP
Acid retinoic
Activarea genei
TESUT
Chilomicroni remanenti
chilomicroni
Transport vitA
Retinil palmitat
Depozitarea v. in t.adipos si ficat
Retinol
All-trans-retinol
All-trans-retinal
11-cis-retinal
rodopsina
All-trans-retinal
lumina
opsina
Vit.A si vazul ,11-cis-retinal e component rodopsinei
Esteri retinil
Retinol
Retinol
Retinal
B-caroten
Esterii retinil
B-caroten
dieta
Acizi grasi
Acizi grasi coA
CELULA INTESTINALA
Sursa de vit A
RBP
RBP


Слайд 381CICLUL VIZUAL
Vit A este o componentă a pigmenţilor vizuali din celulele
Bastonasele contin pigmentul vizual rodopsina care este format din 11-cis retinal şi opsina (o glicoproteina).

Слайд 382CICLUL VIZUAL
Când rodopsina este expusă la lumină, 11-cis-retinalul este convertit la
Acest fapt conduce la generarea unui impuls nervos care e transmis de nervul optic la creier.
Regenerarea rodopsinei necesită izomerizarea all-trans-retinalului în 11-cis retinal (prin absorbţie de fotoni luminoşi) care se combină spontan cu opsina. ---- ciclul este încheiat.

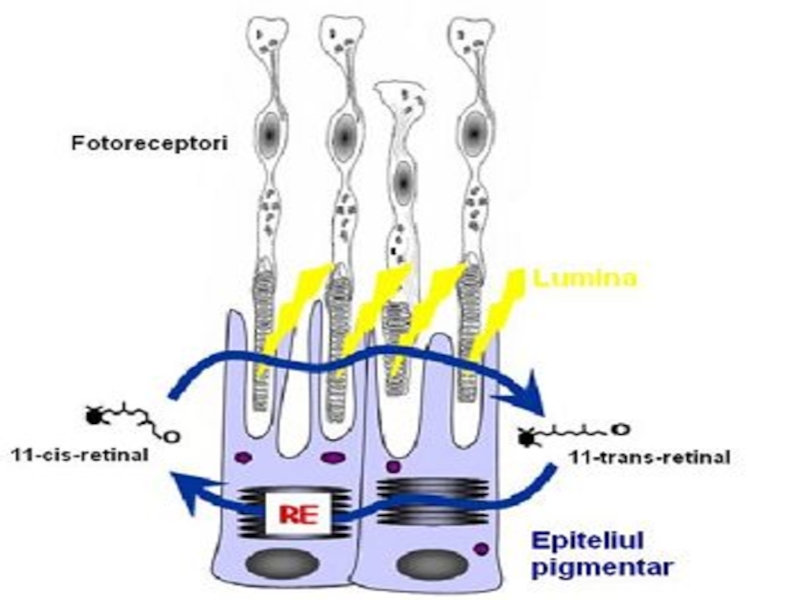
Слайд 384CICLUL VIZUAL
O parte din trans-retinal este disponibil pentru un nou ciclu
O altă parte însî este convertit în acid retinoic şi nu mai este disponibil pentru a forma rodopsina.
Aceste pierderi sunt compensate de aportul alimentar de vitamina A sau din rezervele stocate în ficat

Слайд 385ROLUL RETINOLULUI
necesar pentru reproducere si crestere
favorizează spermatogeneza şi dezvoltarea placentei
mentine integritatea celulei epiteliale a tractului gastrointestinal, pielii, tractului respirator sau urinar şi a glandelor salivare
necesar pentru menţinerea stratului mielinic din ţesutul nervos
necesar pentru creşterea dinţilor şi oaselor

Слайд 386RETINOL ŞI ACIDUL RETINOIC
necesari pentru sinteza glicoproteinelor sau mucopolizaharidelor
sunt implicaţi
Acidul RETINOIC – promovează creştrerea şi diferenţierea celulelor epiteliale

Слайд 387ALTE EFECTE:
La oxidarea AG
Biosinteza gliceridelor, fosfolipidelor, Col, h. steroizi
Creşte asimilarea purinelor
Sunt reglatori ai permeabilităţii membranelor
Afectează expresia genetică implicată atât în proliferarea cât şi diferenţierea celulelor normale sau maligne

Слайд 388VITAMINA A
Se conţine în produse de origine animală (unt, ficat etc.).
. Unele produse de origine")
Слайд 389CARENŢA VITAMINEI A
se caracterizează prin:
Orbirea nocturna
La nivelul ochilor: xeroftalmia (uscarea
La nivelul pielei şi mucoaselor: hiperkeratoza, descuamaţie, ulcerări, inflamaţii.
vitamina A este utila in tratarea unor boli de piele (acnee, psoriasis).
; în cazuri grave")
Слайд 390VITAMINA D (ANTIRAHITICĂ)
Se cunosc mai multe forme:
D2 – ergocalciferol - origine
D3 – colecalciferol – origine animală ( se formează din 7-dehidrocolesterol)
Structura chimică: derivaţi ai ciclului steran:
Se cunosc mai multe forme:D2 – ergocalciferol - origine vegetală (se formează din")
")
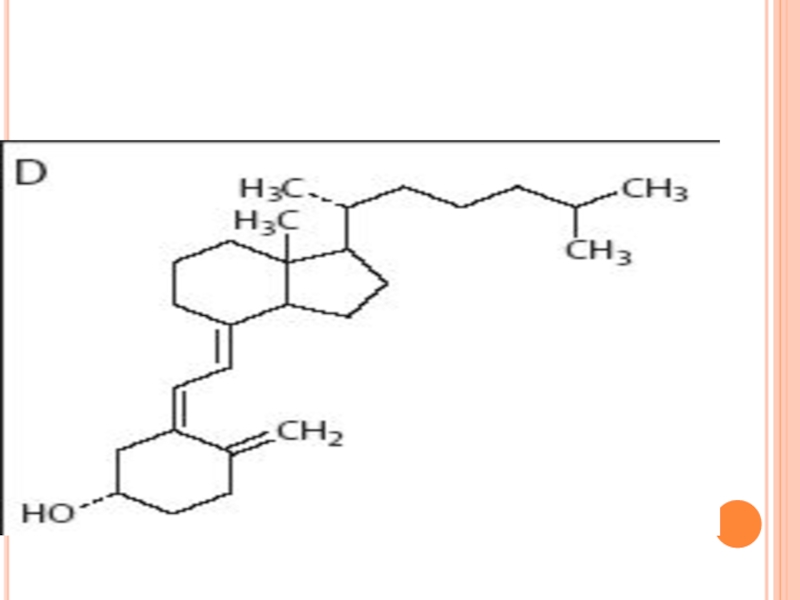
Слайд 393METABOLISM:
În intestin se supune acţiunii emulsionante a bilei, apoi este absorbită
Prin sânge – în ficat ajunge şi vit D3 formată din 7- dehidrocolesterol în piele, sub acţiunea radiaţiilor ultraviolete din lumina solară.

Слайд 394CALCITRIOLUL
În ficat vit D3 suferă o hidroxilare – 25- hidroxi- colecalciferol
În rinichi 25- hidroxicolecalciferol suferă o nouă hidroxilare – 1,25-dihidroxicolecalciferol - calcitriol (E- 1α-hidroxilază) – transportat la ţesuturile ţintă: intestin şi oase. Are rol important în menţinerea nivelului Ca seric.
Reglarea – PTH şi nivelul de Ca în sânge.
 - forma")
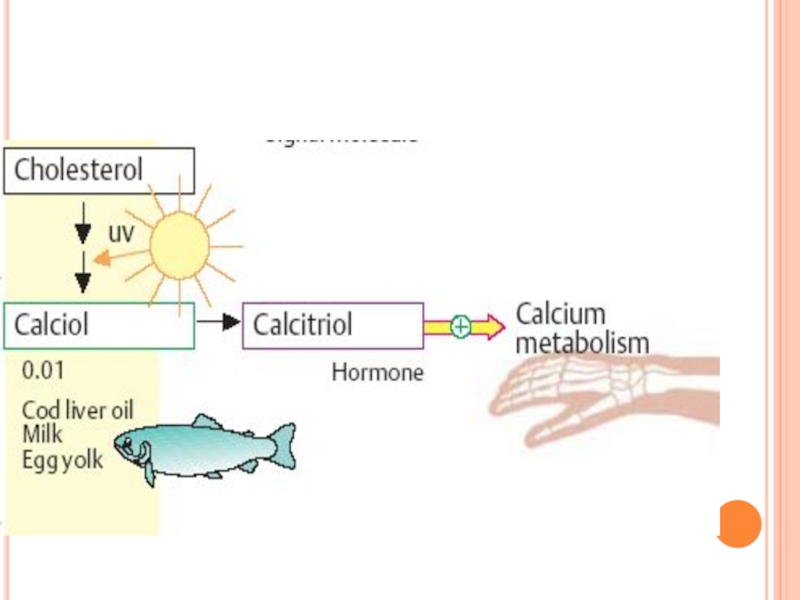
Слайд 396EFECTELE VITAMINEI D
La nivel de intestin: măresc absorbţia Ca şi fosfatului
La nivelul tubilor renali: favorizează reabsorbţia fosfaţilor (contracarând eliminarea lor stimulată de PTH)
La nivelul osului: stimulează mobilizarea Ca şi fosfatului din os care necesită prezenţa PTH.
intervine în mineralizarea oaselor prin mărirea formării de hidroxiapatită.

Слайд 397VITAMINA D (ANTIRAHITICĂ)
Carenţa - apariţia şi dezvoltarea rahitismului la copii
este afectat scheletul: oasele sunt mai moi, picioarele se deformează sub greutatea corpului; fontanelele se închid cu întârziere, se reţine dezvoltarea dinţilor; tonusul muscular este redus etc.
În sânge scade conţinutul calciului şi fosfatului.
 Carenţa - apariţia şi dezvoltarea rahitismului la copii din cauza dereglării metabolismului")
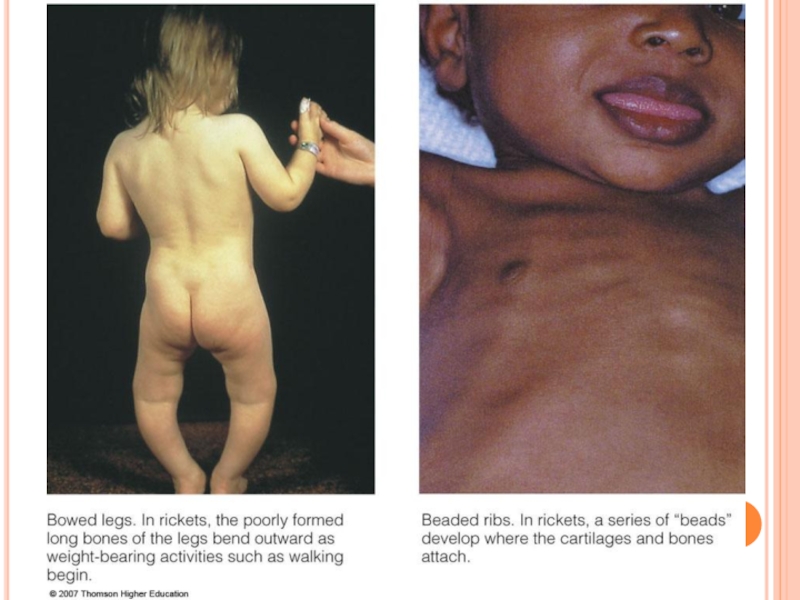
Слайд 399SURSE DE VITAMINA D
ficatul de peste, somon, sardine, macrou, cantitati mici
galbenusul de ou, unt, ciuperci.

 Structura chimică: are la bază tocolul (5,7,8-trimetiltocol)")
Слайд 401METABOLISMUL VITAMINEI E
Absorbită la nivelul intestinului subţire - sânge-ficat şi alte
Se excretă prin bilă şi intestin (în prealabil se supine unei oxidări şi se conjugă cu a glucuronic)

Слайд 402ROLUL VITAMINEI E
Este antioxidant puternic - protejează oxidarea AG nesaturaţi în
intervine în metabolismul seleniului (care întră în componenţa glutation peroxidazei ce asigură protecţia membranelor faţă de acţiunea radicalilor peroxizi).
Sunt implicate în sistemul E respiratorii, favorizând cuplarea fosforilării oxidative

Слайд 403DEFICIENŢA VITAMINEI E
are ca urmare afectarea aparatului reproductiv: stagnarea embriogenezei,
distrofie musculară,
infiltrare adipoasă a ficatului,
Distrugerea globulelor roşii
Reducerea stabilităţii membranelor şi sărăcirea în colagen a ţesutului conjuctiv
Ateroscleroză, cancer.

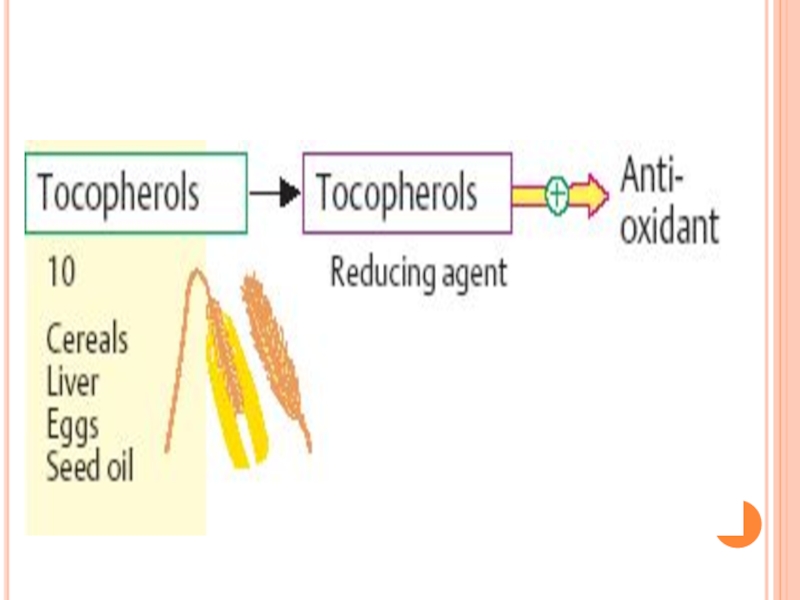
Слайд 405VITAMINA E
Se conţine în uleiuri vegetale (porumb, floarea soarelui etc.) carne,
Necesarul:
Zilnic: 7mg la adult şi 2-3 mg la copii
 carne, unt, gălbenuş de ou,")
Слайд 406VITAMINA K (ANTIHEMORAGICĂ)
K1- filochinona (izolată din vegetale)
K2- menachinona (izolată din ţesuturile
Mai deosebim:
Ftiocolul- constituient al membranelor lipiduce din bacilul Koch
Menadiona - produs de sinteză folosit în terapeutică (are structură şi acţiuni asemănătoare cu a produsului natural)
La baza structurii: nucleul p-naftochinonei
 K1- filochinona (izolată din vegetale)K2- menachinona (izolată din ţesuturile animale şi bacteriile")

Слайд 408ROLUL VITAMINEI K
Participă la coagularea sângelui – este direct implicată în
Vitamina K ajută la modificarea restului de Clu în γ-carboxiglutamic (se adaugă o grupă carboxil ce poate fixa Ca)
Intervine şi în procesele de fosforilare


Слайд 410CARENŢA VITAMINEI K
Carenţa acestei vitamine are ca consecinţă apariţia hemoragiilor în

Слайд 411CARENŢA VITAMINEI K
Hemoragiile pot apărea când bolnavul utilizează cantităţi considerabile de
La om hipovitaminoza K în condiţii obişnuite se întâlneşte relativ rar deoarece această vitamină se sintetizează de microflora intestinală.

Слайд 412Antivitaminele acestei vitamine, de ex. dicumarolul sunt utilizate în clinică ca
Există analogi sintetici hidrosolubili a acestei vitamine, de ex. vicasolul.
ANTIVITAMINELE VITAMINEI K

Слайд 413LIPIDOZE TISULARE
Boli ereditare, caracterizate prin acumularea de sfingolipide în creier şi

Слайд 414LIPIDOZE TISULARE
1. Neimann-Pick - deficienţa de sfingomielinază
sfingomielina SM- se acumulează în
Hepato- şi splenomegalie, tulburări digestive, întîrziere psihomotorie, convulsii
Decedează în vârsta fragedă

Слайд 415LIPIDOZE TISULARE
2. afecţiunea Tay-Sachs – lipsa β-N-acetilhexozaminidazei (hidrolază ce scindează N
Acumularea de GM2 - tumefierea celulelor ganglionare ale scoarţei creierului – creşte în volum
pe retina ochiului – apar pete de culoare roşie-vişinie – gangliozid GM2
Acumularea de")
Слайд 416LIPIDOZE TISULARE
3. Boala Krabbe – deficit de galactocerebrozidază
Demielinizări la nivelul creierului
Paralizie,
deces în vârsta fragedă

Слайд 417LIPIDOZE TISULARE
4. boala Gaucher – deficit de hidrolază ce scindează glucoza
Splenomegalie (se acumulează în splină); tulburări hematologice ( membrana eritrocitelor are cantităţi mari de gangliozide)
Osteoporoză, retard mintal

Слайд 418LIPIDOZE TISULARE
5. leucodistrofia metacromatică – deficit de sulfatidază
Acumularea sulfatidelor în substanţa
Tulburări neurologice şi psihice

Слайд 419LIPIDOZE TISULARE
boala lui Fabry (lipogranulomatoza) - deficienţa ceramidazei lizozomale (acide)
cauzează
moştenită autosomal recesiv
leziuni granulomatoase în piele, articulaţii şi laringe,
disfuncţii moderate a sistemului nervos;
de asemenea poate implica inima, plămînii, şi nodulii limfatici.
De obicei fatală în timpul primilor câţiva ani de viaţă.
 - deficienţa ceramidazei lizozomale (acide) cauzează acumulări de ceramide. moştenită")





